
Abstract
Abstract
Padhy, Gayatri, Anamika Gangwar, Manish Sharma, Kalpana Bhargava, and Niroj Kumar Sethy. Plasma proteomics of Ladakhi natives reveal functional regulation between renin–angiotensin system and eNOS–cGMP pathway. High Alt Med Biol. 18:27–36, 2017.—Humans have been living in high altitudes for more than 25,000 years but the molecular pathways promoting survival and performance in these extreme environments are not well elucidated. In an attempt to understand human adaptation to high altitudes, we used two-dimensional gel electrophoresis combined with MALDI-TOF/TOF to identify plasma proteins and associated pathways of ethnic Ladakhi natives residing at 3520 m. This resulted in the identification of 36 differential proteins compared with sea-level individuals. Proteins belonging to coagulation cascade and complement activation were found to be less abundant in Ladakhi natives. Interestingly, we observed lower abundance of angiotensinogen (ANGT) and subsequent analysis also revealed lower levels of both ANGT and angiotensin II (Ang II) in Ladakhi natives. Concomitantly, we observed elevated levels of eNOS, phosphorylated eNOS (Ser1177), and plasma biomarkers for nitric oxide (NO) production (nitrate and nitrite) and availability (cGMP). These results suggest that functional interplay between renin–angiotensin system and NO–cGMP pathway contributes to the hypoxia adaptation in Ladakhi natives. These findings will augment the present understanding of higher NO and NO-derived metabolite availability during human adaptation to high altitude.
Introduction
T
With respect to ethnic populations living in the Himalayas, many physiological and genetics studies have been performed on residents of the Tibetan Plateau. Multiple genomewide scans on Tibetans have independently identified members of hypoxia-inducible factor (HIF) pathways namely HIF2A (EPAS1) gene and the prolyl hydroxylase domain protein 2 (also known as EGLN1) gene for high-altitude adaptation (Beall et al., 2010; Bigham et al., 2010; Simonson et al., 2010; Yi et al., 2010; Peng et al., 2011; Xu et al., 2011). Unfortunately, very limited omics-based studies have been carried out on populations residing on the southern side of the Himalayas, that is, in the alluvial plains of the Indian subcontinent. Studying genetic contribution of the endothelial nitric oxide synthase (eNOS) gene polymorphisms in Sherpas, Droma et al. (2006) have reported that the Glu298Asp and eNOS4b/a polymorphisms of the eNOS gene may provide advantages for high-altitude adaptation. In corroboration, Ahsan et al. (2005) have also reported that eNOS wild-type homozygotes (GG, BB) were more prevalent in Ladakhi monks contributing to higher NO levels associated with high-altitude adaptation. Similarly independent studies have also reported that the I allele (Droma et al., 2008) or II genotype (Qadar Pasha et al., 2001) of ACE gene is also responsible for high-altitude adaptation. Furthermore, association of ACE I/D and AGT M235T polymorphisms in the renin–angiotensin system (RAS) genes has been reported as a risk factor for high-altitude pulmonary edema (HAPE) (Srivastava et al., 2012).
In this study, we aimed to profile the plasma proteome of Ladakhi natives to identify proteins and associated pathways facilitating human adaptation to high altitude. Using two-dimensional gel electrophoresis (2-DGE) combined with MALDI-TOF/TOF, we could successfully identify 36 differential plasma proteins in Ladakhi natives as compared with sea-level individuals. Pathway analysis revealed lower levels of complement and coagulation proteins in high-altitude natives, which are subsequently validated by immunoblotting. We also identified lower abundance of angiotensinogen (ANGT) in Ladakhi natives and further validated the expression levels of ANGT and angiotensin II (Ang II), the key members of renin–angiotensin pathway. In addition, we have also estimated the levels of eNOS, phosphorylated eNOS (Ser1177), plasma nitrate, and nitrite as well as cGMP levels both in Ladakhi natives and in sea-level individuals.
Materials and Methods
Study group and blood collection
Ten healthy, nonsmoking, high-altitude male natives (age: 19–27 years, height: 165 ± 2 cm, weight: 56.4 ± 1 kg) permanently residing at Leh, Ladakh (3520 m) for generations participated in this study (HAN group). These subjects had not visited altitudes below 2500 m in the last 1 year. Equal number of healthy normobaric male volunteers (age: 19–27 years, height: 170 ± 2 cm, weight: 60 ± 1 kg) permanently residing at Delhi (716 m) were included in the study as control (NC group). Subjects receiving antibiotic treatment, with acute or chronic clinical conditions, and high-altitude travel (above 2500 m) in last 6 months were excluded from this study. All the subjects were allowed to eat their normal diet and restrained from any heavy physical exercise 7 days before collection of samples. The study protocol was approved by the Institute's ethical committee and written consent was obtained from all the participants after providing information about the experimental procedures involved in the study. Overnight fasting venous blood samples from antecubital vein were collected in EDTA vacutainers in the morning (between 08:00 hour and 09:00 hour) at Delhi and Leh, respectively. Plasma was separated from blood on centrifugation at 1000 g for 15 minutes at 4°C and was stored with protease and phosphatase inhibitor for further use.
Sample preparation for 2-DGE
The blood plasma samples were depleted for high abundant proteins (albumin, prealbumin, transferrin, fibrinogen, IgG, IgA, IgM, α2 microglobulin, haptoglobin, Apo A1, Apo B, lipoprotein A, factor H, and ceruloplasmin) using ProteoMiner protein enrichment kit (Bio-Rad laboratories, CA) according to the manufacturer's instructions. Depleted plasma (100 μL) was precipitated using 10 volumes of 10% trichloroacetic acid (TCA) in acetone on incubating at −20°C overnight. After centrifugation at 10,000 g for 10 minutes at 4°C, the protein pellet was washed with ice-cold acetone and was dissolved in lysis buffer (8 M urea, 2 M thiourea, 40 mM Tris–HCl, 4% (w/v) 3-[(3-Cholamidopropyl)dimethylammonio]-1-propanesulfonate [CHAPS], and 0.5% Igepal CA-630). The protein concentrations were determined using Bradford reagent (Sigma, St. Louis, MO).
2-DGE and silver staining
For the first dimension isoelectric focusing, IPG strips (Immobiline Dry Strip, pH 4–7, 18 cm; GE Healthcare, PA) were rehydrated overnight with 500 μg of precipitated protein in 350 μL of rehydration buffer [7 M urea, 2 M thiourea, 1.2% (w/v) CHAPS, 20 mM DTT, 0.4% (w/v) ASB-14, 0.25% (v/v) pH 4–7 ampholytes, and 0.005% (w/v) bromophenol blue]. After passive rehydration, isoelectric focusing was carried out for a total of 60,000 Vh (50 μA/strip) at 20°C using IPGphor system (Amersham Pharmacia Biotech, NJ). The focused IPG strips were equilibrated and reduced first with equilibration buffer [6 M urea, 30% v/v glycerol, 2% (w/v) SDS, and 50 mM Tris pH 8.8] containing 1% (w/v) DTT for 15 minutes and then alkylated with equilibration buffer containing 2.5% (w/v) iodoacetamide for 15 minutes. The second dimension separation was carried out on 12% SDS PAGE using EttanDalt Six Electrophoresis system (GE Healthcare, PA) with constant current of 25 mA per gel at 25°C for 6 hours. The proteins were observed with silver staining.
Image acquisition and gel analysis
The silver-stained gel images were digitalized on Investigator™ ProPic II (Genomic Solutions, United Kingdom) and were analyzed on Progenesis Same Spots software (version 4.0; Nonlinear Dynamics, United Kingdom). The normalized spot volume was used for spot calculation and comparison between the two groups. The statistical significant spots (p < 0.05) between the high-altitude natives and sea-level groups were identified and excised for mass spectroscopy (MS).
In-gel tryptic digestion and dried droplet spotting
The excised silver-stained gel protein spots were washed with milliQ water twice for 10 minutes each and were subjected to in-gel trypsin digestion. In brief, the gel pieces were treated with Farmer's reagent (30 mM potassium ferricyanide and 100 mM sodium thiosulfate) (1:1), washed thrice with the milliQ water, and subsequently incubated with 10 mM ammonium bicarbonate and acetonitrile (1:1) for 5 minutes at room temperature. The gel pieces were dehydrated with acetonitrile and were dried by Speed Vac (Heto MAXI dry plus, United Kingdom). Freshly prepared trypsin (20 ng/μL) (Sequencing grade; Promega, Madison, WI) was added to the dried gel spots and incubated for 30 minutes at 4°C and further overnight at 37°C. The tryptic-digested peptides were extracted by sonicating with 50% acetonitrile and 0.1% TFA and were dried in Speed Vac.
Mass spectroscopy analysis and database search
The tryptic-digested peptides were resuspended in 5 μL of 50% v/v acetonitrile: 0.1% v/v TFA. The resolubilized peptides were mixed with saturated solution of cyano-4-hydroxy-cinnamic acid matrix (1:1) (Bruker Daltonics, Germany), were spotted onto the MALDI target plate, and allowed to air dry. The peptide mass spectrum was acquired by MALDI TOF/TOF (Ultraflex III; Bruker Daltonics, Bremen, Germany) in the reflectron ion mode at the accelerating voltage of 25 kV using the Flex Control software (Version 3.0; Bruker Daltonics). The instrument was calibrated with mass standards: bradykinin (m/z 757.39), Ang II (m/z 1046.54), Ang I (m/z 1296.68), substance P (m/z 1347.73), bombesin (m/z 1619.82), ACTH fragment 1–17 (m/z 2093.08), ACTH fragment 18–39 (m/z 2465.39), and somatostatin (m/z 3147.47). For peptide mass fingerprinting, the monoisotopic peak list was generated in postprocessing and true peptide mass list was exported to Biotools (Version 3.1; Bruker Daltonics) using Flex Analysis 3.0 without using smoothing function and submitted to MASCOT database search engine (Matrix science, Boston, MA). The SWISS-PROT database was used with the following search parameters: Homo sapiens as taxonomy with ±100 ppm peptide mass tolerance, one maximum missed cleavage site permitted and carbamidomethyl (cysteine) as fixed modification. Proteins that returned MOWSE (MOlecular Weight SEarch) scores over a threshold score with p < 0.05 were considered significant (Pappin et al., 1993). For each identified protein, at least one peptide was selected for confirmation by MS/MS analysis. The MS/MS spectra were acquired in LIFT mode using the same spot on the target and fragment mass tolerance of ±0.8 Da was applied.
Bioinformatic analysis
The pathway was generated through Kyoto Enclyclopedia of Genes and Genomes pathway. The functional enrichment associated with altitude adaptation based on ontologies of differentially expressed proteins was subject to subsequent analysis using Toppgene suite (http://toppgene.ccmc.org) and a p value less than 0.05 was considered significant. The functional interaction network analysis of the identified proteins was analyzed by Cytoscape plugin GeneMANIA (www.genemania.org).
Estimation of ANGT levels
Plasma levels of ANGT were measured using human ANGT ELISA kit (WUHAN EIAab Science co. Ltd, China) according to the manufacturer's instructions. In brief, 100 μL of standard and plasma samples were added to the assay plate and incubated for 2 hours at 37°C. Furthermore, 100 μL of detection reagent A was added to each well and incubated for another 1 hour at 37°C. Subsequently, reagent B was added to each well with incubation for 30 minutes at 37°C. The plate was washed and incubated for 15 minutes with substrate solution at 37°C. The reaction was stopped by adding stop solution, the absorbance was measured at 450 nm, and concentration of ANGT was determined using standard curve of ANGT.
Measurement of NOx (nitrate+nitrite) levels in plasma
The blood plasma was passed through 10 kDa cutoff membranes and the filtrate was used for measuring NOx levels using nitrate/nitrite colorimetric assay kit (Cayman Chemical, Ann Arbor, MI) according to the manufacturer's instructions.
Estimation of cGMP levels
The plasma cGMP concentration was estimated using cyclic GMP EIA kit (Cayman Chemical, Ann Arbor, MI) according to the manufacturer's instructions.
Validation of protein expression by immunoblot analysis
Plasma containing 50 μg of total protein was separated on 10% SDS-PAGE gels and transferred onto nitrocellulose membranes. The membranes were blocked in 5% nonfat milk in PBS containing 0.1% Tween-20 (PBST) overnight at room temperature, washed and probed with C3, C4B, C1S, and β-tubulin (Santa Cruz Biotechnology, Inc.) Ang II (Novus Biologicals), eNOS, and phosphorylated eNOS (Ser1177) (Cell Signaling Technologies, Danvers, MA) at room temperature for 2 hours. Subsequently, the membranes were washed with PBST thrice for 5 minutes each and were then incubated with its respective secondary antibody for 1.5 hours at room temperature. The membranes were washed and observed on X-ray films using chemiluminiscent peroxidise substrate (Sigma, St. Louis, MO). The autoradiograms were scanned and densitometry analysis was performed with Image J software (http://rsbweb.nih.gov/ij/). All measurements were normalized to β-tubulin and are expressed as mean control density.
Statistical analysis
All biochemical data are represented as mean ± SEM of three independent experiments. Statistical analysis of ELISA and immunoblot was done using student's t-test and a p value less than 0.05 was considered significant. All analysis was performed using GraphPad Prism software (version 5).
Results
Comparative proteome analysis between Ladakhi natives and sea-level controls
Comparative plasma proteome profiling of Ladakhi natives by 2-DGE combined with MS analysis resulted in identification of 36 differentially expressed proteins in comparison with sea-level residents (Fig. 1 and Table 1). Among the identified proteins, 16 proteins were found to be downregulated, whereas 20 proteins were upregulated in Ladakhi natives as compared with sea-level residents (Table 1). Three proteins namely apolipoprotein A-IV (72%), inter-alpha-trypsin inhibitor heavy chain H4 (58%), and CD5 antigen-like protein (75%) were more abundant in Ladakhi natives than in sea-level individuals. Proteins belonging to blood coagulation cascade namely fibrinogen beta chain (19%), alpha-2-macroglobulin (9%), and fibrinogen gamma chain (7%) were found in lower levels in Ladakhi natives. Proteins such as NADH dehydrogenase [ubiquinone] 1 alpha subcomplex subunit 2 (NDUA 2), glutathione peroxidase 3 (GPx 3), antithrombin-III (ANT 3), ANGT, apolipoprotein M, apolipoprotein L1, and apolipoprotein E were found to be lower in abundance than in sea-level controls (Fig. 1). Interestingly, significant lower levels of vasoconstrictor ANGT was observed in Ladakhi natives (40%, p < 0.05), suggesting downregulation of RAS.
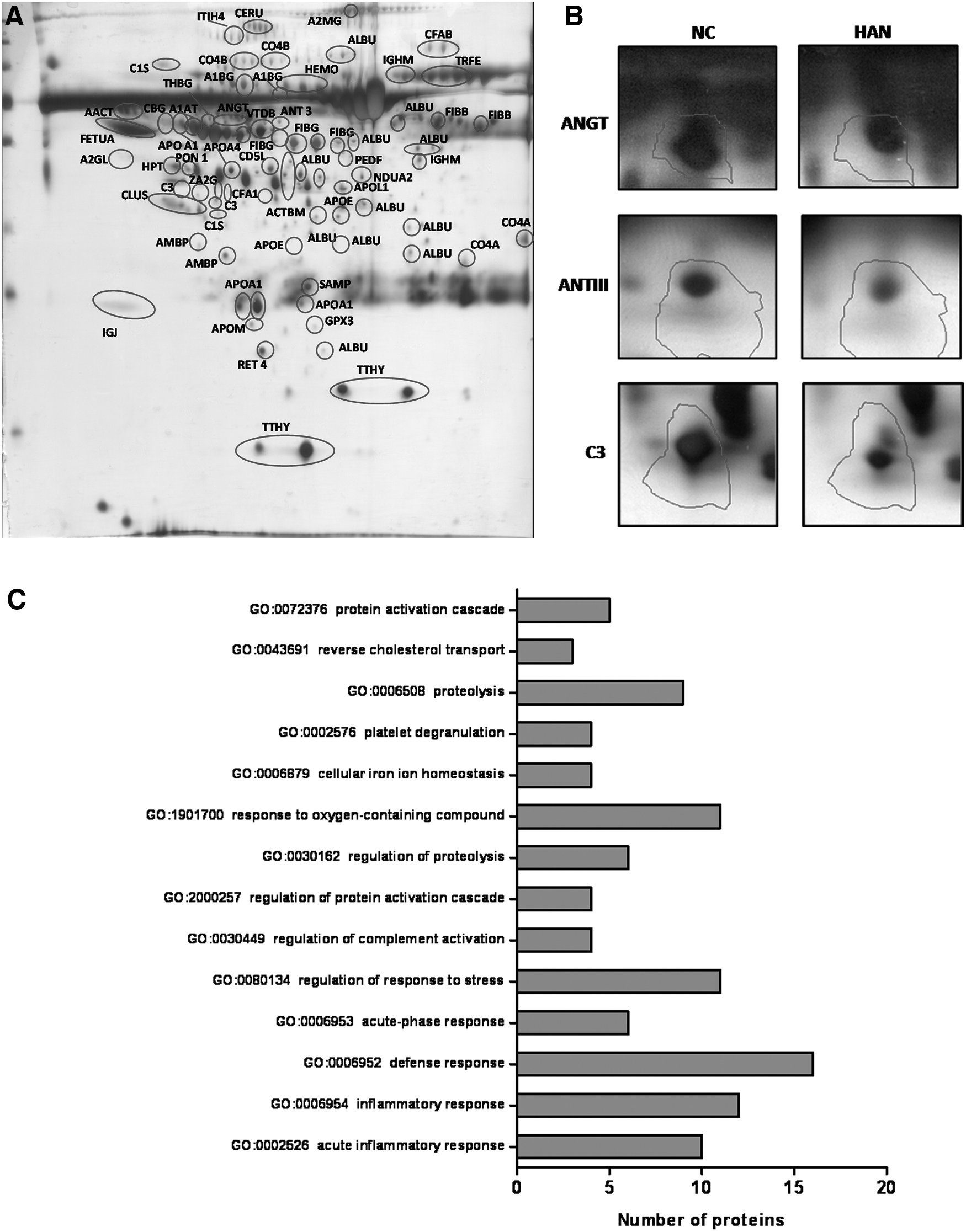
Identification of differential plasma proteins in Ladakhi natives.
Accession No.: Swissprot database accession number. pI represents observed isoelectric point and molecular weight in kDa, respectively, of the matched protein. Score: MASCOT probability-based MOWSE (molecular weight search) score calculated for peptide mass fingerprinting. Protein score is −10 × log (p), where p is the probability that the observed match is a random event and greater than 56 are significant (p < 0.05). Sequence coverage: Percentage of identified sequence to the complete sequence of the known protein.
% Change represents increment (↑) and reduction (↓) in high-altitude native versus control.
MS, mass spectroscopy; MS/MS, tandem mass spectroscopy.
Functional classification of the identified proteins
The differentially expressed proteins were imported to Toppgene suite and a corresponding gene list was created from the protein list. Toppgene data explicitly in their prioritization approach for identifying the significant biological processes (p < 0.05). Gene ontology represented in biological processes revealed perturbations belonging to defense response (GO:0006952), platelet degranulation (GO:0002576), proteolysis (GO:0006508), regulation of complement activation (GO:0030449), response to oxygen containing compound (GO:1901700), cellular iron ion homeostasis (GO:0006879), and reverse cholesterol transport (GO:0043691) categories (Fig. 1C).
Blood coagulation and complement proteins are less abundant in Ladakhi natives
We identified proteins belonging to blood coagulation (TETN, FIBG, AMBP, ANT3, A2MG, and FIBB) and complement cascade (C1S, C3, C4A, C4B, and CFAB) for Ladakhi natives (Table 1). Interestingly all these proteins were found to be lower in Ladakhi natives than in normobaric controls except tetranectin (TETN). Our Western blot studies also revealed 1.7-fold (p < 0.05), 3.6-fold (p < 0.001), and 2.2-fold (p < 0.001) lower levels of C1S, C3, and C4B in Ladakhi natives, respectively, than in normobaric controls (Fig. 2). In addition, we have also observed 19% lower levels of ANT 3 for Ladakhi natives (Fig. 1B).
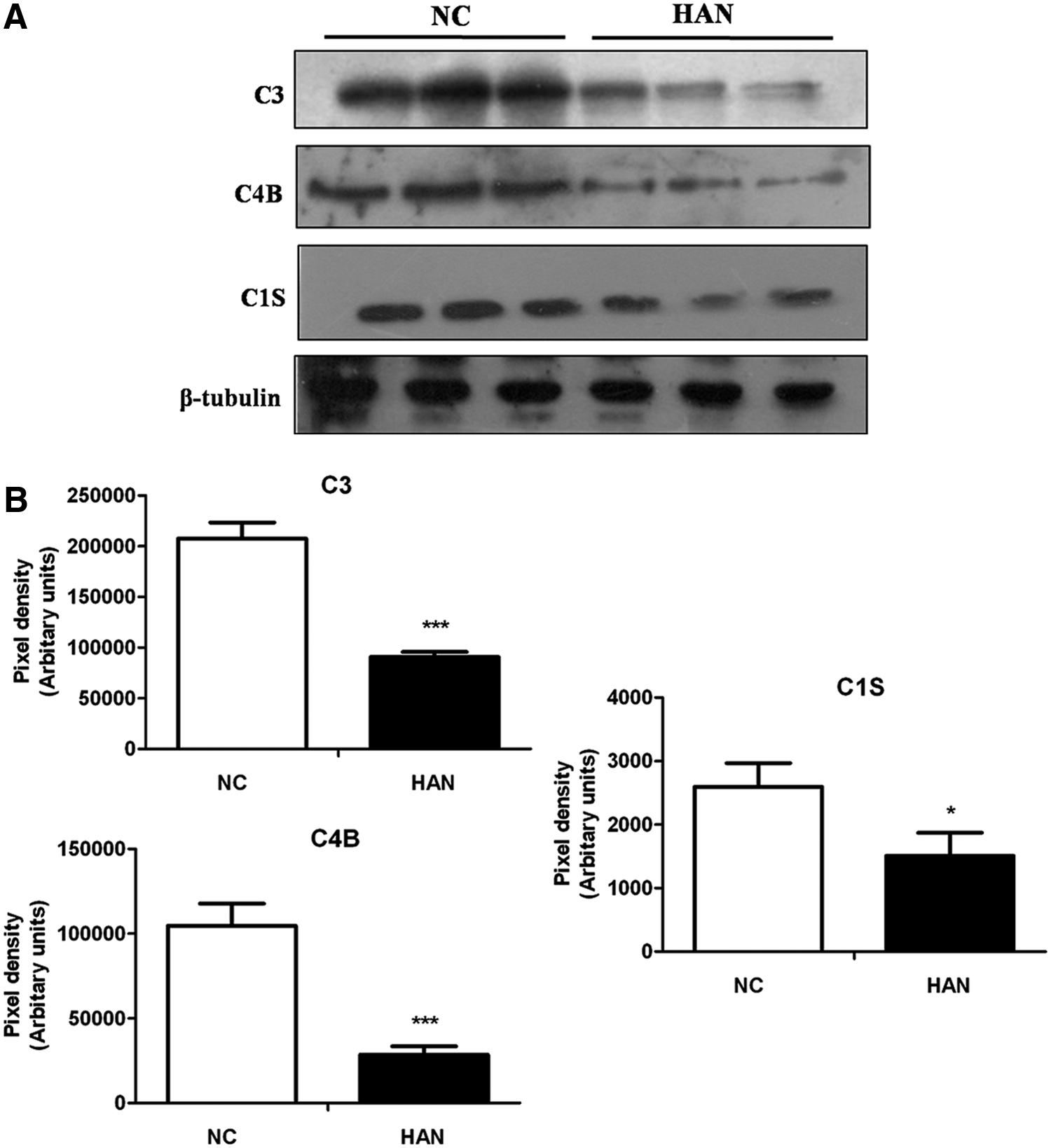
Western blot analysis for validation of expression levels of complement proteins.
Evaluation of RAS in Ladakhi natives
The proteins of the RAS play an important role in the regulation of vascular tone. ANGT the precursor of this pathway undergoes cleavage and enzymatic conversion to form the active mediator Ang II. Our proteomics studies revealed 33% (p < 0.05) lower levels of ANGT in Ladakhi natives than in sea-level controls. In corroboration, both ELISA-based and Western blot-based estimation of plasma ANGT and Ang II levels also revealed 24% (p < 0.01) and 23% (p < 0.05) lower levels in Ladakhi natives, respectively, than in normobaric controls (Fig. 3). Since NO counteracts RAS, we measured the eNOS and phosphorylated eNOS (Ser1177) levels by immunoblotting (Fig. 3B). Interestingly we observed increased levels of eNOS (1.4-fold, p < 0.05) and phosphorylated eNOS (Ser1177; 1.3-fold, p < 0.05) in Ladakhi natives, indicating increased NO synthesis (Fig. 3C). In addition, ELISA-based estimation of NO availability markers NOx (5.9-fold, p < 0.001) and cGMP (1.4-fold, p < 0.01) also corroborated higher levels of NO metabolites in Ladakhi natives (Fig. 4). These cumulative results suggest that Ladakhi natives exhibit higher plasma levels of functional eNOS and NO metabolites than normobaric controls.
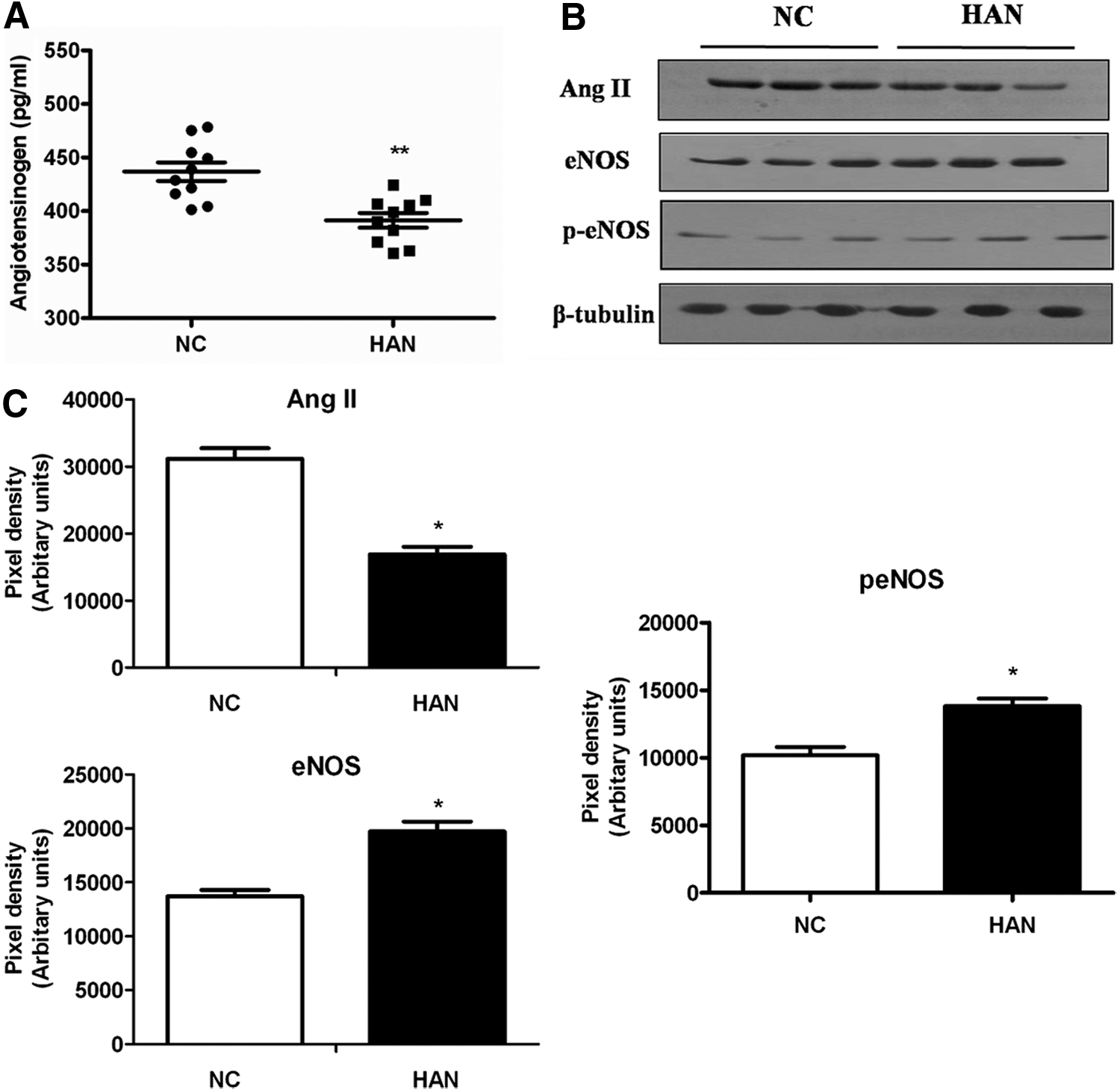
Cross-talk between RAS and NO pathway.
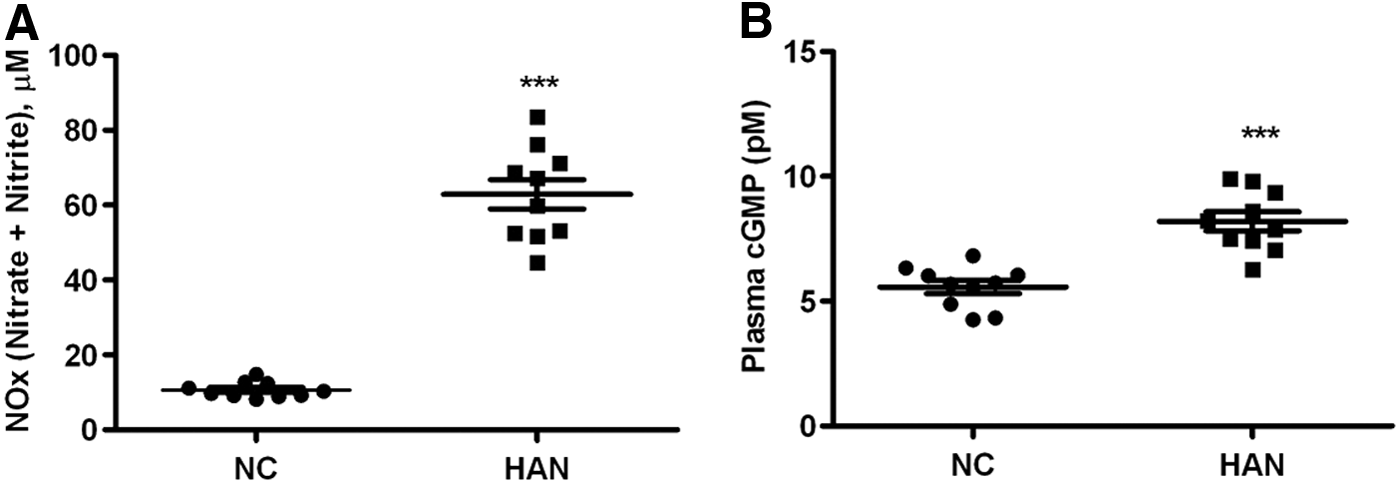
Increased levels of NO biomarkers in Ladakhi natives.
Discussion
Hypobaric hypoxia is an unavoidable environmental stress experienced at high altitude both by high-altitude natives and sojourners. Individuals ascending to high altitude suffer from several respiratory and neurological disorders, including acute mountain sickness, HAPE, and high altitude cerebral edema (HACE) (Wilson et al., 2009), whereas humans residing at high altitude for generations have adapted well to this environment through natural selection (West, 1982). To identify molecular pathways promoting survival and performance of high-altitude natives, we carried out a plasma proteomics study for Ladakhi natives permanently residing at 3520 m and compared it with that of sea-level individuals. This comparative approach resulted in the identification of 36 differentially expressed proteins in Ladakhi natives.
The identified proteins fall into the major classes of blood coagulation and blood pressure regulation, complement activation, acute phase proteins, antioxidant proteins, and transport proteins. We observed significant low levels of blood coagulation proteins in Ladakhi natives. Remarkably, the Ladakhi natives exhibited 19% lower levels of ANT 3, a glycoprotein that plays an important role in inhibiting the serine proteases involved in coagulation cascade as compared with sea-level individuals. Hypobaric hypoxia exposure results in increased blood coagulation, contributing to deep vein thrombosis and embolism (Singh and Chohan, 1972; Anand et al., 2005; Mieske et al., 2010; Chen et al., 2012; Pawlinski, 2014). Increased procoagulatory state with increased altitude has been reported for high-altitude sojourners (Pichler Hefti et al., 2010). The lower levels of ANT 3 observed in this study suggest attenuation of coagulatory pathways in Ladakhi natives despite hypobaric hypoxia. We also observed lower levels of complement pathway proteins (C1S, C3, C4A, C4B, and CFAB) for Ladakhi natives. The complement system, composed of more than 30 proteins, responds by means of recognition and activating mechanisms to foreign proteins, tissue injury, apoptosis, and necrosis. The classical, lectin, and alternative complement activation pathways converge on the C3 component, which results in initiation of inflammation through recruitment and activation of inflammatory cells (Zipfel and Skerka, 1994; Markiewski and Lambris, 2007). Western blot-based estimation of C1S, C3, and C4B also revealed lower levels of these proteins in Ladakhi natives. In a similar study, Ahmad et al. (2013) have also reported lower levels of C3 and C4A for high-altitude natives. Chronic hypoxia-induced complement activation plays a pathophysiological role in the development of pulmonary arterial hypertension (PAH) and genetic deletion of C3 results in significant attenuation of PAH during chronic hypoxia (Bauer et al., 2011). Studies on animal models have reported significant contribution of complement system in promoting vascular leakage and hypoxia-induced thrombosis (Bossi et al., 2011). It is noteworthy that molecular intercommunication exists between the coagulation and fibrinolysis cascade members thrombin, human coagulation factors XIa, Xa, IXa, and plasmin with the central complement components C3 and C5 during inflammation (Amara et al., 2010; Oikonomopoulou et al., 2012). The observed lower abundance of both coagulation and fibrinolysis cascade proteins in Ladakhi natives suggests altered vascular homeostasis that enables Ladakhi natives to survive under hypoxia.
Exposure to chronic hypoxia results in elevation of blood pressure mediated through the RAS (Landmesser et al., 2002; Nangaku and Fujita, 2008; Foster et al., 2010). The most powerful biological product of this pathway is Ang II, which directly constricts vascular smooth muscle cells, enhances myocardial contractility, and plays a primary role in the pathogenesis of hypertension through AT1 receptor (Grassi et al., 1996). ANGT is a single chained, 61 kDa glycoprotein constitutively secreted by hepatic cells into the circulation and is the precursor for Ang II throughout the circulation (Kobori et al., 2007). Our present plasma proteomics-based studies revealed 33%–49% decreased levels of plasma ANGT in Ladakhi natives (Fig. 1B). Our subsequent estimation of plasma levels of ANGT as well as Ang II also revealed lower levels of both these proteins in Ladakhi natives. Analyzing genomic regions of Tibetans and Andeans, Bigham et al. have reported positive natural selection on HIF pathway, RAS, and globin family genes (Bigham et al., 2010). Our present results along with these studies suggest that lower levels of RAS proteins are an effective method to attenuate hypoxia-induced vasoconstriction by Ladakhi natives.
The vasoconstrictor Ang II and vasodilator NO signaling pathways mutually regulate each other in a feedback mechanism. Both animal and human studies have reported functional interplay between Ang II and NO–cGMP pathways in controlling vascular structure and function (Yan et al., 2003). Since we observed lower levels of ANGT and Ang II for Ladakhi natives, we measured eNOS protein levels along with plasma marker for NO activity (cGMP), and NO availability (nitrate and nitrite levels). We observed 1.4-, 1.3-, 1.4-, and 5.9-fold higher plasma levels of eNOS, phosphorylated eNOS (Ser1177), cGMP, and NOx, respectively, for Ladakhi natives, suggesting high NO production and availability. NO and associated metabolites play an integral role in human response to hypoxia (Levett et al., 2011). Higher levels of eNOS-mediated NO synthesis and NO–cGMP-mediated vasodilation represent important mechanisms for effective Tibetan pattern of high-altitude adaptation (Erzurum et al., 2007; Beall et al., 2012). Similar higher plasma levels of nitrate and nitrite have also been reported for Indian Himalayan highlanders (Qadar Pasha et al., 2001; Ahsan et al., 2004, 2005; Beall et al., 2012). Our present results along with these previous studies suggest that Ladakhi natives also offset ill effects of high altitude by virtue of higher plasma NO and associated metabolite levels similar to high-altitude native Tibetans.
This study suffers from limitations of using 2D gels for identification of plasma proteins. The smaller dynamic range of this technique does not permit identification of a wide range of plasma proteins. Moreover, this technique also suffers from disadvantages such as high sample requirement, reproducibility, labor intensive and nonsuitability for automation. In this study, we have used three technical replicates of each sample and have analyzed the data with statistical stringency to overcome these limitations. The use of high throughput techniques for plasma profiling might reveal additional regulatory pathways for high-altitude survival as well as adaptation. The other disadvantages of this study are the use of limited number of volunteers and nonapplication of multiple correction testing. In future, such discovery proteomics studies with large number of volunteers may elucidate additional mechanisms for high-altitude adaptation. Although the present study provides pieces of evidence for possible interaction between NO and Ang II signaling pathways, elucidation of precise molecular interactions is warranted.
Conclusion
This study reports lower levels of complement and coagulation proteins for Ladakhi natives. The observed lower levels of ANGT and Ang II with concomitant higher levels of eNOS and phosphorylated eNOS (Ser1177) suggest higher NO availability for Ladakhi natives. This was further supported by higher levels of NO metabolites (nitrate, nitrite, and cGMP), indicating that Ladakhi natives follow a Tibetan pattern of high-altitude adaptation. Our present results will augment the current understanding of human adaptation to high altitude.
Footnotes
Acknowledgments
This work is supported by Defence Research and Development Organization (DRDO), Ministry of Defence, Government of India. The authors thank Director, DIPAS Dr. Shashi Bala Singh for supporting this study and all the volunteers for their participation. G.P. is a recipient of the Senior Research Fellowship (SRF) from CSIR, Government of India, and A.G. is a recipient of the DST-Inspire Fellowship from the Government of India.
Author Disclosure Statement
No competing financial interests exist.