
Abstract
Abstract
Masschelein, Evi, Joke Puype, Siacia Broos, Ruud Van Thienen, Louise Deldicque, Diether Lambrechts, Peter Hespel, and Martine Thomis. A genetic predisposition score associates with reduced aerobic capacity in response to acute normobaric hypoxia in lowlanders. High Alt Med Biol 16:34–42, 2015—Given the high inter-individual variability in the sensitivity to high altitude, we hypothesize the presence of underlying genetic factors. The aim of this study was to construct a genetic predisposition score based on previously identified high-altitude gene variants to explain the inter-individual variation in the reduced maximal O2 uptake (ΔV
Introduction
A
The high inter-individual variability in the sensitivity to high altitude drives research to search for genetic factors underlying these differences. Twin studies estimated the presence of genotype-environment (hypoxia) interaction or the heritability of hypoxia-induced responses in lowlanders [e.g., sensitivity to high-altitude illness or hypoxic ventilatory response (MacLeod et al., 2013)]. Recently, our group found the hypoxia-induced change in maximal oxygen uptake (V
To date, research has primarily focused on identifying genetic variants underlying differential susceptibility to high-altitude illness. Variants in 17 genes, such as nitric oxide 3 (NOS3), b2-adrenergic receptor (ADRB2), angiotensinogen (AGT), aldosterone synthase (CYP11B2), and heat-shock protein genes (HSP), have shown some association with high-altitude illness (MacInnis et al., 2010; Martin et al., 2010; Stobdan et al., 2008). In contrast to studies on the adaptations of lowlanders to high altitude, studies on highlanders have revealed many new insights. Epidemiological studies made in Tibetan, Ethiopian, and Andean natives report major population-specific differences in adaptive mechanisms to protect against hypoxia in their highland habitat (Beall, 2006; Beall, 2007). Genome-wide association studies (GWAS) of Tibetans revealed several variants in genes, including Endothelial PAS domain-containing protein 1 (EPAS1), Egl nine homolog 1 (EGLN1), and peroxisome proliferator-activated receptor alpha (PPARA) (Beall et al., 2010; Hanaoka et al., 2012; Jeong et al., 2014; Peng et al., 2011; Petousi and Robbins, 2014; Simonson et al., 2010; Yi et al., 2010).
Phenotypes related to high-altitude adaptation are multifactorial in nature and many genetic variants with most probably small effects contribute to the overall phenotype. Moreover, the common weakness of high-altitude studies is that they are based on small sample sizes, which are insufficient to detect genotype-phenotype associations with small effect sizes (Bouchard, 2011). Therefore, the development of a genetic predisposition score (GPS) offers one approach to study the effect of a set of single nucleotide polymorphisms (SNPs) simultaneously, for which a larger overall effect size may be expected.
To the best of our knowledge, the GPS approach has not been used to explain the inter-individual variability in the physiological responses to hypoxia. Therefore, the aim of this study was to define a GPS based on previously identified gene variants for different high-altitude phenotypes (high-altitude illness, gene variants in high-altitude populations, genes within hypoxia-signaling pathways) to explain the inter-individual variation in two phenotypes: 1) aerobic capacity (V
Materials and Methods
Study population
Ninety-six healthy young male Belgian lowlanders (<300 m altitude) were included in this study. Subjects included in this study were selected from different earlier studies in our laboratories (Masschelein et al., 2012; 2014; and unpublished data), but genotypic data presented here are original. All aforementioned studies (Masschelein et al., 2012; 2014; and unpublished data) were approved by the local ethics committee and were in accordance with the Declaration of Helsinki. In total, the study cohort consisted of 96 healthy male subjects. Inclusion criteria were (1) non-smoking, (2) no history of cardiovascular or respiratory disease, and (3) no residence at>1.500 m during 6 months prior to the study. All subjects gave written consent following a medical examination and after being informed of all experimental procedures.
Exercise testing
All subjects performed a maximal incremental exercise test to volitional exhaustion on a cycle ergometer (Avantronic Cyclus II, Leipzig, Germany) in both normoxia and acute normobaric hypoxia. As indicated above, subjects were selected from three different studies in which slightly different exercise protocols and hypoxic conditions were used. In study 1 (S1, unpublished data) subjects (n=61) initial workload was set at 60 W and was increased until exhaustion by 35 W every 3 min. In study 2 (n=20, S2, Masschelein et al., 2014) and study 3 (n=15, S3, Masschelein et al., 2012), initial workload was set at 50 W and was increased by 20 W per min. These two protocols are expected to give a similar maximal oxygen uptake (V
Genotyping
Anonymously coded blood samples were drawn from each subject. We genotyped 40 SNPs using a matrix-assisted laser desorption ionization time-of-flight (MALI-TOF) mass spectrometry iPLEX Gold platform (Sequenom Inc., San Diego, CA, USA). We assessed the angiotensin-converting enzyme (ACE) rs4341 SNP, using TaqMan SNP Genotyping Assay (#4351379, Life Technologies, Gent, Belgium), as a marker for ACE Insertion(I)/Deletion(D) polymorphism because it is previously found that these two polymorphisms are in linkage disequilibrium (Glenn et al., 2009; Tanaka et al., 2003). We assessed 41 SNPs in 21 genes potentially relevant for high-altitude tolerance or aerobic capacity. We chose these polymorphisms on the basis of evidence from previously reported associations and with reported minor allele frequencies>5% (Supplementary Table S1; supplementary material is available online at www.liebertpub.com/ham).
Statistical analysis
Data were analyzed using SAS Enterprise Guide 4.3 (SAS Institute Inc, Cary, NC). To test whether the observed genotype frequencies were in the Hardy-Weinberg equilibrium a χ2 test with one degree of freedom was used. To model for the three different hypoxic protocol groups, we introduced two dummy variables. Comparisons between the different study samples for age, height, weight, V
Genetic predisposition score
A genetic predisposition score (GPS) was calculated to study multiple contributing gene variants simultaneously. This approach reflects the polygenic nature of the complex hypoxia-response phenotype under study and lowers the chance of false negative findings. Individual SNPs were recoded as 0, 1, and 2 according to the number of protective-hypoxic-adaptor alleles for that particular SNP. The definition of a protective allele was based on the original association findings as the allele protecting against high-altitude sickness, or the allele most frequently present in high-altitude populations, etc. (for original references see Supplementary Table S1).
A stepwise regression analysis was first applied to detect a subset of SNPs to be associated with the two phenotypes of interest (V
Receiver operating characteristic curve
Two subgroups were made according to the hypoxia-induced ΔV
Results
Descriptive statistics
Descriptive statistics of the three different study cohorts are presented in Table 1. No significant differences in the three study groups for weight, height, BMI, and V
Data are presented as means±SE. *p<0.05, different from study group N=61
Genotyping
Supplementary Table S1 shows the 41 SNPs in 21 genes selected for genotyping. Two SNPs with a genotype efficiency <95% were excluded from the analysis. Four SNPs were flagged as being out of Hardy-Weinberg equilibrium (p<0.01) and were removed from further analyses. Supplementary Table S2 shows the single-SNP association analyses. Significant associations for V
Genetic predisposition score
Stepwise regression analysis revealed subsets of SNPs yielding significant association with V
Figure 1 and 2 show, respectively, the ANCOVA analyses of the GPS influence on V

Genetic predisposition score for V

Genetic predisposition score for the drop in V
d, dummy variable; GPS, genetic predisposition score; r, partial correlation.
Receiver operation characteristic analysis
Next, we evaluated the ability of the GPS to discriminate between the low- versus high- ΔV
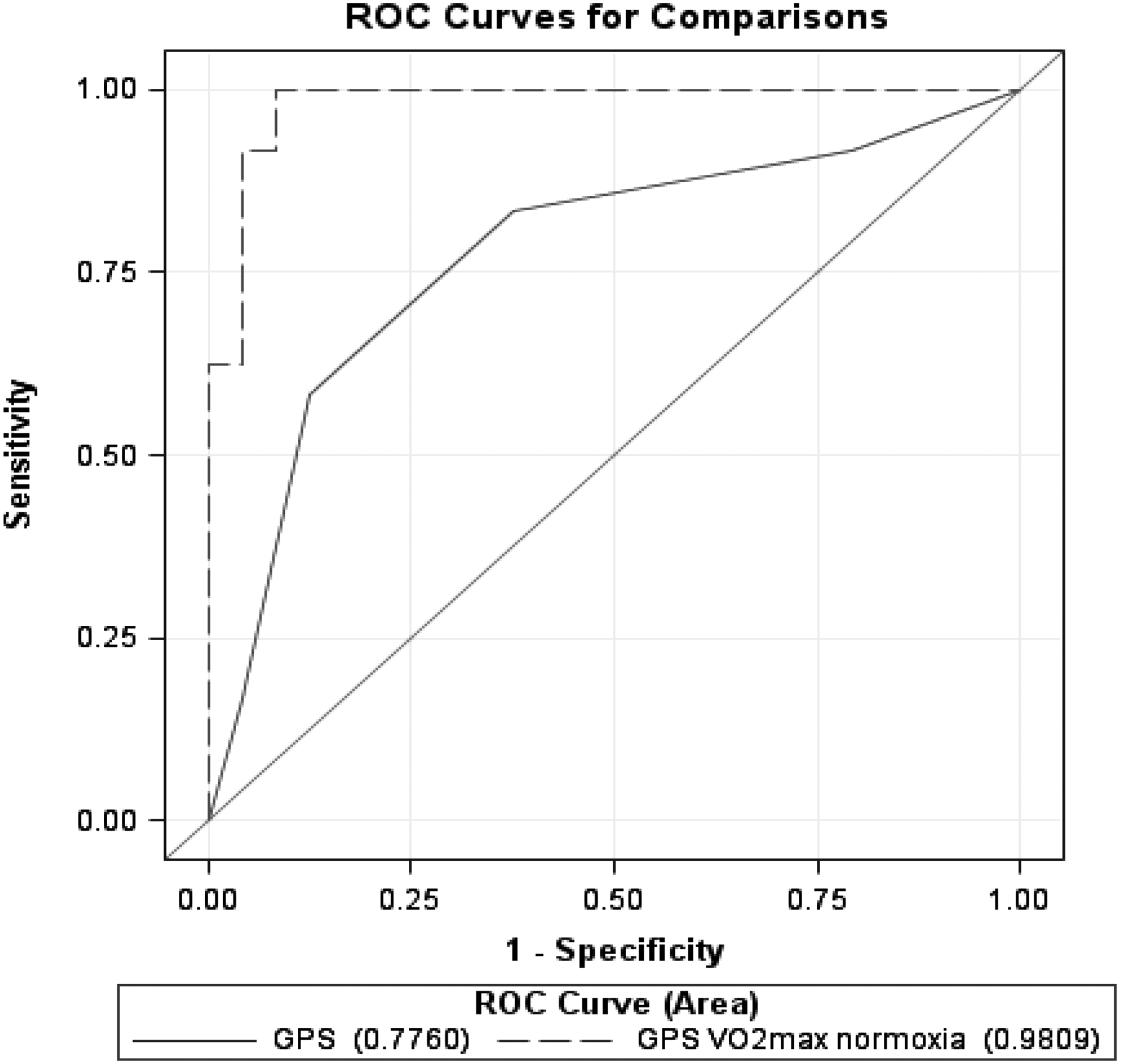
Two different models to predict low- versus high- ΔV
Discussion
Aerobic working capacity in lowlanders is impaired at high altitude. However, there is a considerable individual variation found in high-altitude sensitivity. This study aimed to contribute to the understanding of the genetic etiology of high-altitude sensitivity. The main goal of the study was to quantify the degree of explained variance of a genetic predisposition score for the decrease in maximal aerobic capacity in response to hypoxia (hypoxia-induced ΔV
Most studies report data on just one polymorphism and explained variance is relatively small. In this regard, the combined influence of several gene variants is likely to explain a larger amount of individual variability in the phenotype of interest. Recent research showed the possibility of polygenic profiles for athletic performance, as proposed by studies of Ruiz et al. (2009), Santiago et al. (2010), and Williams and Folland (2008), or for responses to endurance training (Bouchard et al., 2011) or cardiorespiratory rehabilitation (Thomaes et al., 2013). To the best of our knowledge, apart from some genetic–epidemiological observations regarding altitude illness and genetic selection in high-altitude populations, data from well-controlled hypoxia intervention studies to prove genotype-dependence of the diminished aerobic capacity in response to hypoxia are lacking. However, there is one study in lowlanders demonstrating an association between genetic markers used to discriminate ancestry and V
In calculating a GPS based on this subset of SNPs, every polymorphism was considered to have an equal influence on the phenotype, which is a simplified approach compared to an effect-size weighting approach (Belsky et al., 2013; Janssens et al., 2007). Our main finding was that within the selected set of 35 gene variants, based on a stepwise multiple regression analysis, we were able to identify six polymorphisms [rs6735530 (EPAS1), rs4253778 (PPARA), rs833070 (VEGFA), rs4341 (ACE), rs1042713 (ADRB2), and rs1042714 (ADRB2)], to be associated with the decrease in maximal aerobic capacity in response to hypoxia.
Polymorphisms in the EPAS1 and PPARA gene have recently been selected as candidate genes in genome-wide association studies involved in the evolutionary adaptation in the high-altitude Tibetan population (Beall et al., 2010; Hanaoka et al., 2012; Jeong et al., 2014; Peng et al., 2011; Petousi and Robbins, 2014; Simonson et al., 2010; Yi et al., 2010). Both genes are implicated in the hypoxia-inducible factor (HIF) oxygen signaling pathway. EPAS1, which encodes HIF-2α, is an element of the transcription factor HIF2 that seems to be the primary mediator of erythropoietin expression in kidneys in response to hypoxia (Kapitsinou et al., 2010). A mutation in this gene was previously associated with polycytosis (Percy et al., 2008) and, interestingly, several gene variants in this gene were found to be associated with lower hemoglobin levels amongst Tibetans adapted to living at high altitude (Beall et al., 2010). This suggests that Tibetans have evolved a blunted erythropoietic response to hypoxia. Simultaneously, variants of PPARA were also associated with lower hemoglobin levels in Tibetans (Simonson et al., 2010). PPARA encodes for a major transcriptional regulator of lipid metabolism, yet recently has been considered to be one of the candidate genes for high-altitude adaptation in Tibetans (Simonson et al., 2010). PPARA has been shown to interact with the HIF-pathway [e.g., a HIF-dependent downregulation of PPAR expression was found during hypoxia in mice (Narravula and Colgan, 2001)]. In addition, the antidiabetic agent tesaglitazar, a PPARA agonist, decreased hemoglobin levels in patients with type 2 diabetes (Wilding et al., 2007). Besides the involvement of these genes in regulating erythropoiesis, they might also play a role in regulating energy metabolism (Ge et al., 2012).
The EPAS1 haplotype that was previously associated with decreased hemoglobin concentration, was also highly associated with increased lactate concentration and the putatively advantageous PPARA haplotype was correlated with decreased activity of fatty acid oxidation (Ge et al., 2012). The current study for the first time demonstrates the importance of EPAS1 and PPARA for hypoxia-induced drop of maximal aerobic capacity in a lowland population during acute exposure to hypoxia.
VEGFA is a key factor involved in angiogenesis and vascular permeability, and is upregulated in hypoxia (D'Hulst et al., 2013; Levy et al., 1995). Based on the known function of VEGFA in hypoxic conditions, several studies have assessed the function of VEGFA in the pathogenesis of high-altitude illness (Buroker et al., 2012; Ding et al., 2011; 2012; Hanaoka et al., 2009). The haplotype (rs1413711, rs833070, and rs3025000) in the VEGFA gene has previously been shown to be associated with acute mountain sickness (AMS) (Ding et al., 2012). Here we report for the first time a significant contribution of a VEGFA polymorphism (rs833070) to the inter-individual variation in diminished maximal aerobic capacity in response to hypoxia.
The β2-adrenergic receptor (ADRB2) is a G-protein coupled cell surface epinephrine receptor that acts as the primary epinephrine receptor in the lungs. At high altitude, the epinephrine response is mainly mediated through the activation of β2-adrenergic receptors facilitating oxygen delivery (Mazzeo and Reeves, 2003). A recent study has indicated an association between polymorphisms in the ADRB2 gene (rs1042713 and rs1042714) and high altitude pulmonary edema in Indian highland population (Stobdan et al., 2010). Furthermore, there is suggestive evidence that the ADRB2 gene may be associated with endurance performance (Wolfarth et al., 2007; Sarpeshkar and Bentley, 2010). We demonstrated a link between the polymorphisms rs1042713 and rs104271 and the decreased aerobic capacity in response to acute hypoxia in lowlanders. Many studies have investigated the insertion (I)/deletion (D) polymorphism of the ACE gene (ACE I/D polymorphism) in relation to lowlanders' adaptation to high altitude. Studies showed the I allele to be associated with aspects of endurance performance, with higher observed frequencies in elite distance runners (Meyerson et al., 1999) and mountaineers (Montgomery et al. 1998; Tsianos et al., 2005). The I allele has been reported to be advantageous in maintaining higher arterial oxygen (Woods et al., 2002). We demonstrated an association between the ACE I/D polymorphism and hypoxia-induced ΔV
Based on the aforementioned six polymorphisms, a GPS model was created. The GPS model (Fig. 2) was significantly associated and a higher number of ‘increasing/protective alleles’ resulted in a better hypoxic tolerance (i.e., smaller decrease in maximal aerobic capacity in response to hypoxia). Furthermore, GPS was found to be an independent predictive variable with a partial explained variance (R2) of 23% (p<0.0001, Table 2). For each additional ‘protective’ allele, the drop in hypoxia-induced ΔV
Additionally to the primary purpose of this study, we also examined the influence of our set of 35 genetic variants on V
Our findings are exploratory in nature, given the small available sample and the use of three slightly different hypoxia exposure protocols. Also the pre-set selection or ‘snapshot’ of selected gene variants among the various polymorphisms can potentially affect altitude sensitivity. We are aware that the selection of gene variants may have left out other SNPs that might be associated with aerobic capacity in hypoxia. Furthermore, we assumed allelic effects to be co-dominant, as well as each SNP to have an equal additional effect. Our model does not take into account potential complex interactions between gene variants. The significance of a GPS based on previously identified gene variants for different high-altitude phenotypes (e.g. high-altitude illness) was tested in predicting the diminished maximal aerobic capacity in response to hypoxia. Although there is no formal evidence of pleiotropic gene actions related to high-altitude illness and reduced exercise capacity at altitude, common hypoxia-induced physiological pathways might be active in both phenotypes.
We observed a 11% larger drop in ΔV
Conclusion
In conclusion, we found that a genetic predisposition score, composed of six polymorphisms, was significantly associated with a diminished maximal aerobic capacity in response to acute hypoxia. The GPS explained 23% of the overall variance in hypoxia-induced ΔV
Footnotes
Acknowledgments
The authors thank Monique Ramaekers for skillful technical assistance during the experiments. The participation of the subjects is also gratefully acknowledged.
Author Disclosure Statement
The authors do not have any financial conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.