
Abstract
Abstract
Kong, Fanyi, Shixiang Liu, Qiong Li, and Lin Wang. Sleep architecture in partially acclimatized lowlanders and native Tibetans at 3800 meter altitude: What are the differences? High Alt Med Biol 16:223–229, 2015.— It is not well known whether high altitude acclimatization could help lowlanders improve their sleep architecture as well as Native Tibetans. In order to address this, we investigated the structural differences in sleep between Native Tibetans and partially acclimatized lowlanders and examined the association between sleep architecture and subjective sleep quality. Partially acclimatized soldiers from lowlands and Native Tibetan soldiers stationed at Shangri-La (3800 m) were surveyed using the Pittsburgh Sleep Quality Index (PSQI), Hamilton Anxiety Scale (HAMA), and Hamilton Depression Rating Scale (HAMD). The sleep architecture of those without anxiety (as determined by HAMA>14) and/or depression (HAMD>20) was analyzed using polysomnography and the results were compared between the two groups. One hundred sixty-five male soldiers, including 55 Native Tibetans, were included in the study. After partial acclimatization, lowlanders still exhibited differences in sleep architecture as compared to Native Tibetans, as indicated by a higher PSQI score (8.14±2.37 vs. 3.90±2.85, p<0.001), shorter non-rapid eye movement (non-REM) sleep (458.68±112.63 vs. 501±37.82 min, P=0.03), lower nocturnal arterial oxygen saturation (Sp
Introduction
W
Hypoxemia at high altitude is most severe during sleep (Wickramasinghe et al., 1999), but Native Tibetans were reported to have better capacity to adapt to a hypoxic environment at high altitude and, therefore, to have a better quality sleep than Han lowlanders (Wu et al., 2005), and the ability of Native Tibetans to adapt to high altitude may largely be attributed to genetic adaptations (Niermeyer et al., 1995; Wu et al., 2006). However, it is still not known whether acclimatization to high altitudes could help relieve hypoxia in lowlanders, thereby improving their sleep architecture to similar levels as Native Tibetans. The present study investigated the differences in sleep structure between Native Tibetans and partially acclimatized lowlanders at high altitude and explored the influence of sleep architecture on subjective sleep quality.
Materials and Methods
Study participants and setting
Soldiers stationed at Shangri-La in Diqing Tibetan Autonomous Region, Yunnan Province, China, were surveyed in this study. Shangri-La has an altitude of 3800 m, which is in the range considered as “very high altitude” (from 3500 to 5800 m) by Davis et al. (2005). New recruits, including Native Tibetans and lowlanders, were trained at the boot camp at an altitude of 1800 m for 3 months before being sent to Shangri-La. After they had been partially acclimatized to the higher altitude for at least 3 months, basic demographic and health information were collected from each participant, including age, ethnicity, body mass index (BMI), length of service, education, military rank, cigarette smoking, and alcohol consumption. We also surveyed each participant using the Pittsburgh Sleep Quality Index (PSQI) (Buysse et al., 1989), Epworth Sleepiness Scale (ESS) (Johns, 1991), Hamilton Anxiety Scale (HAMA) (Hamilton, 1959), and the Hamilton Depression Rating Scale (HAMD) (Hamilton, 1967). Poor sleepers were defined as those with PSQI score >5 (Buysse et al., 1989).
Inclusion and exclusion criteria for participation
All Chinese Han and Native Tibetan soldiers who had been deployed in Shangri-La for at least 3 months were initially screened for inclusion in the study. Those soldiers with an HAMA score >14 (Hamilton, 1959) and/or HAMD score >20 (Hamilton, 1967) were excluded, to rule out the potential influences of anxiety and depression on sleep quality.
Measurements of sleep architecture
Sleep architecture was measured by polysomnography (PSG). Measures recorded included electroencephalogram (EEG), electrooculogram, electromyogram, chest wall abdominal movements, oronasal airflow, arterial oxygen saturation (Sp
Prior to this study, none of the subjects had ever had a PSG or had taken any hypnotics. Alcohol and caffeinated beverages were not available in camp, and neither black nor green tea consumption was permitted for participants during the study period. Participants were recorded for two consecutive nights, but only the data collected from the second night's PSG were included for analysis. On the nights of the study, participants were asked to go to bed at 22:00 and were asked to wake up at 7:00 the next morning.
The following data were obtained and calculated from PSG recordings: sleep period time (SPT, time from falling asleep to last awakening), total sleep time (TST, calculated as a sum of stages 1, 2, and 3+4 non-REM and REM sleep), sleep onset latency (SOL, time from lights out to the first episode of stage 2 sleep), wake after sleep onset (WASO, total time of wakefulness after sleep onset), number of arousals (assessed by scoring EEG arousals that was defined as wakefulness episodes of 3–15 sec) (Hori et al., 2001), sleep efficiency index (SEI, calculated as [TST/SPT]×100%), stage 1 sleep to TST ratio, stage 2 sleep to TST ratio, stage 3+4 sleep to TST ratio, time in REM and non-REM sleep, non-REM sleep latency, REM latency, mean Sp
Ethics
Written informed consent was obtained from each participant. The research project was approved by the Ethics Committee of Kunming General Hospital of Chengdu Military Command, People's Liberation Army (China).
Statistical analysis
Continuous variables with a normal distribution were expressed as mean values and standard deviations. Those consistent with the skewness distribution were expressed as medians (quartiles). Qualitative variables were analyzed using percentages. Comparisons of continuous variables between two groups with normal distributions were analyzed using Student's t-test, and comparisons of two groups with a skewed distribution were analyzed using the Mann-Whitney U test. The chi-square test (χ2 test) was used to test for differences in proportions of qualitative variables.
A stepwise multiple linear regression analysis was performed to adjust for possible confounding variables found to be significantly different between the two groups, including length of service. All statistical levels quoted are two-tailed. The 95% confidence intervals (CIs) were calculated where appropriate. A p value <0.05 was considered statistically significant. All data were analyzed using SPSS for Windows (version 16.0, SPSS Inc. Chicago, IL, USA, 2006).
Results
Sociodemographic and health profiles of subjects
Two hundred forty male soldiers serving on the Diqing Plateau were initially screened for this study, with 165 of those included in the study based on the exclusion criteria outlined in the Methods section. A participant flow sheet is outlined in Figure 1. Of the soldiers enrolled in this study, 55 were Native Tibetans and 110 were Chinese Han. Prior to joining the army, 49 of the 55 Tibetans were residents of Lhasa (3650 m), and the remaining soldiers were from Qamdo (3240 m), Dingjie (4500 m), Shigatse (3836 m), Mangkang (3780 m), Linzhi (3000 m), and Deqin (3380 m). The average age of the enrolled soldiers was 21.21±3.30 years and the median service time was 11 months (11 to 41 months). None of the subjects had a previous medical history. Han lowlanders and Native Tibetan soldiers were similar in terms of age (21.50±3.47 vs. 20.64±3.10, p=0.49), marital status (4.54% vs. 9.1%, p=0.61), body mass index (21.88±2.04 vs. 22.14±3.05, p=0.78), HAMA score (9.45±5.76 vs. 8.10±8.74, p=0.61), HAMD score (8.41±4.55 vs. 10.20±11.92, p=0.54), ESS (7.38±4.79 vs. 6.00±3.76, p=0.41), cigarette smoking (45.45% vs. 36.36%, p=0.69), alcohol consumption (13.64% vs. 9.10%, p=0.73), education level (p=0.24), and military rank (p=0.057).
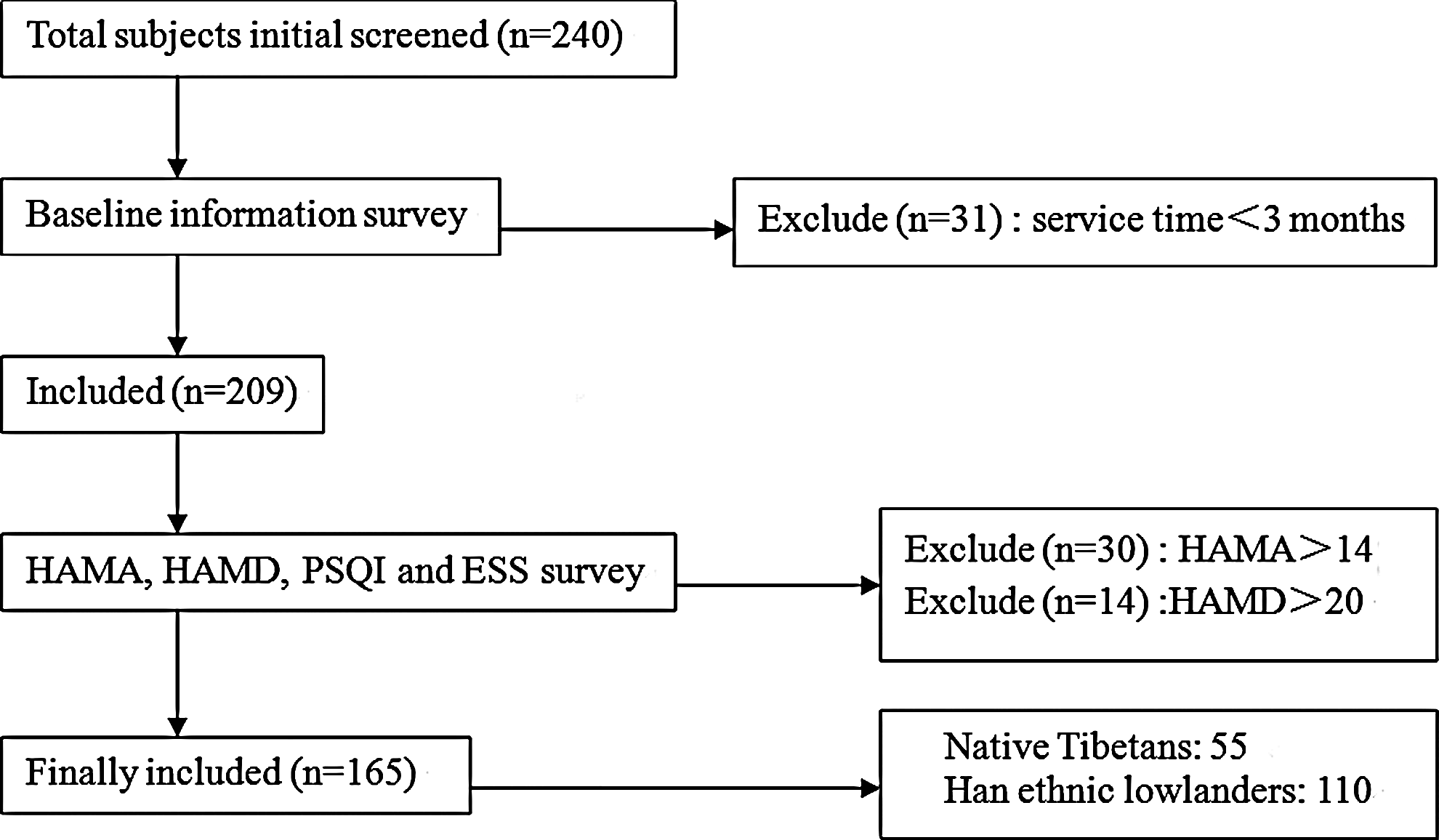
Participant flow sheet.
Han soldiers coming from the lowlands had a longer service time (median 20 vs. 11 months, p=0.047), higher PSQI (8.14±2.37 vs. 3.90±2.85, p<0.001), and a greater proportion of poor sleepers than the Native Tibetan soldiers (90.91% vs. 45.45%, P=0.004). See details in Table 1.
Student t test; †χ2 test; $Kruskal Wallis test.
Sleep parameters were mostly similar in partially acclimatized Han lowlanders and Native Tibetans
No significant differences between the partially acclimatized Han lowlanders and the Native Tibetans were detected in most of the sleep architecture recordings (Table 2). The only variable tested that was statistically different between the two groups in the initial analysis was their nocturnal Sp
Student's t test; †Mann-Whitney U test.
Distribution of SpO2 reduction during the sleep period was mostly similar in partially acclimatized Han lowlanders and Native Tibetans
The majority of periodic breathing (as reflected by the number of times Sp
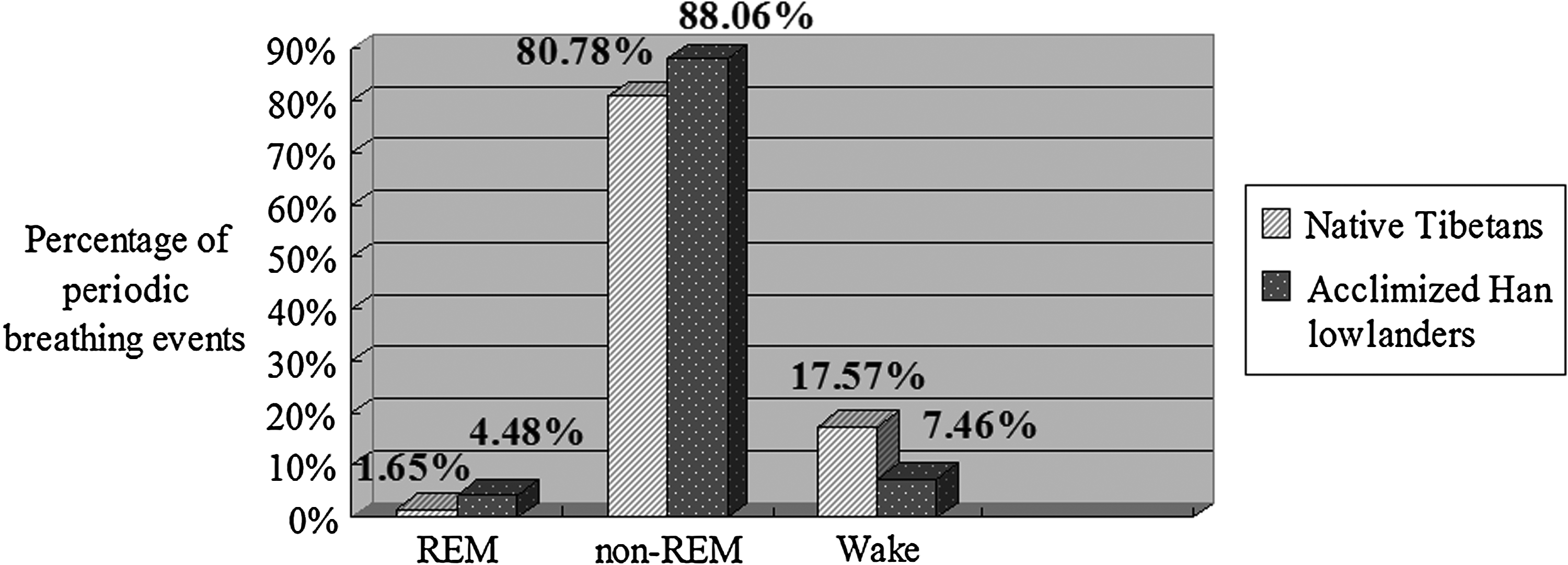
Periodic breathing events comparison between two groups.
Subjective sleep quality and related to SOL, non-REM and nocturnal Spo2
Despite limited objective differences in sleep architecture, subjective PSQI evaluation suggests that sleep differences between the two groups exists. To address the inconsistencies between the objective and subjective measures, we conducted a multiple linear regression analysis to explore the association between PSQI and sleep architecture. In this regression model, age, service time, BMI, ESS, HAMA, HAMD were adjusted as possible confounders. Using this adjustment, SOL (β=0.08, 95% CI: 0.01 to 0.15) and non-REM latency (β=0.011, 95% CI: 0.001 to 0.02) were slightly positively correlated with PSQI, while mean nocturnal Sp
Length of stay at high altitude did not affect sleep architecture
In order to determine if the length of time the Han soldiers had been at high altitude affected the sleep variables, a multiple linear regression analysis was employed to adjust potential confounders such as age, BMI, HAMA, and HAMD. However, no significant associations were detected between service time at high altitude and any of the sleep parameters for lowlanders.
Discussion
Our present study concentrated on the sleep architecture differences between partially acclimatized lowlanders and Native Tibetans living in high altitude. The majority of the sleep parameters obtained from PSG between the two groups at an altitude of 3800 m were not significantly different. However, two distinct differences were detected: partially acclimatized lowlanders had a non-REM sleep time curtailment and a lowered nocturnal Sp
We did not detect any differences in sleep structure between partially acclimatized lowlanders and native Tibetans. Additionally, the distribution of periodic breathing throught the whole sleep period was also similar in both groups. These results differ from current available studies that reported notable differences in sleep structure for lowlanders exposure to high altitude, including frequent awakening and reduction in non-REM sleep (Himashree et al., 2002), a shorter sleep time, a shorter amount of time spent in stage 2 non-REM sleep, a longer time spent in stage 1 non-REM sleep (Plywaczewski et al., 2003), and most evidently, a significant increase in periodic breathing (Bloch et al., 2010). One possible explaination for the variance between our data and those previously published results is that the previous studies were conducted with populations of lowlanders that were only acutely exposed to high altitude (Goldenberg et al., 1992; Salvaggio et al., 1998; Plywaczewski et al., 2003; Przybylowski et al., 2003; Panjwani et al., 2007) rather than the partially acclimatized lowlanders in our studies.
Because of this, the reported increase in periodic breathing in lowlanders with acute exposure to high altitude may largely be the result of physiological adaptive measures to hypoxia, meant to prevent sleep-hypoventilation. However, according to results by Bloch et al. (2010), periodic breathing is independently correlated with days of acclimatization for partially acclimatized lowlanders, and the ventilation appeared to decrease after years of high-altitude exposure due to hypoxic desensitization, suggesting that the partial acclimatization does alter these measures and therefore, explains the discrepancies between these studies. Additionally, genetic variations between Native Tibetans and lowlanders may also account for some of the variability between the two groups (Peng et al., 2011; Xu et al., 2011).
Acute exposure to high altitude generally leads to an increase in stage 1 and stage 2 non-REM sleep and decrease in REM sleep, while total sleep time is unchanged. However, some studies have suggested that no changes in stage 1 and stage 2 non-REM sleep and REM sleep occur below the altitude of 4000 m (Nicholson et al., 1988; Normand et al., 1990). In our studies with soldiers partially acclimatized to high altitude, we did not detect a difference in non-REM sleep time between the two groups. However, after we adjusted for the length of time the soldier had spent in the army, lowlanders did have a shorter non-REM sleep than Native Tibetans. This may be due in part to the potential influence of maladjustment of army life on one's psychological state, especial on sleep quality.
In the present study. lowlanders also had a lower arterial oxygen saturation level as compared to the Native Tibetan group, despite having a median of 20 months of deployment at high altitude. According to our understanding, long-term exposure to high altitude is thought to maintain a higher oxygen saturation to help compensate for hypoxia in lowlanders. However, lowlanders had higher ventilatory sensitivity to CO2 in response to hypoxia as opposed to decreasing their ventilatory recruitment threshold like highlanders (Slessarev et al., 2010). The mechanism underlying ventilatory acclimatization to altitude or hypoxia has been suggested as a progressive increase in ventilation and Pa
A more recent study has also shown that after 10 days at 3850 m, lowlanders had a greater ventilatory response to CO2 as compared to Native Tibetans (Slessarev et al., 2010), but after several months of high altitude exposure, those lowlanders experience a lower Pa
Intriguingly, Tibetans residing at 3658 m have greater hypoxic ventilatory responses than do partially acclimatized Han who came to 3658 m as children (Zhuang et al., 1993). This suggests that the environmental stress of severe lifelong exposure to high altitude may have helped the Native Tibetans to evolve better biological adaptations to non-chemoreflex drives to breathe, and the ability to compensate for diminished arterial oxygen saturation at high altitude, which has been shown to vary markedly among human populations (Gallagher, 2004). Native Tibetans have likely developed these biological adaptations to allow for higher maximum oxygen uptake, greater ventilation, greater hypoxic ventilatory responses, and larger lung capacity (Wu et al., 2005), all of which will help these people to maintain better arterial oxygen saturation.
We believe that this explains, and may contribute to, the phenomenon that native Tibetans are genetically predisposed to adequate oxygenation and that this capacity for better arterial oxygen satuaration confers resistance to the hypoxia environmental stressors (Niermeyer et al., 1995; Peng et al., 2011). Together, these data suggest the lower arterial oxygen saturation in the lowlanders as compared to the Native Tibetans that we observed in our study may be due to both genetic variations and a lower hypoxic ventilatory response driven by CO2.
The multiple linear regression analysis in the current study revealed that hypoxemia and difficulty in sleep induction were the main sleep disturbances at high altitude, as reflected by the correlation of prolonged SOL, non-REM latency, decreased nocturnal Sp
However, our findings do agree with the results from a previous study where subjects with severe hypoxemia had a higher component score of PSQI in sleep latency (Kong et al., 2011). In addition, a relatively low nocturnal temperature in the study area (about 6°C in the unheated room) may also contribute to the prolonged time spent in sleep induction to some extent as it was suggested that sleep-onset was accelerated by warm temperature and correlated to increased foot temperature for adults (Raymann et al., 2007).
It is important to understand that this study has some key limitations. First, the altitude used in this study may not be high enough to clarify the discrepancy of sleep architecture between Native Tibetans and partially acclimatized lowlanders. Second, the enrolled cases were all young male participants that may inevitably result in a selection bias. However, few people from the lowlands have ever lived at an extremely high altitude for an extended period of time, let alone older males or females from the lowlands, making the possibility of broadening the cohort difficult. Finally, the possible imbalance between Native Tibetans and lowlanders among the excluded participants might have introduced a bias. Conclusions need to be confirmed by future study; however, given the relatively large sample size that received PSG at a high altitude, and the statistical significance of the observed effects in the present study, these limitations are unlikely to compromise the results.
Conclusions
The present study determined that for young male soldiers stationed at an altitude of 3800 m, partially acclimatized lowlanders and Native Tibetans demonstrated largely similar sleep architecture as determined by PSG in a very low room temperature. However, lowlanders suffered less time in non-REM sleep and had lower arterial oxygen saturation than Native Tibetans. Hypoxemia, difficulty in sleep induction, and time in deep sleep were associated with poor subjective sleep quality.
Footnotes
Acknowledgments
This research was supported by the Special Project of the “Eleventh Five-year Plan” for Medical Science Development of the People's Liberation Army (No. 06MA141).
Author Disclosure Statement
No competing financial interests exist.