
Abstract
Abstract
Saxena, Saurabh, Dhananjay Shukla, and Anju Bansal. Expression of monocarboxylate transporter isoforms in rat skeletal muscle under hypoxic preconditioning and endurance training. High Alt Med Biol 17:32–42, 2016—Previously, we have reported the regulation of monocarboxylate transporters (MCT)1 and MCT4 by physiological stimuli such as hypoxia and exercise. In the present study, we have evaluated the effect of hypoxic preconditioning and training on expression of different MCT isoforms in muscles. We found the increased mRNA expression of MCT1, MCT11, and MCT12 after hypoxic preconditioning with cobalt chloride and training. However, the expression of other MCT isoforms increased marginally or even reduced after hypoxic preconditioning. Only the protein expression of MCT1 increased after hypoxia preconditioning. MCT2 protein expression increased after training only and MCT4 protein expression decreased both in preconditioning and hypoxic training. Furthermore, we found decreased plasma lactate level during hypoxia preconditioning (0.74-fold), exercise (0.78-fold), and hypoxia preconditioning along with exercise (0.67-fold), which indicates increased lactate uptake by skeletal muscle. The protein–protein interactions with hypoxia inducible factor-1 and MCT isoforms were also evaluated, but no interaction was found. In conclusion, we say that almost all MCTs are expressed in red gastrocnemius muscle at the mRNA level and their expression is regulated differently under hypoxia preconditioning and exercise condition.
Introduction
T
The transport of lactate is facilitated by pH-sensitive protein-linked monocarboxylate transporters (MCTs) that are responsible for transport of many other monocarboxylates also, such as pyruvate, ketone bodies (viz., acetoacetate and β-hydroxybutyrate), and branched chain ketoacids (viz., α-ketoisocaproate formed from the transamination of leucine) (Halestrap and Meredith, 2004). Currently, 14 MCT members are known (Halestrap and Wilson, 2012). Functional characterization of MCT isoforms has been extended to seven isoforms (MCT1-4, 6, 8, 10) with the seven remaining MCT family members being classified as orphan MCTs (MCT5, 7, 9, 11–14). A number of reviews and articles have examined the tissue-specific localization and physiological functions of MCT isoforms in both humans and rodents (Chidlow et al., 2005; Gill et al., 2005; Hashimoto et al., 2005; Pierre and Pellerin, 2005; Chiry et al., 2006; Halestrap, 2012; Halestrap and Wilson, 2012). Regulation of MCTs has been demonstrated to occur through transcriptional, translational, and post-transcriptional mechanisms (Cuff and Shirazi-Beechey, 2002; Enoki et al., 2006; Chenal and Pellerin, 2007). These regulatory pathways appear to be age and tissue dependent, which further complicates the understanding of these pathways (Enoki et al., 2006; Chenal and Pellerin, 2007).
MCT1 and 4 have major functions in skeletal muscle. MCT1 is widely expressed in different mammalian tissues, including oxidative skeletal muscle (Bonen et al., 2006), and is confined in muscle to sarcolemmal, subsarcolemmal, and interfibrillar compartments of oxidative fibers (Hashimoto et al., 2005). The presence of MCT1 at sarcolemmal and mitochondrial membranes facilitates uptake of lactate into muscle fiber and mitochondria, respectively (Bonen et al., 2000; Dubouchaud et al., 2000; Brooks, 2002; Butz et al., 2004; Halestrap and Wilson, 2012; Thomas et al., 2012), after which mitochondrial LDH converts it into pyruvate to allow its oxidation (Cruz et al., 2012), whereas the main function of MCT4 is cellular lactate extrusion (Dimmer et al., 2000; Halestrap and Wilson, 2012; Thomas et al., 2012) and it is prominently found in tissues, which are glycolytic in nature (e.g., white skeletal muscle). However, as working skeletal muscles can switch from net lactate release to utilization (Brooks et al., 1998) and sarcolemmal vesicle preparations respond to trans-stimulation (Brown and Brooks, 1994), MCTs have been identified to be bidirectional in function, facilitating exchange down H+ and lactate anion concentration gradients (Roth and Brooks, 1990a, 1990b).
In our previous study, we have found that the expression levels of MCT1 and MCT4, which are major MCTs in skeletal muscle, are affected by exercise and hypoxia preconditioning with cobalt chloride (i.e., CoCl2.6H2O) (Saxena et al., 2010). This finding prompted us to hypothesize that the gene expression of other MCTs in skeletal muscle also might be regulated under physiological stimuli such as hypoxia preconditioning and exercise, which might lead to altered muscle physiology and overall work performance. Therefore, we studied the mRNA expression of MCTs in skeletal muscle. We also studied the protein–protein interactions of MCT1, MCT2, and MCT4 with hypoxia inducible factor-1 (HIF1) to understand their regulation by using the coimmunoprecipitation technique.
Materials and Methods
All chemicals were purchased from Sigma-Aldrich. Antibodies were purchased from Santa Cruz Biotech.
Animals
Male Sprague-Dawley rats (170 ± 10 g) were used for all experiments. Animals were provided by the experimental animal facility of our institute. Animals were housed in a controlled environment with a 12-hour light/12-hour dark cycle and access to food (Lipton India Ltd.) and water ad libitum. Body weight and food and water intake were measured daily. Since three rats were housed per cage, average food and water intake per individual rat was estimated from the total food and water intake per cage. All animal procedures were approved by the Institutional Animal Ethics Committee and the guidelines of Universities Federation for Animal Welfare (UFAW) for animal research were followed. In our previous study (Saxena et al., 2010), we have reported that oral supplementation of 10 mg/kg body weight CoCl2.6H2O for 15 days significantly augments swimming time till exhaustion in rats. Therefore, all the experiments were conducted with the same dosage of cobalt chloride.
Exercise (training) protocol and cobalt supplementation
An experiment with 32 rats was set, which were divided into four groups (n = 8 each) according to the treatment and exercise conditions: (1) nonsupplemented sedentary (Con); (2) nonsupplemented training (Tr); (3) Co2+-supplemented sedentary (Cob); and (4) Co2+-supplemented training with the same training protocol (CobTr). Rats in the nonsupplemented groups were given distilled water with the help of gastric cannula, and the rats in the cobalt-supplemented group were given 10 mg/kg CoCl2.6H2O/day for 15 days 2 hours before exercise. Exercise was done by Tr and Cob+Tr groups only. In both the groups, the exercise regime was the same, that is, the rats were subjected to swimming training for 30 minutes/day and the training time was increased 10 minutes/day until it reached 1 hour (4th day). The rats were trained for 1 hour/day up to the 15th day and the training was given 6 days a week. This exercise protocol has been chosen because it has been shown to effectively increase the swimming time till exhaustion in rats in our previous studies (Saxena et al., 2010, 2012).
Muscle samples
On the 16th day, the rats were sacrificed under ketamine (80 mg/kg i.p.) anesthesia, and the whole gastrocnemius muscle was collected from the calf region of the hind limb of the rats. The red gastrocnemius muscle (100–150 mg) was then separated from the whole muscle, washed in phosphate-buffered saline (PBS) twice, snap-frozen after removal of hair and fatty tissues, and stored at −80°C for subsequent biochemical, mRNA, and protein expression studies. The red gastrocnemius muscle was chosen because it is actively recruited during swimming and is highly oxidative in nature (Delp and Duan, 1996; Gonchar, 2005). Before killing, blood samples were collected by retro-orbital puncture.
Protein expression studies
Western blot technique was used to study the protein expression of HIF1α; MCT1, 2, and 4; peroxisome proliferator-activated receptor-α (PPARα); and peroxisome proliferator-activated receptor gamma coactivator-1α (PGC1α). Muscle samples were homogenized and cytosolic and nuclear fractions were isolated by using the Nuclear/Cytosolic Fractionation Kit (Biovision) as per the manufacturer's instructions. HIF1α, PPARα, and PGC1α expression levels were studied in the nuclear fraction, whereas MCT1, 2, and 4 expression levels were examined in the cytosolic fraction. Protein concentrations were quantified by using Bradford's reagent. Fifty micrograms of protein was electrophoresed on SDS-PAGE, electroblotted onto a nitrocellulose membrane. The membrane was blocked with 5% nonfat dry milk for 1 hour, washed, and probed with respective rabbit polyclonal antibodies. The membrane was washed with PBS Tween-20 (0.1%) and incubated with anti-rabbit-IgG-HRP conjugate (1:40 000) for 2 hours. The membrane was then incubated with chemiluminescent substrate (Sigma-Aldrich) and the bands were developed using X-ray films (Kodak). All antibodies were purchased from Santa Cruz Biotech and the chemiluminescence peroxidase substrate kit was purchased from Sigma. The intensities of the bands were quantified by using ImageJ software (National Institutes of Health; freely available at http://rsb.info.nih.gov/ij/).
mRNA expression studies
The reverse transcription–polymerase chain reaction (PCR) technique was used to study mRNA expression in the muscle. Total RNA was extracted from muscle homogenates by using the Trizol method. The RNA quality and quantity were checked by both spectrophotometry and agarose gel electrophoresis. cDNA was prepared using the first-strand c-DNA synthesis kit (Fermentas) as per the manufacturer's instructions. PCR primers for various genes were purchased from Integrated DNA technologies, Inc. The details of sequences, annealing temperature, and amplicon size are represented in Table 1. The PCR was conducted using a thermal cycler (MJ Research) with the following conditions: initial denaturation was carried out at 94°C for 2 minutes, followed by denaturation at 94°C for 1 minute, annealing for 1 minute, and extension at 68°C for 1 minute for a total of 30 cycles, followed by final extension at 68°C for 10 minutes. The PCR products were electrophoresed on 2% agarose gel and visualized by UV transillumination. The images of the PCR products were acquired and the intensities of the bands were quantified by using ImageJ software (National Institutes of Health; freely available at http://rsb.info.nih.gov/ij/).
MCT, monocarboxylate transporters.
Data analysis
All the experiments were performed on three different occasions and data are presented as mean ± SD. One-way analysis of variance with post hoc Bonferroni analysis was used to determine statistical significance between groups and p < 0.05 was considered as significant.
Results
mRNA expression of MCTs
Figures 1–3 represent the mRNA expression of various MCTs with their relative optical densities. The mRNA expression of MCT1 increased both after hypoxic preconditioning with cobalt (7.34-fold, p < 0.01) and training (8.64-fold, p < 0.01). Cobalt supplementation along with training further increased the mRNA expression level of MCT1 significantly in red gastrocnemius muscle compared with control (9.77-fold, p < 0.01). Hypoxia preconditioning reduced MCT2 mRNA expression (0.45-fold, p < 0.01) significantly, but exercise training enhanced it (4.28-fold, p < 0.01) compared with control. Cobalt supplementation along with training decreased the MCT2 mRNA expression level significantly (0.49-fold, p < 0.01) compared with training only. Moreover, MCT3 mRNA expression was enhanced in response to cobalt supplementation (1.9-fold, p < 0.01) and training (1.85-fold, p < 0.01) relative to control. Cobalt supplementation along with training did not affect the expression compared with training only. MCT4 mRNA expression reduced significantly in response to hypoxia preconditioning (0.51-fold, p < 0.01) and training (0.62-fold, p < 0.01) relative to control. Cobalt supplementation along with training further decreased (0.47-fold, p < 0.01) it compared with training only. Furthermore, MCT7 mRNA expression remained unaltered after exercise training relative to control. It reduced in cobalt-supplemented (0.71-fold, p < 0.01) rats and cobalt supplemented along with training (0.74-fold, p < 0.01) rats compared with control and trained groups, respectively. Cobalt supplementation did not have any effect on MCT9 mRNA expression, but it increased in the trained group (1.53-fold, p < 0.01) compared with control. Moreover, hypoxia preconditioning with cobalt (2.12-fold, p < 0.01) and training (2.47-fold, p < 0.01) enhanced MCT10 mRNA expression in rat gastrocnemius muscle compared with control. Cobalt supplementation along with training (1.2-fold, p < 0.05) increased it relative to the training group. Furthermore, there was an increase in MCT11 mRNA expression after cobalt supplementation (2.32-fold, p < 0.01), exercise training (1.41-fold, p < 0.01), and cobalt supplementation along with training (3.58-fold, p < 0.01) compared with control. Similarly, MCT12 mRNA expression level also increased after hypoxia preconditioning with cobalt chloride (2.87-fold, p < 0.01), exercise training (1.43-fold, p < 0.01), and cobalt supplementation along with training (2.59-fold, p < 0.01) relative to control. The mRNA expression of MCT13 was lowered in cobalt-supplemented rats (0.71-fold, p < 0.01) compared with control. Trained rats had a higher level (1.77-fold, p < 0.01) of MCT13 relative to control, but cobalt supplementation along with training (1.51-fold, p < 0.01) reduced it compared with trained rats. Neither cobalt supplementation nor exercise changed MCT8 mRNA expression in red gastrocnemius muscle of rats. In addition, we did not find any expression of MCT5, MCT6, and MCT14 in rat red gastrocnemius muscle.
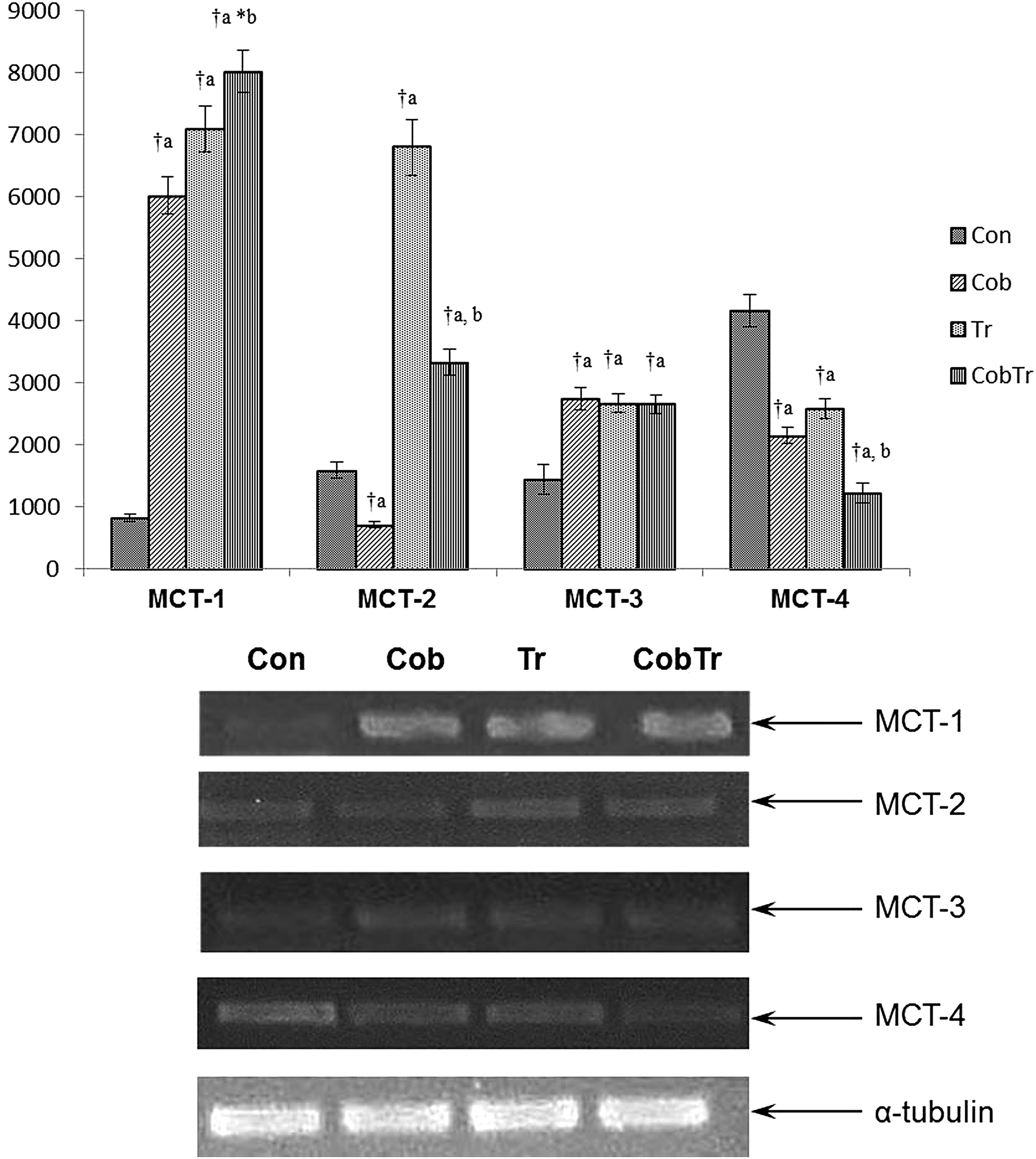
Effect of hypoxia preconditioning with cobalt chloride supplementation (10 mg/kg body weight CoCl2.6H2O for 15 days) on mRNA expression of lactate transporters (MCT1–4). Data represent the mean ± SD of three independent experiments. Data analyzed by one-way ANOVA with post hoc Bonferroni analysis. aSignificantly different from control. bSignificantly different from training. †Significantly different (p < 0.01). *Significantly different (p < 0.05). MCT, monocarboxylate transporters.

Effect of hypoxia preconditioning and exercise on mRNA expression of orphan transporters. Data represent the mean ± SD of three independent experiments. Data analyzed by one-way ANOVA with post hoc Bonferroni analysis. aSignificantly different from control. bSignificantly different from training. †Significantly different (p < 0.01).
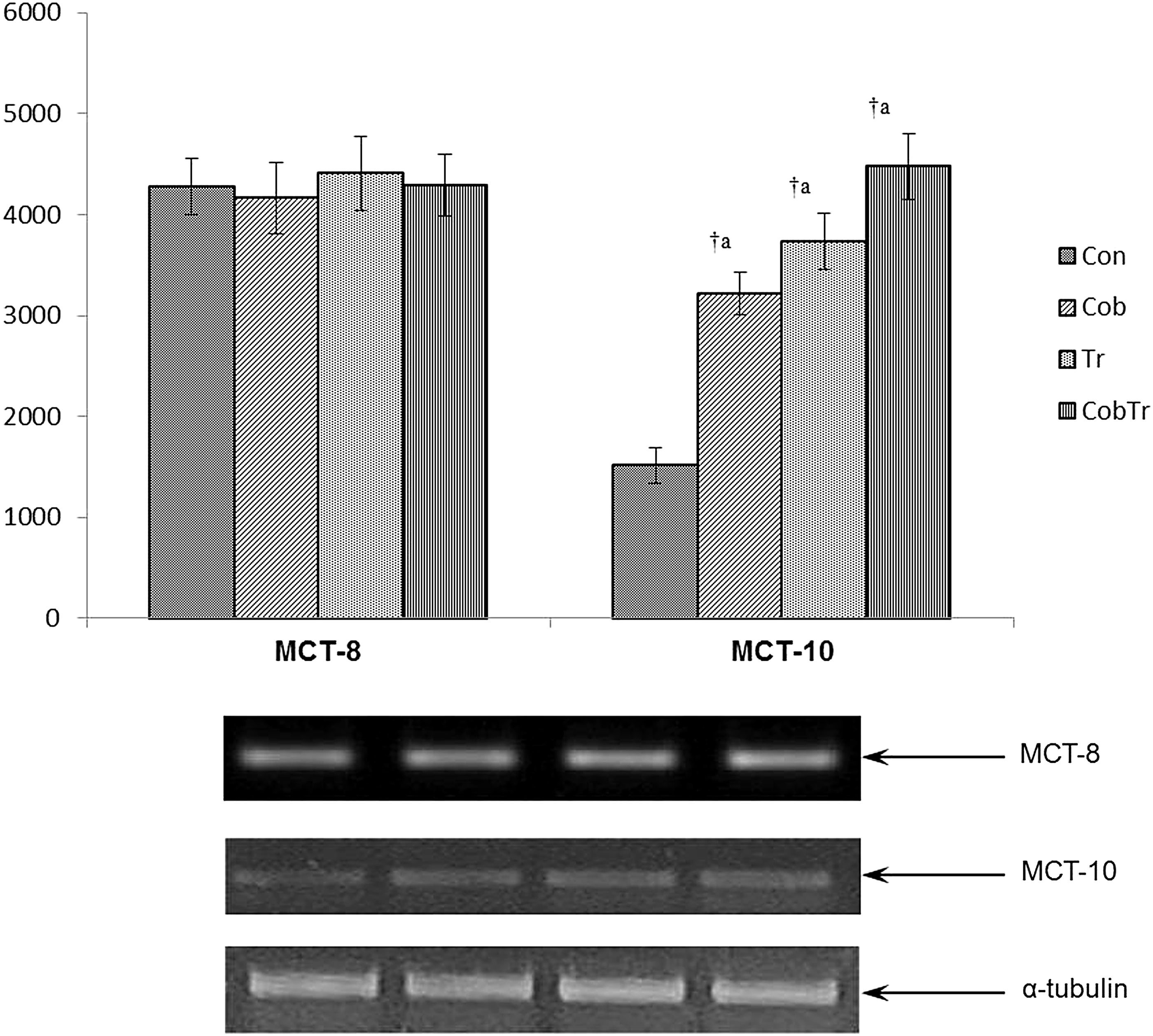
Hypoxia preconditioning and exercise alter the mRNA expression of MCT10, but not MCT8. Data represent the mean ± SD of three independent experiments. Data analyzed by one-way ANOVA with post hoc Bonferroni analysis. aSignificantly different from control. bSignificantly different from training. †Significantly different (p < 0.01).
Protein expression level of MCTs
The protein expression levels of MCT1, 2, and 4 with their quantitative measurement as shown by relative optical density of bands are shown in Figure 4. The protein expression level of MCT1 was enhanced after cobalt supplementation (3.78-fold, p < 0.01) and training (5.09-fold, p < 0.01) compared with control. Cobalt supplementation along with training enhanced the protein expression of MCT1 compared with training only (1.33-fold, p < 0.01). MCT2 protein expression was lowered after cobalt supplementation (0.27-fold, p < 0.01), but increased in response to training (3.1-fold, p < 0.01) relative to control. Cobalt supplementation along with training also reduced MCT2 expression level (0.65-fold, p < 0.01) compared with training only. On the other hand, the protein expression of MCT4 was reduced after cobalt supplementation (0.76-fold, p < 0.01) and training (0.34-fold, p < 0.01) compared with control. Cobalt supplementation along with training reduced (0.42-fold, p < 0.01) the protein expression compared with trained rats.
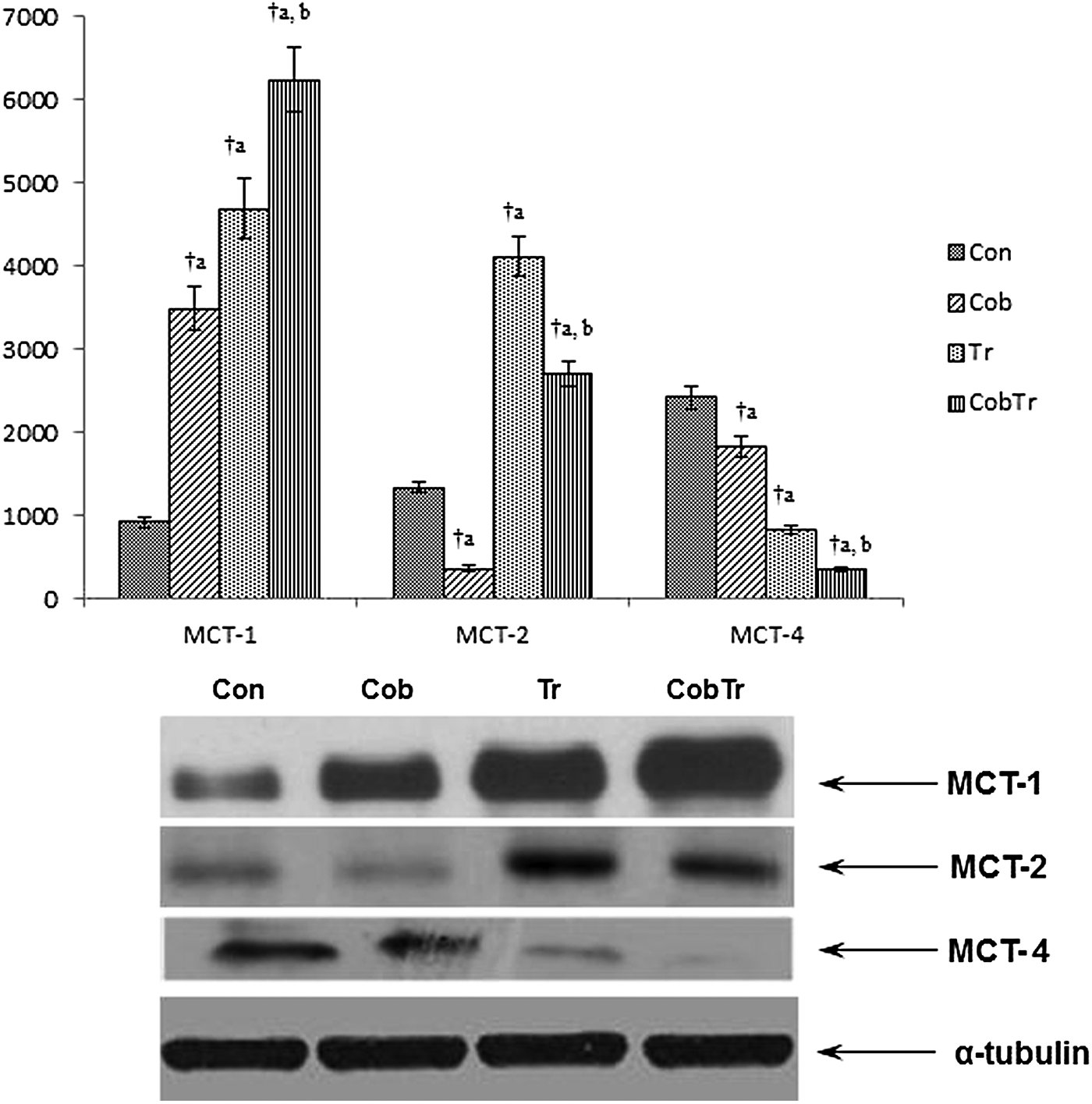
Effect of hypoxia preconditioning and exercise on protein expression levels of MCT1, 2, and 4. Data represent the mean ± SD of three independent experiments. Data analyzed by one-way ANOVA with post hoc Bonferroni analysis. aSignificantly different from control. bSignificantly different from training. †Significantly different (p < 0.01).
Protein expression level of hypoxia and exercise-responsive genes
Figure 5 represents the protein expression levels of HIF1α, PGC1α, and PPARα. The protein expression level of HIF1α increased significantly in hypoxia preconditioned rats (4.90-fold, p < 0.01) compared with control. Training also enhanced (4.73-fold, p < 0.01) HIF1α protein expression relative to control. Cobalt supplementation along with training increased HIF1α expression (2.09-fold, p < 0.01) compared with training. Similarly, the PGC1α protein expression level increased in cobalt-supplemented (2.72-fold, p < 0.01) and trained rats (3.74-fold, p < 0.01) compared with control. Cobalt supplementation along with training enhanced PGC1α expression (1.53-fold, p < 0.01) relative to training only. Moreover, PPARα expression did not alter in the cobalt-supplemented group relative to control, but increased in trained rats (2.77-fold, p < 0.01). The cobalt supplementation along with training group did not show any alteration in PPARα protein expression compared with trained rats.
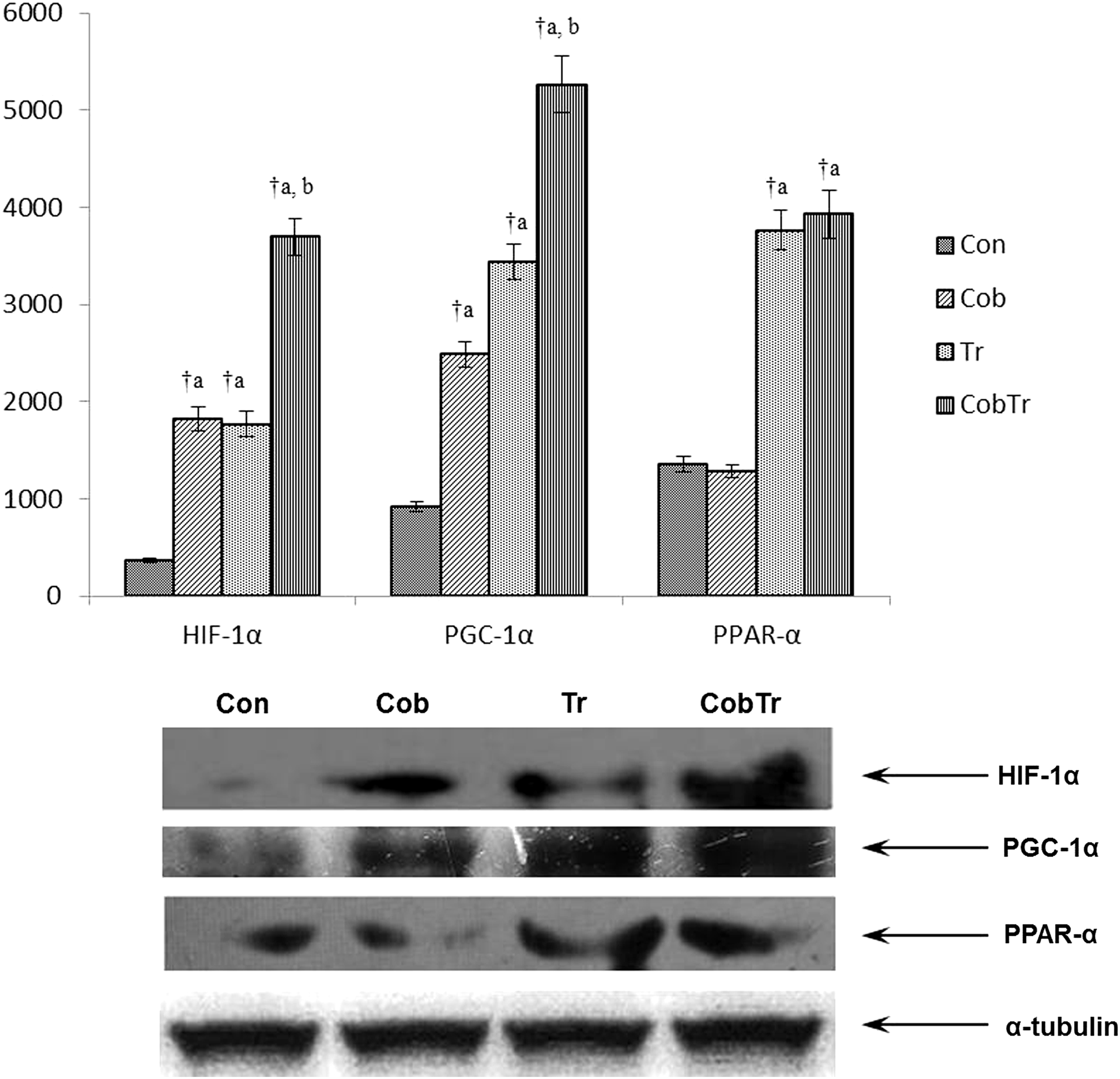
Hypoxia preconditioning and exercise induce the protein expression levels of hypoxia and exercise-responsive genes. Data represent the mean ± SD of three independent experiments. Data analyzed by one-way ANOVA with post hoc Bonferroni analysis. aSignificantly different from control. bSignificantly different from training. †Significantly different (p < 0.01).
Lactate level
A significant reduction in the plasma lactate level was noted after cobalt supplementation (0.78-fold, p < 0.01) and training (0.74-fold, p < 0.01) compared with control. Cobalt supplementation along with training further reduced the plasma lactate level (0.90-fold, p < 0.05) compared with training (Table 2).
Values are mean ± SD (n = 8 per group). Data analyzed by one-way ANOVA with post hoc Bonferroni analysis.
Significantly different from control.
Significantly different from training.
Significantly different (p < 0.01).
Significantly different (p < 0.05).
Food and water intake
There was a significant increase in food intake in the training group compared with control (p < 0.05). Rats of the cobalt supplementation along with training group also took more food compared with control (p < 0.05). Cobalt supplementation only did not have any effect on food intake in rats when compared with controls. Moreover, training with and without cobalt supplementation decreased gain in body weight compared with control (p < 0.05), whereas cobalt supplementation alone did not affect it (Table 3).
Values are mean ± SD (n = 8 per group). Data analyzed by one-way ANOVA with post hoc Bonferroni analysis.
Significantly different from control.
Significantly different (p < 0.05).
Discussion
The major finding of the present study is that most of the isoforms of MCTs are expressed in red gastrocnemius muscle at least at the transcriptional level. In our knowledge, this is the first report to demonstrate the mRNA expression of various MCTs in skeletal muscle and change in their expression levels under hypoxia preconditioning and exercise. We also found that hypoxia preconditioning and endurance training affect the mRNA expression of these genes differently. Lactate is a metabolic intermediate and is utilized as respiratory fuel in tissues such as red skeletal muscle, brain, and heart by converting into pyruvate in mitochondria (Gladden, 2008; Cruz et al., 2012). In the present study, a decrease in the plasma lactate level was noticed, indicating toward an enhanced lactate uptake by skeletal muscle (Table 2). Moreover, we have previously demonstrated that hypoxia preconditioning enhances endurance performance in rats with an increase in skeletal muscle mitochondrial density and aerobic respiratory enzyme activities, indicative of enhanced aerobic respiration by skeletal muscle (Saxena et al., 2012). Moreover, cobalt chloride supplementation did not significantly affect nutrition uptake or change in body weight in rats (Table 3).
Of its 14 isoforms, only MCT1-4 catalyzes proton-coupled transport of lactate (Meredith and Christian, 2008; Halestrap, 2012). MCT1 is ubiquitous in its protein expression (Bonen et al., 2006). In skeletal muscle, it is mainly found on sarcolemmal and mitochondrial membranes (McClelland and Brooks, 2002; McClelland et al., 2003; Butz et al., 2004; Hashimoto et al., 2005). As part of the lactate shuttle mechanism, MCT1 facilitates uptake of lactate from the interstitium and plasma (Bergman et al., 1999; Dubouchaud et al., 2000; Brooks, 2002). The mRNA as well as protein expression of MCT1 increased in cobalt-supplemented and trained rats compared with control. Cobalt supplementation along with training further enhanced the mRNA and protein expression levels of MCT1 compared with trained rats, which is in agreement with our previous study (Saxena et al., 2010). In addition, in our study, an increase in PGC1α protein expression was noted in hypoxia-preconditioned and trained rats. Benton et al. (2008) and Summermatter et al. (2013) have established a strong positive relationship between PGC1α and MCT1. Our study also suggests the same that a high level of PGC1α induces MCT1 overexpression in skeletal muscle.
MCT2 protein has been found to be widely expressed in rodent tissues. It is abundantly expressed in rat heart and skeletal muscle, brain, liver, and kidney (Bonen et al., 2006). In humans, it has been found to be expressed in the same tissues as in rats (Bonen et al., 2006). Although MCT2 has remarkably similar substrate specificity to MCT1, it is shown to be a high-affinity pyruvate transporter in humans (Lin et al., 1998). To date, no significant role of MCT2 has been found in skeletal muscles, but in neurons, it is known to play a critical role in lactate uptake in the brain (Pierre et al., 2000, 2002; Cortes-Campos et al., 2013). In the present study, training induced MCT2 protein as well as mRNA expression in red gastrocnemius muscle of rats, which is in accordance with the study of Takimoto et al. (2013) and Takimoto and Hamada (2014). Moreover, a reduction in MCT2 mRNA and protein expression was found in cobalt-supplemented rats and suggests that MCT2 expression may be partially regulated by HIF.
MCT3 is supposed to be localized exclusively to the basal membrane of the retinal pigment epithelium (Philp et al., 2003) and choroid plexus confined to basal membrane (Halestrap and Wilson, 2012). In addition, some studies confirm its expression in human aorta, kidney, glycolytic skeletal muscle, and vascular smooth muscle cell lines (Zhu et al., 2005; Wang et al., 2006) as well. While MCT3 is not very well (functionally) characterized, it has been reported to play a significant role in facilitating the transport of lactate out of the retina (Halestrap and Wilson, 2012). We found a very low expression of MCT3 mRNA in control rats, which increased significantly by cobalt supplementation and training.
MCT4 possesses great similarities to MCT1 in tissue distribution, regulation, and substrate/inhibitor specificity, but its tissue-specific localization and substrates affinities are different. It is predominantly expressed in highly glycolytic cells such as white muscle and white blood cells, which suggests that its prime physiological function is lactate efflux (Juel and Halestrap, 1999; Manning et al., 2000). In the present study, the protein as well as mRNA expression of MCT4 decreased with cobalt supplementation. Endurance training only also implied the same effect.
A previous report (Ullah et al., 2006) has shown that HIF1 upregulates expression of MCT4, but MCT1 and MCT2 are not regulated by it as both of them do not contain a hypoxia-responsive element in their promoter. Contrary to this, in our study, we have found MCT1 and MCT2 to be affected by HIF1 overexpression; and MCT4 expression was downregulated. Moreover, Pérez de Heredia et al. (2010) have reported MCT4 protein expression under HIF1 overexpression to be unchanged, despite high overexpression at the mRNA level. Therefore, in an attempt to get more insight of the relationship of HIF1 and abovementioned proteins, we also studied the protein–protein interaction of HIF1 with MCT1, 2, and 4. First, we immunoprecipitated HIF1 along with its interacting proteins in solution, and then using the Western blotting technique, we deciphered whether abovementioned proteins interact with HIF1 or not. Our results confirm that there is no direct interaction between HIF1 and these proteins (data not shown).
MCTs, 5–7, 9, and 11–14, are considered together and called as orphan transporters because they are poorly characterized in terms of their substrate and role in physiology (Bonen et al., 2006; Halestrap, 2012). MCT7 protein is mainly expressed in the brain, vas deferens, testes, and liver in rats (Bonen et al., 2006). In humans, it is highly expressed in the brain (Price et al., 1998) and is supposed to play a role in aluminum transport across the blood–brain barrier (Yokel et al., 2002). In the present study, MCT7 mRNA expression was lowered with cobalt supplementation, whereas training did not affect it much. The expression of MCT9 has been found in the endometrium, ovary, breast, brain, testis, kidney, retina, and adrenal gland (Halestrap and Meredith, 2004). The function of MCT9 is not known. Endurance training enhanced MCT9 mRNA expression, whereas cobalt supplementation did not affect its expression. The functions of MCT11 and 12 are not yet known. The expression of MCT11 has been found in the skin, ovary, breast, lung, pancreas, and choroid plexus, whereas MCT12 is found to be present in the kidney (Halestrap and Meredith, 2004). Both of these MCTs had a similar expression pattern. Cobalt supplementation enhanced their expression in skeletal muscle with or without training, whereas training had no effect on it. MCT13 has been found to be expressed in breast and bone marrow stem cells (Halestrap and Meredith, 2004). Its physiological role is still unknown. It has been shown that MCT13 mRNA expression increases after exposure to PPARα agonists in mouse small intestine (Hirai et al., 2007). Perhaps this is the reason why MCT13 expression enhanced in the trained group in our study, whereas it was unaffected in cobalt-supplemented groups.
MCT8 and MCT10 are known to transport thyroid hormones (Friesema et al., 2004, 2008; Schwartz et al., 2005), although MCT10 (also known as T-type amino acid transporter-1; TAT1) is also capable of transporting system T-aromatic amino acids (Friesema et al., 2008). They do not transport monocarboxylate. MCT8 is expressed in a variety of tissues, including the liver, kidney, heart, skeletal muscle, brain, pituitary, and thyroid (Visser et al., 2011). It plays an important role in regulating thyroid hormone uptake in the brain, which is essential for its normal development (Visser et al., 2011). On the other hand, the MCT10 presence has been demonstrated in the kidney, small intestine, skeletal muscle, placenta, and heart (Kim et al., 2001, 2002). We did not find any change in MCT8 expression under exercise or hypoxia preconditioning, whereas mRNA expression of MCT10 increased as a result of both training and cobalt supplementation. Cobalt supplementation along with training could not further modify this increase in trained rats.
Therefore, our study concludes that majority of MCTs are expressed in skeletal muscle at least at the mRNA level. Hypoxia preconditioning and exercise affect the mRNA expression of these MCTs differently, although the mechanism of such an action requires further study.
Footnotes
Acknowledgments
Financial assistance for this work was provided by the Defense Research and Development Organization, Government of India. The authors acknowledge the technical support and encouragement provided by Dr. Rameshwar Singh and Mr. Bhagwat Singh. Saurabh Saxena was a recipient of the Senior Research Fellow (SRF) scholarship from the Council for Scientific and Industrial Research (CSIR), Government of India, and Dhananjay Shukla was a recipient of the Senior Research Fellowship from University Grants Commission (UGC), Government of India.
Author Disclosure Statement
The authors have no conflicts of interest for this study.