
Abstract
Abstract
DiPasquale, Dana M., Fred W. Kolkhorst, and Michael J. Buono. Acute normobaric hypoxia reduces body temperature in humans. High Alt Med Biol 16:61–66, 2015.— Anapyrexia is the regulated decrease in body temperature during acute exposure to hypoxia. This study examined resting rectal temperature (Trec) in adult humans during acute normobaric hypoxia (NH). Ten subjects breathed air consisting of 21% (NN), 14% (NH14), and 12% oxygen (NH12) for 30 min each in thermoneutral conditions while Trec and blood oxygen saturation (Sp
Introduction
A
Although reduced body temperature in hypoxia has been widely observed among animals, including higher orders such as simian primates (Tattersall and others, 2002) and human newborns (Cross and others, 1958), its presence in human adults is much less clear. Previous findings have suggested the presence of hypoxia-dependent temperature changes in human adults. Early observations indicated that rectal temperature decreased after 40 min in variable normobaric hypoxia (NH) (as low as 10%); however, this was in slightly cool conditions, 19°C, and no statistical analysis was performed (Kottke, 1948). It can be calculated from the Kolka et al. data (1987) that resting and exercising esophageal temperature in hypobaric hypoxia (HH) was ∼0.3°C lower, and skin temperature, sweating, and metabolic heat production were all reduced compared to normobaric normoxia (NN). While lowered Tb has been observed by others (Cipriano and Goldman, 1975), paradoxically, Tb has also been shown to be both higher (Coste and others, 2004; 2009) and not different in HH than in NN (Cipriano and Goldman, 1975; Miyagawa and others, 2011). Those who found increased Tb examined the role of an 8-h exposure on 24-h mean Tb. It is therefore possible, that these increases were associated with pathophysiological changes in acute mountain sickness (AMS), since elevated Tb correlates with AMS severity in NH (Richardson and others, 2008).
Other research in NH, in which AMS was not intentionally induced, has shown a high degree of variability ranging from no difference (Golja and Mekjavic, 2003; Malanda and others, 2008; Miyagawa and others, 2011; Simmons and others, 2011) to a lowered Tb (Robinson and Haymes, 1990; Richardson and others, 2009) compared to NN. The mixed results are likely because these studies had very different main purposes other than examining Tb changes related to hypoxia, and as such, they are complicated by water immersion baths, hypo- and hyperhydration, ambient temperature variations (e.g., heat, cold), and altitude acclimation. Consequently, methods were not ideal to test the basic question—is Tb reduced in hypoxia in adult humans? We therefore sought to contribute to this small body of research by addressing that question. We hypothesized that acute hypoxia exposure would result in lowered rectal temperature (Trec) and that this effect would be graded with the severity of deoxygenation.
Methods
After giving written informed consent, six men and four women (29±10 y; 175±13 cm; 69.9±17.6 kg; mean±SD) participated in the study, which was approved by the university's institutional review board. Subjects refrained from exercise on the day of testing and, to ensure euhydration, were instructed to drink at least 2 L of water during the 24-h before testing. All testing was performed with the subjects clothed in pants and a shirt and in an environmental chamber in which ambient temperature was maintained at 25°C and relative humidity at 40%.
Subjects remained in a resting semi-recumbent position throughout testing. During a single testing day, subjects were exposed, in order, to NN (FI
We fit multiple linear regression models with cluster adjusted standard errors (to account for the repeated measures) using NN, NH14, andNH12 as indicator variables, and Trec or Sp
Results
As expected, regression analysis indicated that mean Sp
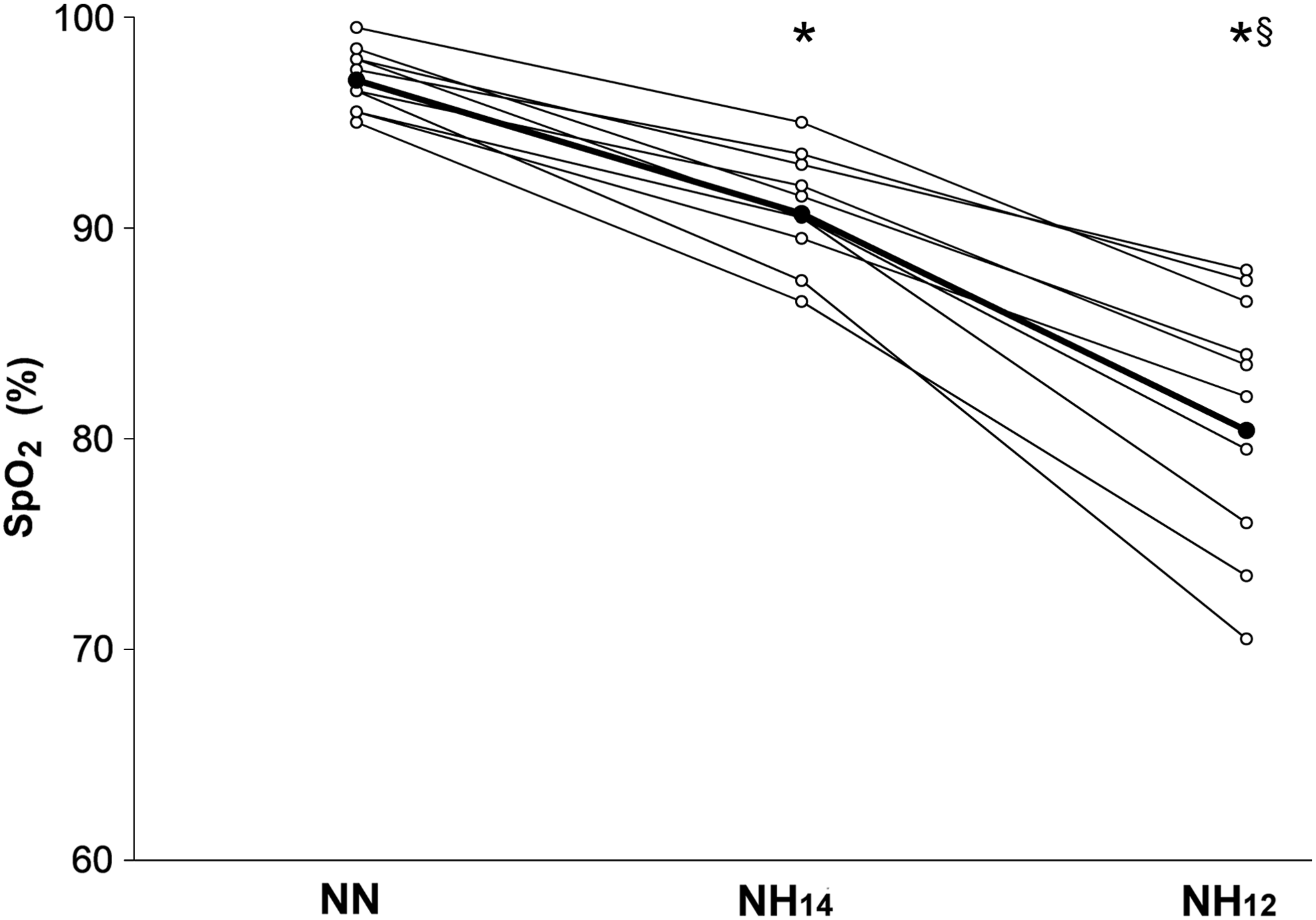
Individual responses of acute normobaric hypoxia on peripheral oxygen saturation (Sp
Trec was reduced in hypoxia, and this effect was graded with the severity of hypoxia. Mean Trec was 0.13°C lower during NH14 (p=0.014) and 0.25°C lower in NH12 compared to NN (p=0.0001). In NH10, Trec decreased by an average of 0.30°C compared to NN. The difference in Trec between NH14 and NH12 was also significant (χ2(1)=4.79, p=0.0287). Figure 2 illustrates the individual and mean responses of Trec.
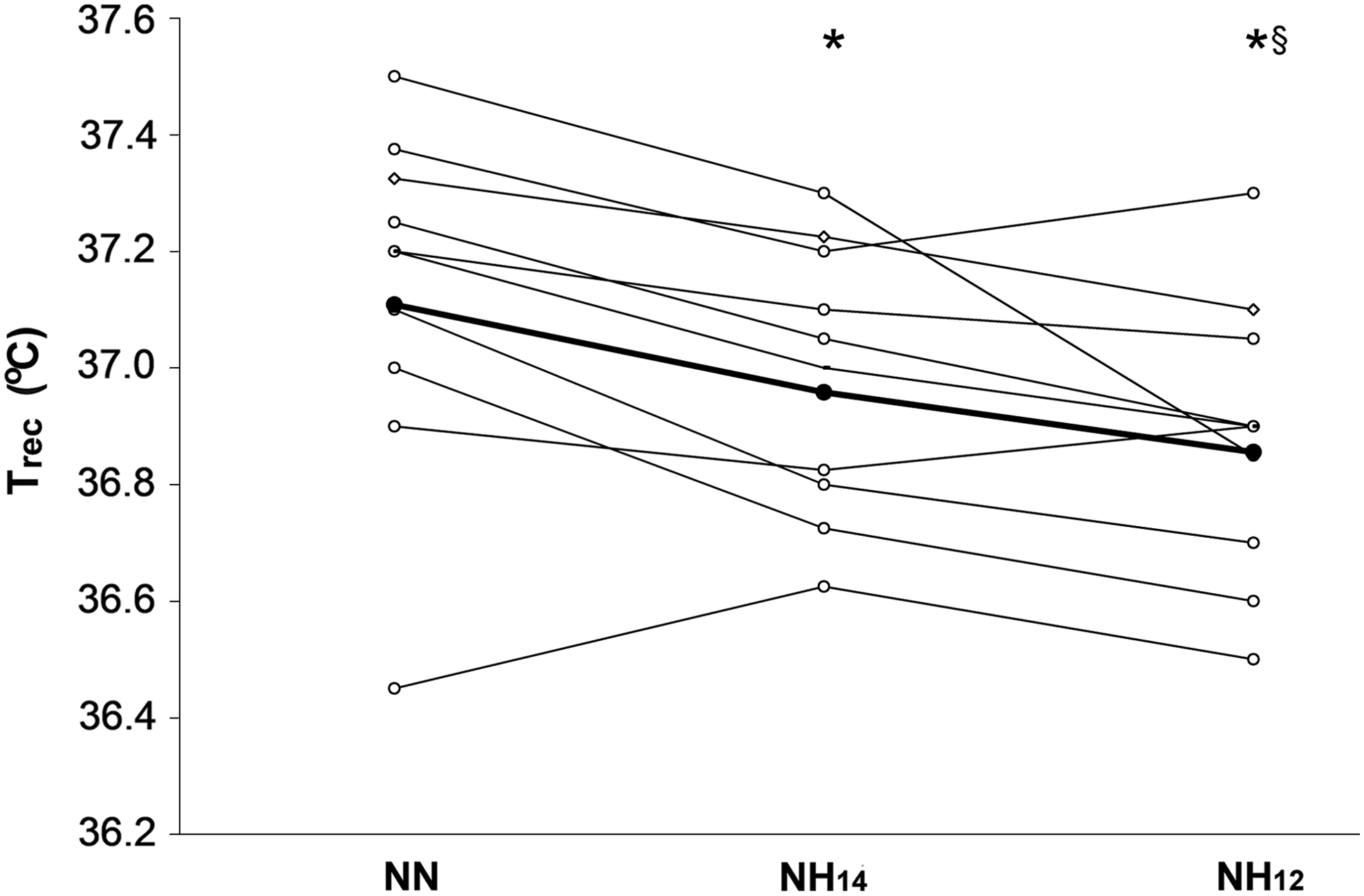
Individual responses of hypoxia on rectal temperature (Trec). The mean response is indicated by the heavy line. During exposure to NH14 and NH12, Trec decreased from that at NN, and they were different from each other. *Significantly different than NN (p=0.0001). §Significantly different than NH14 (χ2(1)=4.79, p=0.0287).
Sp

Individual relationships between rectal temperature (Trec) and peripheral oxygen saturation (Sp
Discussion
In this investigation, hypoxia reduced Trec by as much as 0.30°C. Normal body temperature is usually maintained within fairly narrow limits of ∼36.1–37.7°C (Marieb, 1992) to allow essential enzymatic reactions to occur. Temperatures above 43°C typically lead to death (Hubbard, 1990) and below 34°C lead to abnormal cardiac function such as arrhythmia (Aslam and others, 2006). Therefore, even a small change in Tb can be clinically important. Our findings demonstrate that, in addition to unicellular organisms, lower vertebrates, and human newborns ( Cross and others, 1958; Malvin and Wood, 1992; Branco and Malvin, 1996), a hypoxia-induced reduction in Tb is present in adult humans. Whether the present investigation demonstrates the regulated change in thermal set-point (i.e., anapyrexia) or peripheral responses to hypoxia was not determined. We speculate that, given the very short exposures, the results were not the effect of resetting the set-point, but given longer exposures, they could be (Boutilier, 2001).
Lowered Tb in hypoxia is the result of two hypoxia-induced changes: 1) hypometabolism and 2) direct action on the thermoregulatory brain centers. Hypometabolism is triggered by hypoxia and is considered a regulated change (Mortola 1993; 2004) that conserves energy by reducing oxygen consuming processes like maintaining muscle tone, cell excitation, growth, and heat production. Research suggests that hypoxia-induced hypometabolism precedes and likely causes the drop in Tb (Pedraz and Mortola 1991; Gautier and Bonora 1992; 1994; Giesbrecht and others, 1994). Several putative mechanisms of hypometabolism in hypoxia have been suggested, though they are largely conjectural. For example, oxygen may serve as a regulatory molecule (Hochachka, 1996; Jones, 1996) by being sensed by cytosolic oxygen-dependent enzymes (Mortola, 2004) and thus, directly mediating metabolism at a cellular level. Others have suggested that arginine vasopressin, which is stimulated by hypoxia, could be also be a mediator of hypometabolism (Wood, 1991). There is evidence of a decrease in the uncoupling protein and blood perfusion of brown adipose tissue, suggesting that reduced metabolism of this fat in turn lowers nonshivering thermogenesis (Mortola and Naso, 1997; Mortola and others, 1999). Conversely, observations show that peripheral chemoreceptors are not involved in the hypometabolic response to hypoxia (Gautier and others, 1987; 1989; Gautier and Bonora, 1992).
The second factor contributing to the lowered Tb is direct action on the central nervous system. It has been suggested that activation of the serotonin-cAMP and NO-cGMP in the preoptic region of the hypothalamus is responsible for lowered Tb in hypoxia (Steiner and others, 2002; Steiner and Branco, 2003). More recently, the transient receptor potential vanilloid 1 (TRPV1) ion channel, which plays a role in detection and regulation of body temperature and provides sensation of scalding heat and pain (Gavva and others, 2007), may act in concert with hypoxia via activation of hypoxia-inducible factor-1 alpha (Yuen and others, 2012). TRPV1 is predominantly in the central nervous system but is also present in skeletal muscle where it acts on metabolism (Luo and others, 2012). This suggests a possible dual role of TRPV1 in lowering Tb in hypoxia, although this remains to be determined.
Other possible modulators of Tb in hypoxia such as amino acids, peptides, nucleotides, endogenous opioids, and gas molecules (e.g., carbon monoxide, nitric oxide) have been hypothesized (Steiner and Branco, 2002). Peripheral heat loss contributors, for example, sweat rate (Kolka and others, 1987; DiPasquale and others, 2002), skin blood flow (Baily and others, 1991; Kolka and others, 1987; Rowell and others, 1982; Tattersall and Milsom, 2003; Wagner and others, 1980), and hyperpnea (Mortola, 1996), do not account for the lowering of Tb. Although the mechanisms of hypometabolism and thermal resetting in hypoxia are unclear, both responses help reinforce the other. Decreased metabolic rate lowers heat production and thus, Tb, while reduced Tb helps keep metabolism depressed by reducing ideal temperatures for enzymatic processes.
Hypoxia-induced reduction of Tb appears to be an evolutionarily conserved process that serves a protective role. In fact, this has been demonstrated clinically. For example, treatment with mild hypothermia after cardiopulmonary resuscitation has become the standard of care (Peberdy and others, 2010) since it improves neurologic and cognitive function in comatose survivors of cardiac arrest (Bernard and others, 2002). Therapeutic hypothermia has also been beneficial in improving survival outcomes for infants born with oxygen deficiencies (Shankaran and others, 2005; 2012). Hypoxic Tb reduction has been shown to prevent apoptotic cell death (Zhu and others, 2004) and preserve cognitive functioning after severe hemorrhage (Alam and others, 2005) in animals. Hypoxic Tb depression also causes a leftward shift in the oxyhemoglobin dissociation curve, resulting in greater oxygen loading in the blood (Lister, 1984). This may be of benefit for oxygen-sensitive organs such as the heart and brain (Wood, 1991). Furthermore, the protective effect of reduced Tb has been observed in increased survival rates of animals whose body temperature decreased during hypoxia exposure (Miller and Miller, 1966). These observations demonstrate the therapeutic benefit of lower body temperature, which suggests that reduced Tb is an adaptive response to hypoxia.
It is interesting to note that AMS severity is strongly associated with higher Tb in both NH and HH (Maggiorini and others, 1997; Richardson and others, 2008). AMS—a maladaptive response to hypoxia—is a symptom complex with headache, anorexia, nausea, vomiting, insomnia, lassitude, and/or malaise (Muza, 2007). While AMS is usually self-limiting, symptoms can progress to life-threatening high altitude cerebral or pulmonary edema. AMS is also characterized by changes in affect, mood, and cognitive function (Shukitt-Hale and others, 1991). However, the pathophysiology remains unclear despite decades of research. Traditionally, AMS has been thought to be primarily the result of hypoxia and can occur in both hypobaric hypoxia (HH) and normobaric hypoxia (NH) (Richard and others, 2014). Perhaps part of AMS pathophysiology is the absence of a protective lowering of Tb. This is supported by the finding that AMS is associated with elevated Tb (Maggiorini and others, 1997; Richardson and others, 2008). Thus, anapyrexia, or lack thereof, may be a predictor of AMS susceptibility. This warrants further investigation.
We recognize that this study has limitations. First, there was a possible order effect since subjects were exposed to the graded levels of hypoxia in the same order. Second, it is possible that since exposures were completed back-to-back, the physiological changes in one condition could bleed into the next. Pilot testing indicated that over the course of 90 min, Trec only varied by 0.05°C after 15–20 min. Hence, each subject was exposed to conditions in 30 min increments when Trec appeared stable. There is, of course, the possibility that Trec may change after 30 min, as an increase in the 24-h circadian rhythm of body temperature has been observed after an 8 h exposure to altitude (Coste and others, 2004; 2009). However, others have shown that Trec stabilized by 30 min in hypoxia with similar thermal conditions as ours (Cipriano and Goldman, 1975). Additionally, others have used similar time frames (Richardson and others,2009) or have shown that the largest changes in body temperature occurred within 30 min (Robinson and Haymes, 1990). Subjects participating in longer exposures run the risk of developing AMS (Richardson and others, 2008), with some developing symptoms as soon as 1 h into the exposure and illness by 6 h (Loeppky and others, 2008). Therefore, longer exposures may be confounded by the pathophysiological process of AMS. On the other hand, multiple exposures may be confounded by adaptive responses such as observed after acclimatization. It is unclear how long such changes persist, making it difficult to choose a time point for a second exposure. Therefore, short exposures completed in a single day avoid the influence of AMS, avoid acclimatization effects, avoid circadian variations when testing on different days, and, at worst, underestimate effects by being too short for body temperature to stabilize.
At best, though, these data show the initial thermoregulatory changes encountered in hypoxia exposure, an area that receives little attention but which could shed light on normal and pathophysiological responses to hypoxia. We observed a decrease in Trec with acute exposure to hypoxia. This supports the possibility of anapyrexia in adult humans and confirms that, along with many other animals, humans respond to acute hypoxia by lowering temperature. However, the neural and cellular signaling pathways that control anapyrexia, anapyrexia's role in AMS, and if there are differences in anapyrexia in HH and NH, are unclear and certainly warrant further study.
Footnotes
Author Disclosure Statement
No competing financial interests exist.