
Abstract
Abstract
Goswami, Ananda Raj, Goutam Dutta, and Tusharkanti Ghosh. Naproxen, a nonsteroidal anti-inflammatory drug can affect daily hypobaric hypoxia-induced alterations of monoamine levels in different areas of the brain in male rats. High Alt Med Biol. 17:133–140, 2016.—The oxidative stress (OS)-induced prostaglandin (PG) release, in hypobaric hypoxic (HHc) condition, may be linked with the changes of brain monoamines. The present study intends to explore the changes of monoamines in hypothalamus (H), cerebral cortex (CC), and cerebellum (CB) along with the motor activity in rats after exposing them to simulated hypobaric condition and the role of PGs on the daily hypobaric hypoxia (DHH)-induced alteration of brain monoamines by administering, an inhibitor of PG synthesis, naproxen. The rats were exposed to a decompression chamber at 18,000 ft for 8 hours per day for 6 days after administration of vehicle or naproxen (18 mg/kg body wt.). The monoamine levels (epinephrine, E; norepinephrine, NE; dopamine, DA; and 5-hydroxytryptamine, 5-HT) in CC, CB, and H were assayed by high-performance liquid chromatography (HPLC) with electrochemical detection, and the locomotor behavior was measured by open field test. The NE and DA levels were decreased in CC, CB, and H of the rat brain in HHc condition. The E and 5-HT levels were decreased in CC, but in H and CB, they remained unaltered in HHc condition. These DHH-induced changes of monoamines in brain areas were prevented after administration of naproxen in HHc condition. The locomotor behavior remained unaltered in HHc condition and after administration of naproxen in HHc condition. The DHH-induced changes of monoamines in the brain in HHc condition are probably linked with PGs that may be induced by OS.
Introduction
E
In stressful condition, the changes of behavior and stress hormones may originate from the altered monoamines in the brain (Bhattacharyya and Sur, 1999; Crumeyrolle-Arias et al., 2014; Hu et al., 2014). It has been reported by several investigators that the brain neurotransmitter levels are altered after exposure to HH (Olson et al., 1983; Saligaut et al., 1986; Goroshinskaia and Neskubina, 1998). The norepinephrine (NE) and dopamine (DA) concentrations in the rat whole brain during chronic exposure (7 days at 380 Torr) to HH were reduced to about 40% of normoxia control. The 5-hydroxytryptamine (5-HT) level in the whole brain also decreased to 50%–60% of normoxia control during acute hypoxia (1 day at 420 Torr) (Olson et al., 1983). The NE level was decreased in hypothalamus (H) after exposure to HH compared to normoxia (Saligaut et al., 1986). Goroshinskaia and Neskubina (1998) reported that the monoamine level in H was increased during the initial exposure to hypoxia (3 hours at 9000 m) but was decreased after exposure to intermittent hypoxia (4 hours daily for 3–10 days at 5000 m) in rats. These studies suggest that the HH-induced changes of monoamine levels in the brain may be dependent on the degree and duration of exposure, and hypothalamic monoamines, being the regulator of hypothalamic releasing hormones, are likely to be altered in HHc condition. Similarly, monoamines in the cerebral cortex (CC) and cerebellum (CB) may be linked with the HH-induced cognitive and motor changes.
The brain is highly sensitive to oxygen in HHc condition in rats (Szaflarski et al., 1995). It has been reported that prostaglandin E (PGE) secretion in the brain was increased in HH probably by the production of reactive oxygen species (ROS) (Cao et al., 2012; Park et al., 2012). Several reports have suggested that the PGE might inhibit the release of NE, E, DA, and 5-HT (Starke, 1977; Reimann et al., 1981; Schlicker et al., 1984) in different brain areas, such as CC, striatum, H, and so forth, in stressful conditions. Naproxen, an inhibitor of cyclooxygenase (COX) enzymes and 20 times more potent than aspirin, was also effective in inhibiting the prostaglandin (PG) production in HHc condition (Biswas et al., 1996). Many nonsteroidal anti-inflammatory drugs, such as aspirin, ibuprofen, and naproxen, were used by mountaineers at high altitude to facilitate acclimatization and avoid high-altitude headache (Burtscher, 1999; Burtscher et al., 2001). It has been suggested that PGs may be important mediators between hypoxia and high-altitude headache (Burtscher, 1999). Our earlier study has shown that naproxen has important role in preventing the HH-induced immune changes, which are mediated by corticosterone (CORT) and/or production of PGs in HHc condition (Goswami et al., 2012). The effect of naproxen on the HH-induced immune changes may be related to neurotransmitter levels in the brain, especially in H. However, no studies so far have dealt with the effects of naproxen on HH-induced alteration of brain monoamines.
Therefore, the present study intends to explore the changes of monoamines in H, CC, and CB along with the motor activity in rats after exposing them to simulated hypobaric condition daily that induces different immunological changes in previous experiments and the role of PGs on the daily hypobaric hypoxia (DHH)-induced alteration of brain monoamines, if any, by administering naproxen.
Materials and Methods
Animals
Adult (6–8 weeks of age) male albino rats (Charles-Foster strain) weighing 200–220 gm were used in this study. Animals were housed individually in polypropylene animal cages with food pellet and water ad libitum. The animal room was maintained at a temperature of 25°C ± 1°C with a 12-hour light–12-hour dark cycle (light 7 am to 7 pm). According to the Institutional Animal Ethics Committee, all adequate measures were taken to minimize the pain and discomfort to the rats.
Design of experiments
The brain monoamine [NE, epinephrine (E), DA, and 5-HT] levels in different brain areas, such as CC, CB, and H, and locomotor behavior by open field study were measured in rats exposed either to HHc condition or to control at normobaric condition (CN). There are two groups of rats in normobaric condition (CN and CNP) and two groups of rats in HHc condition (HH and HHNP). Two groups of rats in normobaric condition were CN (vehicle only, without naproxen, or 0 mg/kg body wt. of naproxen) and CNP (18 mg of naproxen/kg body wt.). In HHc condition, two groups of rats include HH (without naproxen and vehicle only) and HHNP (18 mg of naproxen/kg body wt.). The concentrations of brain monoamines, such as NE, E, DA, and serotonin (5-HT), were measured in three different areas of the rat brain, such as CC, CB, and H, in all the four groups. The animals were sacrificed immediately after completion of the last day of exposure to DHH at the same hour of the day (5.00–6.00 pm). Locomotor behavior of all the rats of different groups was measured for 3 consecutive days before the exposure to DHH and during 5 days of exposure to HH.
Exposure to HH
The rats were exposed to simulated DHH at 18,000 ft (380 Torr) in an HHc chamber 8 hours daily for 6 days from 9.00 am to 5.00 pm following the method used by several groups of investigators (Purshottam and Ghosh, 1973; Biswas et al., 1996; Chen et al., 2007; Fan et al., 2009; Ghosh, 2009; Baitharu et al., 2011; Goswami et al., 2012). Briefly, the decompression chamber consists of a metallic body and is connected to a vacuum pump and a mercury manometer. The atmospheric air can circulate through the chamber during the operation of the vacuum pump, while the pressure within the decompression chamber can be monitored continuously. The pressure within the chamber may be adjusted by regulating the flow of air through the chamber. The temperature inside the chamber was maintained at 25°C ± 1°C, and the chamber remained lighted during exposure to the animals. Food and water were available to the animals during exposure. The normobaric rats were placed in a similar chamber but were not subjected to HHc condition.
Treatment of naproxen
Naproxen (RPG Life Sciences Ltd.) was dissolved in 10% alcohol before experiment, and it was administered orally (18 mg/kg body wt.) through a gastric cannula attached to a 1-mL syringe. The daily dose of naproxen was divided equally into two parts: one part (half of the total dose, i.e., 9 mg of naproxen/kg body wt.) was given orally 5–6 minutes before the exposure to DHH, and the other part (rest half of the total dose, i.e., 9 mg of naproxen/kg body wt.) was given immediately after the exposure to DHH on each day. This dose of naproxen (18 mg/kg body wt.) was selected on the basis of optimum effect of naproxen on hematological and immunological parameters in a previous study with different doses of naproxen (Goswami et al., 2012).
Locomotor behavior
Rats were tested for locomotor behavior by open field test as described by Vogel and Huger (2002) and Ray et al. (2003). The open field test apparatus (1 × 1 m) was made up of plywood, surrounded by a wall 40 cm high, with the inner surface painted black. The surface of the floor was divided into 25 squares of the same size. The rats were placed gently in the center of the open field test apparatus, where the rats were free to walk and to get adjusted to the new environment and for training from 5.00 pm to 7.00 pm for 3 consecutive days before the exposure to DHH was started. After the training, the exposure schedule (exposure to DHH for 8 hours/day for 6 days) was started, and the locomotor behavior was studied on each day in all the groups (CN, CNP, HH, and HHNP) from 5.00 pm to 7.00 pm. The number of squares crossed in 5 minutes was counted at an interval of 30 minutes for a total duration of 2 hours (5.00–7.00 pm). Thus, there were five episodes of 5-minute study for 2 hours every day.
Brain monoamine assay by HPLC method
The concentrations of brain monoamines, such as NE, E, DA, and serotonin (5-HT), were measured in three different regions of the rat brain, such as CC, CB, and H, according to the method of Aydin et al. (2008). After completion of the exposure schedules, the rats were sacrificed by decapitation; the brains were rapidly removed, and the different brain areas, such as CC, CB, and H, were dissected out of the brain on a plate (covered with aluminum foil) chilled to 0°C. Each portion of brain areas was weighed and kept at −20°C before further processing. Tissues from different brain areas were homogenized in 300 μL dihydroxybenzylamine (2 ng/20 μL) plus 600 μL ice-cold 0.1 M HCl cocktail solution by the disposable polypropylene microtube with pestle, followed by centrifugation at 3000 rpm for 30 minutes at 4°C. The clear supernatant was collected and filtrated by syringe filter through a 0.2-μm filter paper (Rivera). These freshly prepared samples were immediately analyzed by reverse phase high-performance liquid chromatography (HPLC) (Waters) with the help of HPLC analytical C18 column (Waters Spherisorb, 4.6 × 150 mm, 5 μm) (Waters) using an electrochemical detector (Model 2465; Waters). The mobile phase consisted of citric acid (Merck), sodium citrate (Merck), ethylenediaminetetraacetic acid (EDTA; Merck), heptanesulfonic acid (Merck), glacial acetic acid (Merck), tetrahydrofuran (THF) (Merck), and HPLC grade methanol (Merck). Volume was made up to 1 L with HPLC grade water and and then passed through a 0.2-μm filter paper (Rivera) and degassed under a vacuum and pH adjusted to 4.9. In all instances (standard and sample analysis), the mobile phase flow rate was set at 1 mL/min, with the potential set at 1200 PSI. A 20 μL sample was injected via HPLC pump (Waters 515 HPLC Pump) for detection. The standard mixtures were prepared daily, and different concentrations of aqueous solutions of NE, E, DA, and 5-HT (Sigma) were injected into the column. The peak of each neurotransmitter of each sample was quantified from their respective peak heights obtained from the reference standards by an electrochemical detector. Concentration of the neurotransmitters in the sample was evaluated from reference standards using the Empower software (Empower Chromatography Software; Waters).
Statistical analysis
Data are expressed as mean ± standard error of the mean (SEM). One-way ANOVA was used to compare the data of different groups in normobaric condition and HHc condition, followed by LSD post hoc test using the Statistical Package for Social Science software (SPSS Software 20.0.0). A p value ≤0.05 was considered as a significant difference.
Results
Locomotor behavior
The locomotor activity of rats was measured in open field test in rats both in CN and in HHc condition. The locomotor activity was also measured after administration of naproxen at a dose of 18 mg/kg body wt. in CN condition (CNP) and HHc condition (HHNP). There was no significant change in the locomotor activity of rats after administration of naproxen in CNP, HH, and HHNP groups compared to CN (Table 1).
Values are expressed as mean ± standard error of the mean (SEM) (n = 6 in each groups). There was no significant difference among the different groups.
CN (vehicle only, without naproxen or 0 mg/kg body wt. of naproxen), CNP (18 mg of naproxen/kg body wt.), HHNP (without naproxen, vehicle only) and HHNP (18 mg of naproxen/kg body wt.).
Brain monoamine assay
NE level
NE concentration was decreased in CC [F (3, 20) = 11.589, p < 0.001], CB [F (3, 20) = 7.721, p < 0.001], and H [F (3, 20) = 9.756, p < 0.01] in HH compared to CN. Naproxen did not produce any significant change in all the three brain areas in CNP compared to CN. However, prior treatment with naproxen significantly prevented the HH-induced reduction of NE in CC, CB, and H of the rat brain in HHNP compared to CN and CNP (Fig. 1).
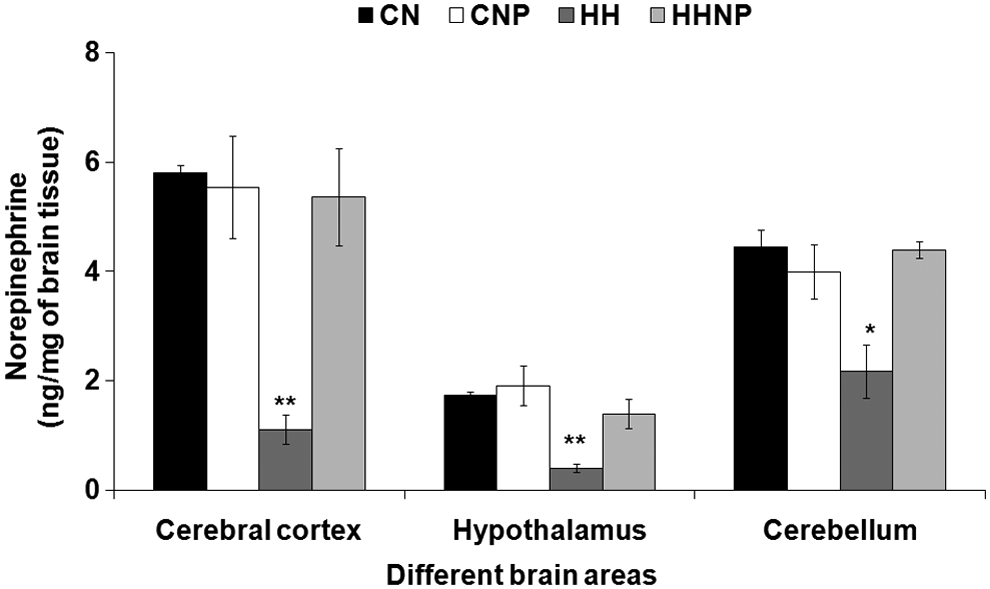
The norepinephrine (NE) level in brain areas [cerebral cortex (CC), hypothalamus (H), and cerebellum (CB)] in CN (control at normobaric condition), CNP (administration of 18 mg of naproxen/kg body wt. in control at normobaric condition), HH (exposure to HH), and HHNP (administration of 18 mg/kg body wt. of naproxen at HHc condition) groups of rats. The NE level was decreased significantly in CC, H, and CB areas of rats' brain in HH group of rats compared to CN and CNP at *p < 0.01 and **p < 0.001. Values are expressed as mean ± SEM (n = 6 in each group). HH, hypobaric hypoxia; HHc, hypobaric hypoxic; SEM, standard error of the mean.
E level
E concentration was decreased in CC [F (3, 20) = 38.767, p < 0.001] but not in CB and H in HH compared to CN. Naproxen was administered at a dose of 18 mg/kg body wt. in CNP and was unable to produce any significant change in all the three brain areas compared to CN. Naproxen protected the HH-induced reduction of E level in CC, but the E level in CB and H remained unaltered (Fig. 2).
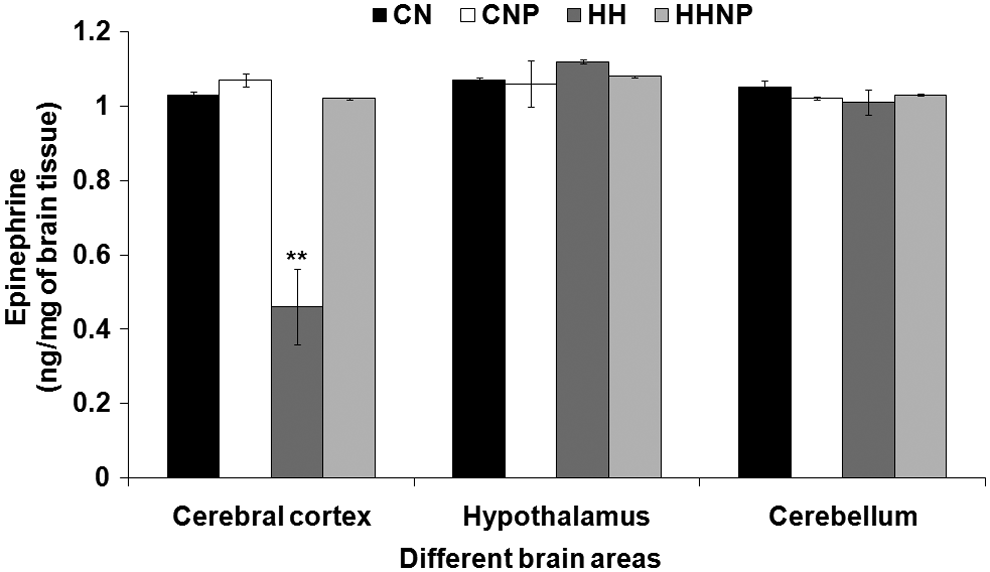
The epinephrine (E) level in brain areas (CC, H, and CB) in CN, CNP (administration of 18 mg of naproxen/kg body wt. in control at normobaric condition), HH (exposure to HH), and HHNP (administration of 18 mg/kg body wt. of naproxen at HHc condition) groups of rats. The E level was decreased significantly only in CC areas of rats' brain in HH group of rats compared to CN and CNP at **p < 0.001. Values are expressed as mean ± SEM (n = 6 in each group).
DA level
Exposure to DHH had markedly increased the concentration of DA in CC [F (3, 20) = 5.098, p < 0.01], CB [F (3, 20) = 8.161, p < 0.05], and H [F (3, 20) = 38.573, p < 0.001] in HH compared to CN. Naproxen significantly altered the DA level in all the three brain areas in CNP compared to CN. Treatment with naproxen significantly recovers the HH-induced reduction of DA level in the CC, CB, and H in HHNP compared to CN and CNP (Fig. 3).
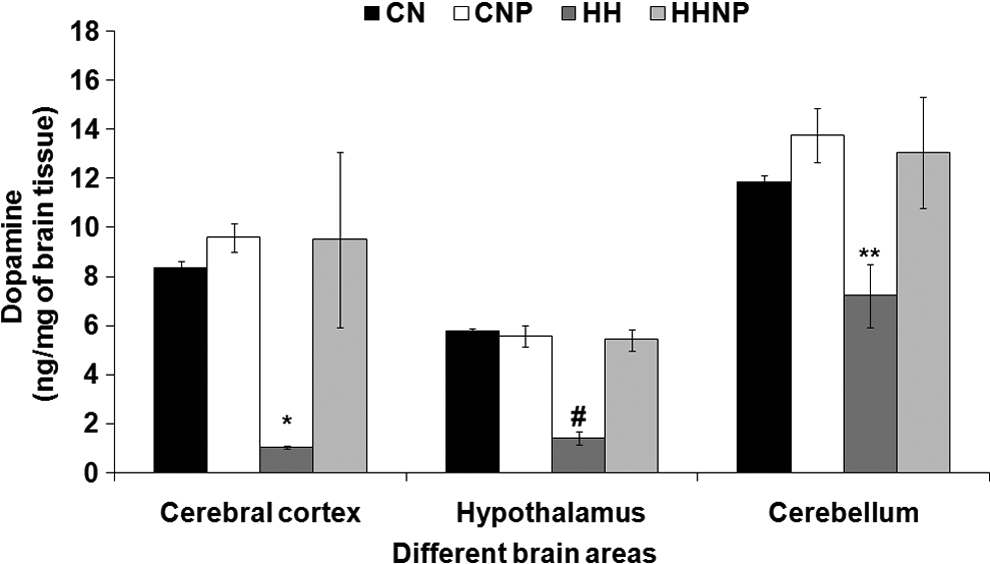
The dopamine (DA) level in brain areas (CC, H, and CB) in CN, CNP (administration of 18 mg/kg body wt. of naproxen in control at normobaric condition), HH (exposure to HH), and HHNP (administration of 18 mg of naproxen/kg body wt. at HHc condition) groups of rats. The DA level was decreased significantly in CC, H, and CB areas of rats' brain in HH group of rats compared to CN and CNP at *p < 0.01,**p < 0.001, and #p < 0.05. Values are expressed as mean ± SEM (n = 6 in each group).
5-HT level
The 5-HT level was reduced only in CC [F (3, 20) = 10.515, p < 0.001], but it remained unchanged in CB and H in HH compared to CN. Naproxen treatment showed no significant change in all the three brain areas in CNP compared to CN. Treatment with naproxen had recovered the HH-induced reduction of the 5-HT level in CC, while the 5-HT level in CB and H remained unaltered after treatment with naproxen in HHNP compared to CNP and CN (Fig. 4).
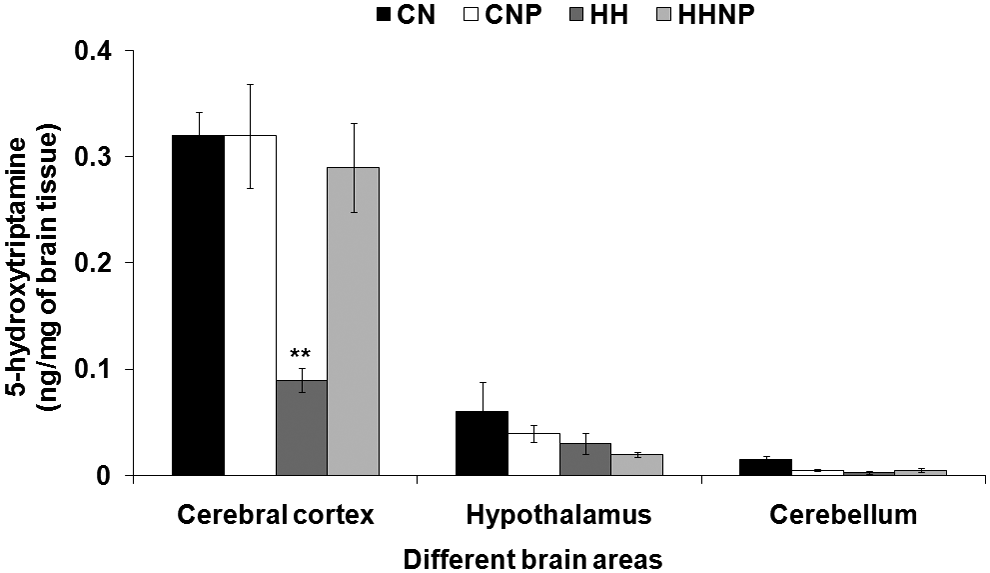
The 5-hydroxytryptamine (5-HT) level in brain areas (CC, H, and CB) in CN, CNP (administration of 18 mg of naproxen/kg body wt. in control at normobaric condition), HH (exposure to HH), and HHNP (administration of 18 mg/kg body wt. of naproxen at HHc condition) groups of rats. The 5-HT level was decreased significantly only in CC areas of rats' brain in HH group of rats compared to CN and CNP at **p < 0.001. Values are expressed as mean ± SEM (n = 6 in each group).
Discussion
It has been widely reported that the different stressors are responsible for changes of the brain functions through the alteration of neurotransmitters in the brain (Crumeyrolle-Arias et al., 2014; Hu et al., 2014). The cognitive and motor functions are affected in HHc condition (Hamilton et al., 1991; Bahrke and Shukitt-Hale, 1993; Shukitt-Hale and Lieberman, 1996). The motor activities are controlled by the CC, CB, and other areas, and the cognitive functions are also linked with cerebral cortical activity. Probably neurons of specific areas of the brain controlling these functions are affected in DHH. The NE and DA levels were decreased in CC and CB in HHc condition in the present study. A similar decrease of these two brain monoamines was reported earlier in the whole rat brain after exposure to HH for 6, 12, and 18 hours compared to the normobaric rats (Cymerman et al., 1972). The present study also demonstrated that the present exposure protocol to HH resulted in significantly reduced E and 5-HT levels only in CC of the rat brain, and these neurotransmitters remained unaltered in CB. Olson et al. (1983) found a similar decrease of 5-HT in the whole brain and brain stem after the rats were exposed to HH from 1 hour to 7 days at 450 Torr. However, the change of 5-HT in that study was of transient nature. Olson et al. (1983) also reported a transient change of NE and DA in that experimental condition. There was no report on the E level in brain areas during HH. In the present exposure protocol of HH, the levels of all the monoamines (E, NE, DA, and 5-HT) were reduced in CC, and NE and DA were decreased in CB. These changes of monoamines in CC and CB may have influence on the motor activity. The impairments of the motor functions in rats have been correlated with the alteration of brain monoamines in different brain regions (Päivärinta et al., 1991; Golembiowska et al., 2012; Khaliq et al., 2012; Qiu et al., 2013). The altered motor behavior in rats infected with herpes simplex virus was found to be associated with the changes of brain DA and 5-HT metabolism in major ascending monoamine system (Päivärinta et al., 1991). However, the observed locomotor activity in the present study remained unaltered in HHc condition. Vataeva et al. (2004) also reported that the locomotor activity (horizontal activity and the number of squares crossed) was not affected after exposure to HH in male rats. The DHH-induced reduction of NE, E, DA, and 5-HT levels was significantly attenuated by the administration of naproxen in HHc condition. However, the locomotor activity remained unaltered also after administration of naproxen in both HHc condition and normobaric condition. Probably the DHH-induced alterations of monoamines in CC and CB were not sufficient to affect the gross motor activity as measured in the open field of the present study. The finer motor movement may be sensitive to the subtle changes of the activity of motor regulatory neural circuitry but probably the gross movement, such as locomotor activity, is not affected in that condition. From the present study, it appears that the locomotor activity was also less likely to be affected by naproxen in DHH.
The NE and DA levels were decreased in H in HHc condition of the present study. The reduction of the hypothalamic NE and DA levels in HHc condition was also reported by others (Saligaut et al., 1986; Goroshinskaia et al., 1993; Goroshinskaia and Neskubina, 1998). However, Goroshinskaia et al. (1993) reported that the NE and DA levels were increased in H after exposure to acute hypoxia (9000 m above the sea level for 3 hours) and considerably decreased after exposure to intermittent hypoxia (at 5000 m 4 hours daily for 3 and 10 days). Considering the protocol of HH exposure in the present investigation, the changes of NE and DA in H appear to be consistent with the results of previous investigations. The present investigation also revealed that the E and 5-HT levels remained unaltered in H. There was no such report on the changes of the E and 5-HT levels in H area of the brain in HHc condition. In different stressful conditions, the regulation of hypothalamic–pituitary–adrenal axis is mediated through the different neurotransmitter systems (Van de Kar, 1991). In systemic physiological stress, such as hypoxia, brain stem catecholaminergic inputs on corticotropin-releasing hormone (CRH) neurons of H are stimulatory (Kronenberg et al., 2008). The 5-HT axonal projections from raphe nucleus also have a stimulatory effect on CRH neurons of H directly or through stimulating the hypothalamic glutamatergic inputs. The concentration of serum CORT was increased in HHc condition following a similar protocol of exposure in simulated chamber (Goswami et al., 2012, 2014). The lower hypothalamic levels of NE, DA, and unaltered E, 5-HT cannot explain the elevated level of CORT in HHc condition of the present study. However, other neurotransmitters, such as glutamate, neuropeptide Y, and serum level of CORT, cytokines, and PGs may also have modulatory effects in HHc condition, and these were not measured in the present study.
The mechanism of changes of the observed brain monoamines in different areas of the brain in HHc condition has not been explored sufficiently in this study. However, prevention of DHH-induced changes of brain monoamine by the administration of naproxen probably indicates that the COX-mediated steps are involved in the process. In the brain, inducible COX-2 is present along with COX-1, which is constitutively expressed (Yasojima et al., 1999; Zarghi and Arfaei, 2011). Naproxen can block both these COX (Dhir et al., 2006; www.patient.info/health/painkillers May 1, 2015). Hypoxia is known to modulate inflammatory gene expression in different tissues, including microglia (Smith et al., 2013). Although PGs and ROS in the brain were not measured in the present study, other investigators showed that brain monoamines might be changed by ROS (Kavitha et al., 2011). The effect of naproxen on DHH-induced changes of brain monoamines may be explained on the basis of its inhibitory action on the HH-mediated increased COX activity and PG secretion. It has been reported that the increased PG level in the brain can reduce the monoamine levels (Yazaki, 1989). The changes of brain monoamines in HHc condition were not associated with changes in locomotor behavior, but other possible physiological effects were not investigated in the present study. However, some immune responses were reported to be changed in rats exposed to a similar protocol of HH (Goswami et al., 2012, 2014) and to other experimental schedule of hypobaric exposure (SaiRam et al., 1998). Neveu et al. (1994) reported that the changes of brain monoamines in stressful conditions had been associated with the immune responses. Several workers also reported that stress-induced changes in sleep and anxiety were linked with brain monoamines (Hale et al., 1984; Filip et al., 2005; Jayanthi and Ramamoorthy, 2005). Therefore, it appears that some of the HH-induced physiological changes may be related to the brain monoamine levels in that condition.
Conclusion
The DHH-induced changes of the brain monoamine levels observed in the present study are recovered by naproxen, which is a blocker of PGs. The study indicates that production of PGs in HHc condition may be involved in the changes of DHH-induced brain monoamines in rats.
Footnotes
Acknowledgments
The research has been supported by the fund from the DIPAS, Delhi, DRDO, Government of India (Task No. TC/290/TASK-114(TKG)/DIPAS/2006). The authors also extend their thanks to the Head of the Department of Human Physiology with Community Health, Vidyasagar University, West Bengal, India, for allowing some laboratory facilities of that department and Mr. Nilotpal Mandal, Research Scholar, Department of Human Physiology with Community Health, Vidyasagar University, West Bengal, India, for his technical assistance.
Author Disclosure Statement
No competing financial interests exist.