
Abstract
Abstract
Nehra, Sarita, Varun Bhardwaj, Santosh Kar, and Deepika Saraswat. Chronic hypobaric hypoxia induces right ventricular hypertrophy and apoptosis in rats: therapeutic potential of nanocurcumin in improving adaptation. High Alt Med Biol. 17:342–352, 2016.—a sustained work load on the right heart on ascent to high altitudes promotes right ventricular hypertrophy (RVH), which eventually undergoes decompensation and promotes pathological damage. However, the exact set of events leading to damage remains unidentified. Curcumin is a natural antioxidant and antihypertrophic agent, but it has poor biostability. Nanotized curcumin (nanocurcumin) has emerged as a promising agent with improved biostability while retaining the therapeutic properties of curcumin. The present study aimed at analyzing the therapeutic properties of nanocurcumin in ameliorating cardiac damage due to chronic hypobaric hypoxia (HH)-induced RVH in comparison to curcumin. Sprague-Dawley rats exposed to HH (25,000 feet, effective oxygen fraction in air [FIO2] ∼0.08, temperature 28°C ± 1°C, relative humidity 55% ± 2% for 3, 7, 14, and 21 days) developed RVH with increased interstitial collagen content, Fulton's index, and cardiomyocyte cross-sectional area while upregulating atrial natriuretic peptide. Tissue damage due to apoptotic cell death was evident by cytochrome-c/caspase-3 activation and TUNEL assay. Concomitant modulation of cyclic guanosine monophosphate (cGMP)/cGK-1, calmodulin-dependent protein kinase II (CaMkinase II), and intracellular calcium levels with increased free radical-induced damage and lipid peroxidation further contributed to the right heart pathology. Nanocurcumin supplementation decreased HH-induced RVH and apoptosis while modulating cardiac cGMP/cGK-1 signaling, and maintaining CaMkinase II, intracellular calcium levels and redox status better than curcumin. Nanocurcumin-mediated antiapoptotic effects might have benefited residents and sojourners at high altitude in preventing hypoxic cardiac damage.
Introduction
H
RVH and ventricular remodeling are accompanied by upregulation of atrial natriuretic peptide (ANP), a hormone and marker of hypertrophy (Sadoshima and Izumo, 1993). Studies have shown that ANP regulates hypertrophy, in part, by modulating cyclic guanosine monophosphate (cGMP) accumulation (Tsai and Kass, 2010). ANP binds to the extracellular domain of GC-A (guanylyl cyclase A), which then increases intracellular levels of cGMP. Under stresses that may cause RVH, ANP-mediated cGMP accumulation activates a downstream cytoplasmic cascade that protects against hypertrophy by initiating a complex set of events. In vitro studies have shown that cGMP acts as a negative modulator of hypertrophy (Calderone et al., 1998; Langenickel et al., 2006). In addition, studies have shown that in patients with heart failure, acute treatment with exogenous ANP is effective in maintaining redox balance (Shono et al., 2007). However, the effects of chronic intrinsic release of ANP in cardiac remodeling and damage have not been investigated. Moreover, the modulatory effect of chronic stress on cGMP levels and its association with RVH remain unknown.
Curcumin, a natural polyphenolic alkaloid, possesses remarkable antioxidant, antiapoptotic, and anti-inflammatory properties (Ghosh et al., 2010; Basnet and Skalko-Basnet, 2011; Dadhaniya et al., 2011). However, rapid metabolism and gastric clearance decrease the bioavailability and biostability and render curcumin an ineffective therapeutic agent (Begum et al., 2008). Recently, nanotization of curcumin to “nanocurcumin” has been shown to highly improve the therapeutic properties of curcumin in a number of diseases (Bisht et al., 2007; Flora et al., 2013; Nehra et al., 2015b), but the effect of nanocurcumin in ameliorating HH-induced RVH has not yet been investigated.
Based upon these findings, we hypothesized that chronic HH might induce RVH and pathological damages in adult male Sprague-Dawley rats. We have previously shown that nanocurcumin protects rodent cardiomyoblasts and primary human ventricular cardiomyocytes from hypoxia-induced hypertrophy and associated damage (Nehra et al., 2015a, 2015b). In the present study, we investigated whether exposure to chronic HH promotes cardiac pathological damage in Sprague-Dawley rats and assessed the improvement in therapeutic potential of nanocurcumin in their amelioration over that of curcumin itself. We aimed also to identify the molecular signaling underlying HH-induced RVH and how nanocurcumin might be superior to curcumin.
Materials and Methods
Ultrapure molecular grade chemicals were used in the whole study. All chemicals were purchased from Sigma-Aldrich or otherwise stated. Curcumin was purchased from Sigma-Aldrich (C7727). Nanocurcumin (average particle size 200 nm, zeta potential −131 mV, percentage oral bioavailability 0.5 ± 0.060) was obtained as a kind gift from Professor Santosh Kar, KIIT University, Odisha, India (Akhtar et al., 2012).
Study design and experimental animals
Age-matched adult male Sprague-Dawley rats (220 ± 10 g) (6–8 weeks) were used for all the experiments (Xi and Serebrovskaya, 2012). The experimental animals were kept in the Institute's Experimental Animal Facility in clean cages. Animals had equal access to water and standard chow (Lipton India Pvt. Ltd.), with proper maintenance of 12 hours light and 12 hours dark cycle at 28°C ± 2°C temperature and relative humidity (55% ± 2%). All experiments were approved by the Institutional Animal Ethics Committee in accordance with the Committee for Purpose of Control and Supervision of Experiments on Animals (27/1999/CPCSEA), Government of India.
Animals were randomly divided into six groups (n = 8): normoxia vehicle control (N), normoxia plus curcumin (N+C), normoxia plus nanocurcumin (N+NC), hypobaric hypoxia vehicle control (HH), hypoxia plus curcumin (H+C), and hypoxia plus nanocurcumin (H+NC). Sterile and neutral phosphate-buffered saline (PBS) was used as a vehicle.
Simulation of animals to HH
Animals were exposed to extreme HH to assess the effective dose of nanocurcumin and tolerance to hypoxic stress. In addition, circulating levels of vascular endothelial growth factor (VEGF) were assessed as an important mitogen for hypertrophic growth and a key marker of angiogenesis (Conway et al., 2001). Tolerance to a decrease in oxygen availability was measured by assessing the time taken by the animal at an extreme high altitude of equivalent to 32,000 feet (FIO2 ∼0.06, temperature 26°C ± 1°C, relative humidity 50% ± 2%) in an animal decompression chamber (SevenStar Instruments) for onset of a hyperventilatory response (expressed as hyperventilatory time [HT]), measured in minutes beyond which the animals start gasping. Different concentrations of nanocurcumin, that is, 5, 10, 20, 50, and 100 mg/kg b.w., were prepared in neutral PBS as previously described (Nehra et al., 2015a). The animals were supplemented with nanocurcumin/curcumin or vehicle just before HH exposure by oral gavage and screened for maximum HT and VEGF release in plasma. The dose of nanocurcumin at which animals showed maximum HT and maintenance of VEGF levels was chosen as optimum for further experiments.
For exposure to acute, subacute, and chronic HH, the animals were initially allowed to acclimatize to mild HH (FIO2 ∼0.117) at a simulated ambient pressure equivalent to 15,000 feet, at 28°C ± 1°C, relative humidity 55% ± 2% for 24 hours in an animal decompression chamber (Ballice Instruments), and thereafter exposed to extreme HH for assessment of RVH at an ambient pressure equivalent to 25,000 feet altitude (FIO2 ∼0.08, temperature 28°C ± 1°C, relative humidity 55% ± 2%) for 3, 7, 14, and 21 days or otherwise kept in normoxia. Treatment with curcumin/nanocurcumin/vehicle was started on first day of HH exposure and continued until the completion of HH exposure. The rate of change in ambient pressure was maintained ∼35 mmHg/min to avoid any tissue injury from a sudden drop or increase in ambient pressure. The animal decompression chamber was opened every 24 hours for 10 minutes at 10:00 AM for dosing, changing of bedding, and food and water replenishment.
Tissue and blood harvesting
At the completion of the experiments, the animals were sacrificed by an overdose of ketamine (80 mg/kg b.w.) and xylazine (10 mg/kg b.w.), intraperitonial in under 15 minutes to avoid reoxygenation–reperfusion injury and perfused with ice-cold PBS (n = 8) or 4% methanol-free neutral paraformaldehyde (PFA, in PBS) (n = 5). Freshly isolated hearts were immediately snap-frozen in liquid nitrogen (−196°C) and stored at −80°C until further use. Blood was isolated by left ventricular puncture in K2-EDTA-coated vacutainers (367863, B.D.) and centrifuged at 8000 g at 4°C for isolation of nonhemolyzed plasma.
For molecular studies, the stored hearts (n = 8) were thawed and homogenized in ice-cold cytosolic extraction buffer (0.5 mM sucrose, 10 mM HEPES, 1.5 mM MgCl2, 10 mM KCl, 10% glycerol, 1 mM EDTA, 1 mM DTT), with a protease inhibitor cocktail (5 μL/mL buffer) (MP Biomedicals) homogenized at 8000 rpm using automatic homogenizer (POLYTRON PE1200E) and centrifuged at 12,000 g for 30 minutes at 4°C. The supernatant containing cytosolic proteins was collected and stored immediately at −80°C until further use.
Assessment of RVH
Animals were screened for RVH development after 3, 7, 14, and 21 days of HH. For morphometric assessment, freshly isolated hearts were carefully incised using a sterile scalpel to remove large vessels and separate the right ventricle (RV), left ventricle with septum (LV+S), and atria. The sectioned tissues were washed twice in PBS to remove excess tissue and liquid and were weighed on a high-precision electronic balance (BSA224S-CW; Sartorius). The Fulton's index of RVH was expressed as RV/LV+S and RV/BW (normalized to animal body weight, expressed as g/kg b.w.) to assess extent of RVH. Circulating and tissue concentration or content of ANP was performed by ELISA (EIA05048R) and western blot, respectively (Wang et al., 2010). Cardiomyocyte cross-sectional area was measured as previously described (Tracy and Sander, 2011).
Since cGMP-cGK-1 signaling is an important mediator of cardiac hypertrophy, tissue levels of intracellular cGMP were measured using the commercially available ELISA kit (CG200; Sigma-Aldrich) and further confirmed by western blot analysis of cGK-1 levels. In addition, since some circulating cytokines play a crucial role in cardiac hypertrophy-mediated pathological damages; circulating concentrations of tumor necrosis factor-α (TNFα) (BD Biosciences; 558535) and interleukin (IL)-6 (550319; BD OptEIA) were measured according to the manufacturer's instructions.
Measurement of blood pressure and heart rate
Measurement of systolic blood pressure (sBP) and heart rate (HR) was performed using noninvasive plethysmography by a tail-cuff method (BioPac; MP150) with calculation of mean arterial pressure as described by Britto et al. (1997). Tail plethysmography has been previously used as a reliable method for measuring sBP and HR with minimized operator interference and animal discomfort (Ghosh et al., 2010). Briefly, immediately after completion of HH exposure, the animals were settled and allowed to relax in animal restrainers (SevenStar Instruments) and allowed to rest for 10–15 minutes. The tails of animals were cleaned by cotton swab, the analytical probe was carefully inserted around the tail, and BP and HR were recorded. The final values for sBP and HR were represented as a mean of three consecutive readings.
Assessment of HH-induced apoptosis in cardiac tissue
HH-induced apoptosis was assessed by estimating tissue expression of cytochrome-c, caspase-3, and active caspase-3 by immunohistochemistry and western blot. TUNEL assay was performed in heart cryosections using a commercially available kit (TUNEL-Apoptosis detection kit, 17-141; Millipore) performed according to the manufacturer's instructions.
Markers of cardiac injury
Assessment of glucose, the myocardial isoform of creatine kinase (CK-MB), and lactate dehydrogenase (LDH) are critical biomarkers of injury with RVH (Vanbilsen et al., 2004). Commercially available kits were used for estimation of plasma glucose concentration (Gluc-PAP; Randox), CK-MB (CK-MB 1296; Randox), and LDH (LD 40I; Randox) activity. Total intracellular calcium was estimated using the Quantichrome™ Calcium Analysis Kit (Bioassay Systems) and confirmed by western blot of calcium/calmodulin-dependent protein kinase II (CaMkinase II) in RVs.
Analysis of oxidative damage
Assessment of HH-induced oxidative damage was done by analyzing free radical-induced damage and lipid peroxidation. Estimation of free radical generation by excessive reactive oxygen species (ROS) formation was done using the dichlorofluorescein diacetate (DCFH-DA) assay. Briefly, the freshly prepared RV lysates were incubated with 100 μM DCFH-DA and incubated in the dark for 15 minutes at room temperature (Cathcart et al., 1983). Fluorescence was optically measured at excitation wavelength 485 nm and emission at 530 nm (FLUOStar Omega; BMG LABTECH).
Assessment of lipid peroxidation was performed using the method described by Utley et al. (1967). Briefly, the RV homogenates were incubated with 20% trichloroacetic acid and 0.67% thiobarbituric acid in a hot water bath at 80°C–90°C for 40 minutes until a pink color developed. The samples were allowed to cool at room temperature, centrifuged at 4000 g, and absorbance of supernatant was read optically at 531 nm (FLUOStar Omega; BMG LABTECH).
Histopathology and immunohistochemistry
The hearts were perfused with 4% methanol-free neutral PFA. The fixed hearts were cryosectioned (25 μm) microtomy (5 μm) for immunohistochemistry and histopathology, respectively. Masson's trichrome staining was performed for assessment of RVH and visualization of collagen accumulation. Tissue collagen accumulation was quantitatively assessed by a hydroxyproline accumulation assay (Sigma-Aldrich; MAK008) according to the manufacturer's instructions.
Every consecutive fifth section was taken for immunohistochemistry. Briefly, the cryosections were incubated with 4% blocking buffer (4% bovine serum albumin, in PBS, 0.3% Triton-X-100, 0.03% H2O2) at room temperature for 60–90 minutes and washed three times with PBST (PBS with 0.1% tween-20) for 10 minutes. The sections were incubated with mouse monoclonal anti-caspase-3 (1:100, GTX108585; GeneTex) and rabbit polyclonal anti-cytochrome-c (1:100, GTX110543; GeneTex) overnight at 4°C. The sections were washed three times with PBS for 10 minutes and incubated in appropriate horseradish peroxidase (HRP)-conjugated anti-rabbit IgG (1:25,000; SantaCruz Biotechnology) for 2–2.5 hours at room temperature. The color was developed using DAB (00-2014; Invitrogen).
The cryosections were stained with Alexa fluor 488-Phalloidin (A12379; Molecular Probes) and counterstained with DAPI for visualization of changes in gross tissue architecture according to the manufacturer's instructions. The mounted sections were visualized under a bright field or fluorescent microscope (Olympus).
Immunoblot analysis
An equal amount of proteins (30 μg) from different experimental groups were resolved in 5%–10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis according to a method previously described (Nehra et al., 2015a). The nitrocellulose membranes were probed with rabbit polyclonal anti-caspase-3 (1:500; SantaCruz Biotechnology) (1:500), rabbit polyclonal anti-cleaved caspase-3 (1:1000, #9661; Cell Signaling Technology), mouse monoclonal anti-cytochrome-c (1:500; SantaCruz Biotechnology, sc13561), rabbit polyclonal anti-cGK-1 (1:100; SantaCruz Biotechnology, sc25429), mouse monoclonal anti-CaMkinase II (1:200; Invitrogen, 137300), and rabbit polyclonal anti-ANP (1:250; SantaCruz Bioctechnology, sc20158) for 2 hours. The membranes were then incubated with appropriate HRP-conjugated anti-mouse or anti-rabbit antibodies (1:25,000; SantaCruz Biotechnology) for 1.5–2 hours. The membranes were developed by using a chemiluminiscent substrate (Sigma-Aldrich) on X-ray films. Densitometric analysis of developed blots was performed using ImageJ software.
Data analysis
Data are expressed as mean ± SD for each experimental group (n = 8). The results were analyzed for statistical significance using one-way ANOVA followed by post hoc Bonferroni's analysis. Pearson's correlation was performed to evaluate the extent of relatedness between various parameters (ANP vs. RV/LV+S, RV/BW, CK-MB, and LDH and HT vs. circulating VEGF levels). Value was considered significant at *p ≤ 0.05 versus normoxia, $p ≤ 0.05 versus HH, and #p ≤ 0.05 versus H+C. Nonsignificant changes were depicted as ns.
Results
Optimization of nanocurcumin dose
The dose of nanocurcumin at which the animals demonstrated a maximum HT and restoration of circulatory VEGF levels was chosen as the optimum for further experiments. It was found that nanocurcumin at a minimum dose of 10 mg/kg b.w. yielded maximum HT and minimum VEGF release (p ≤ 0.05) in plasma with a strong negative correlation (r2 = 0.921). Increasing the dose of nanocurcumin beyond 10 mg/kg b.w. did not impart any further improvement in HT and restoration of VEGF levels. Thus, all experiments were performed at nanocurcumin dose at 10 mg/kg b.w. (Fig. 1) and compared to a similar dose of curcumin (10 mg/kg b.w.).
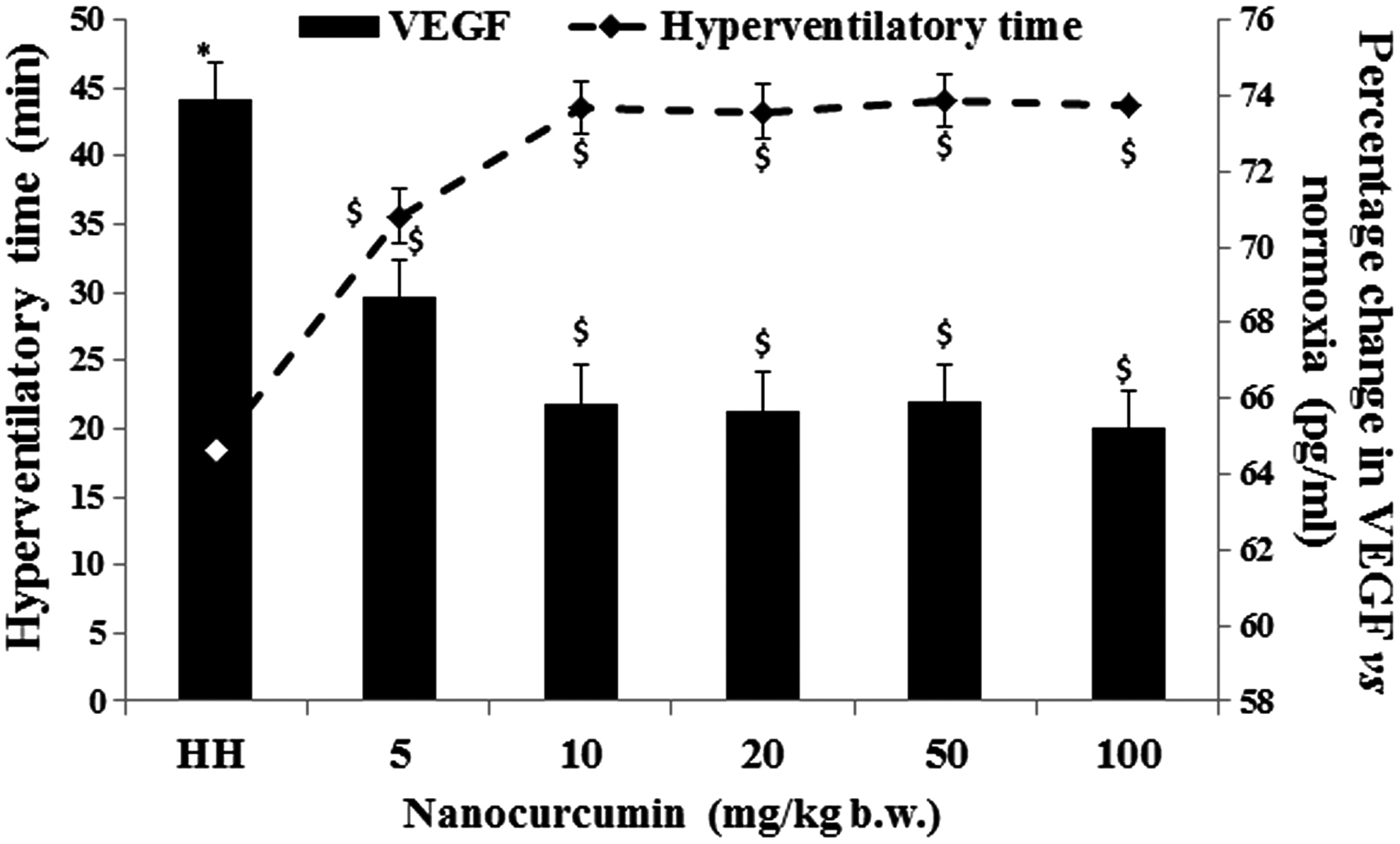
Figure showing effect of HH on HT and circulating VEGF levels in animals (n = 8). Nanocurcumin supplementation (10 mg/kg b.w.) demonstrated maximum HT and restored circulating VEGF levels with strong negative correlation (r2 = 0.921). Data represented as mean ± SD, values were considered significant at *p ≤ 0.05 versus normoxia and $p ≤ 0.05 versus HH. HH, hypobaric hypoxia; HT, hyperventilatory time; VEGF, vascular endothelial growth factor.
Time course study of induction of HH-induced RVH
The morphometric changes in hearts exposed to HH were found to be time dependent. The increments RV size compared to the LV and BW were used as quantitative indices of RVH and expressed in terms of RV/LV+S and RV/BW ratio. The hearts assumed more spheroid geometry with increments in muscle mass, collagen accumulation, and cardiomyocyte cross-sectional area (Fig. 2a–c) with chronic HH. Fulton's index corroborated these data indicating that RVH developed in HH as early as 3 days and reached a peak value at 21 days (p ≤ 0.05) (Fig. 2d, e).

Figure showing Masson's Trichrome staining (arrows) (40 × )
Nanocurcumin-supplemented animals showed effective modulation in heart morphometry, collagen accumulation, and cardiomyocyte cross-sectional area with HH (p ≤ 0.05) when compared to curcumin-treated animals (Fig. 2a–c). No significant change in Fulton's index was observed in animals supplemented with curcumin and nanocurcumin in normoxia (Fig. 2d, e). Collectively, the data clearly demonstrate evidence of an antihypertrophic property of nanocurcumin with HH compared to curcumin.
Changes in hemodynamic parameters and markers of cardiac injury
Animals exposed to acute (3 days) and chronic HH (21 days) showed increases in sBP and HR (3 days) compared to normoxia control animals, but the results were not statistically significant (Supplementary Table S1). In addition, plasma glucose did not change significantly with HH, however, plasma CK-MB and LDH activity increased up to 14 days and levels were maintained up to 21 days (Supplementary Table S2). Total intracellular calcium (Fig. 3a) and CaMkinase II expression levels (Fig. 3b, c) were increased in animals in chronic HH (p ≤ 0.05) compared to normoxia controls. Treatment with nanocurcumin significantly (p ≤ 0.01) restored CK-MB, LDH activity, and calcium content (p ≤ 0.05) compared to curcumin in HH. Although curcumin also imparted protection against cardiotoxicity and maintained calcium homeostasis and CaMkinase II, the results were more pronounced in nanocurcumin-supplemented animals.

Changes in total intracellular calcium levels
Chronic HH induces pathological RVH
Cardiac tissues stained with actin showed discrete structural remodeling with shrinking of tissue boundaries, which increased the interstitial space in chronic HH-exposed rats (Fig. 4a, c). These findings corroborated the increments in circulating ANP levels and tissue expression of ANP in animals exposed to HH within 3 days, which gradually reached peak values by 21 days (p ≤ 0.05) (Fig. 4b, d, e). There was a significant decrease in circulating and tissue ANP levels (p ≤ 0.05) in animals treated with nanocurcumin compared to vehicle or curcumin in chronic HH (Fig. 4b–e). In addition, the changes in cGMP and cGK-1 levels were found to be time dependent under hypoxic stress. Hypoxia-induced increase in cGMP levels was maintained up to 7 days and thereafter showed a sharp decline by 21 days (Fig. 5a). Similarly, high cGK-1 expression levels were observed with acute hypoxia (3 days), which decreased by 7 days and completely diminished by 21 days (Fig. 5b, c). Importantly, although ANP levels increased up to 21 days of HH, cGMP levels gradually declined after 7 days, clearly depicting transformation of HH-induced RVH from an adaptive response to a decompensated state that might be cGMP dependent. In addition, that chronic hypoxic stress-evoked metabolic damage was evident by an increase in LDH activity. Similarly, cardiac metabolic stress was demonstrated by a significant increase in CK-MB levels (p ≤ 0.05) (Supplementary Table S2). No significant change was observed in glucose levels (Supplementary Table S2). Importantly, nanocurcumin-supplemented animals showed significant (p ≤ 0.05) modulation of ANP, cGMP, and cGK-1 levels and reductions in LDH and CK-MB activity compared to curcumin- and vehicle-treated animals during chronic hypoxia.
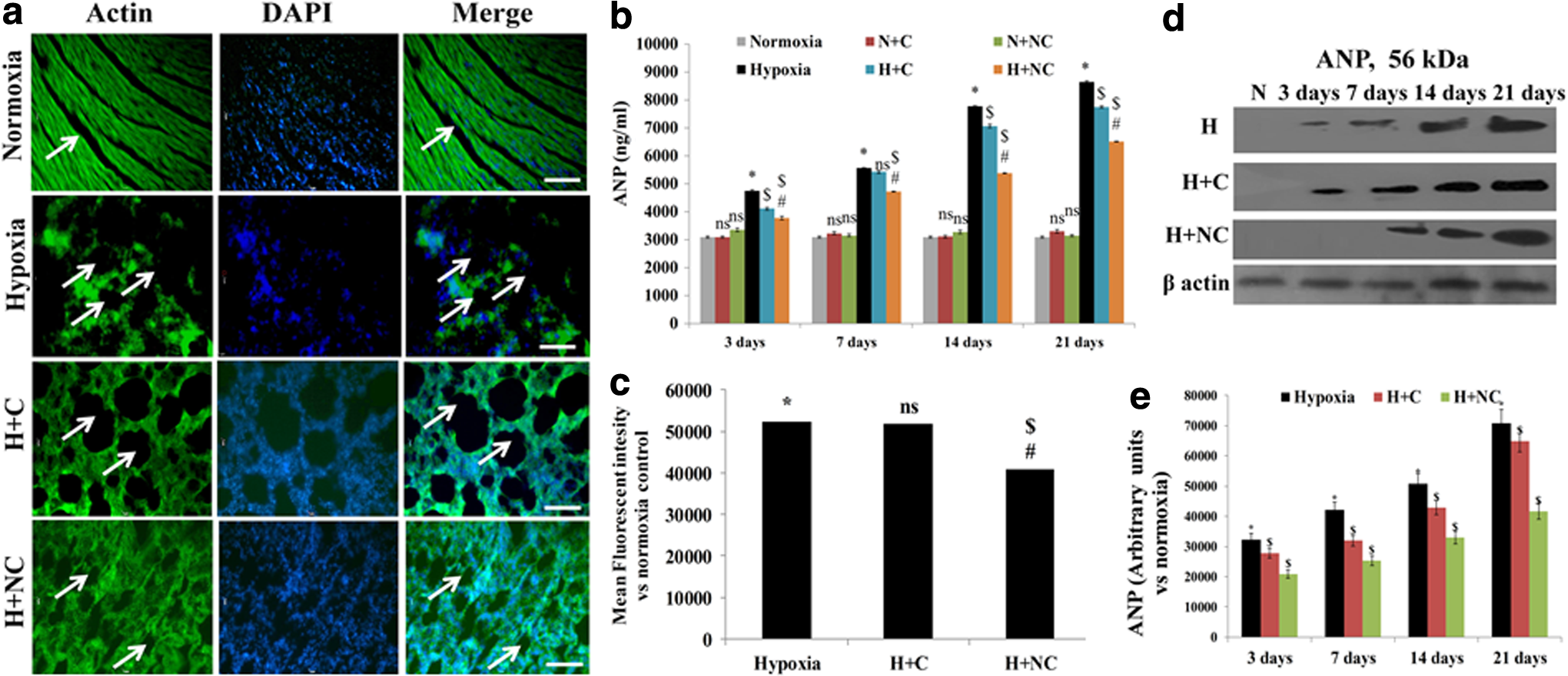
Figure showing cross sections of hearts exposed to chronic HH (21 days) with fluorescent staining for Phalloidin-488 to stain actin filaments to visualize tissue boundaries and interstitial space (arrows)
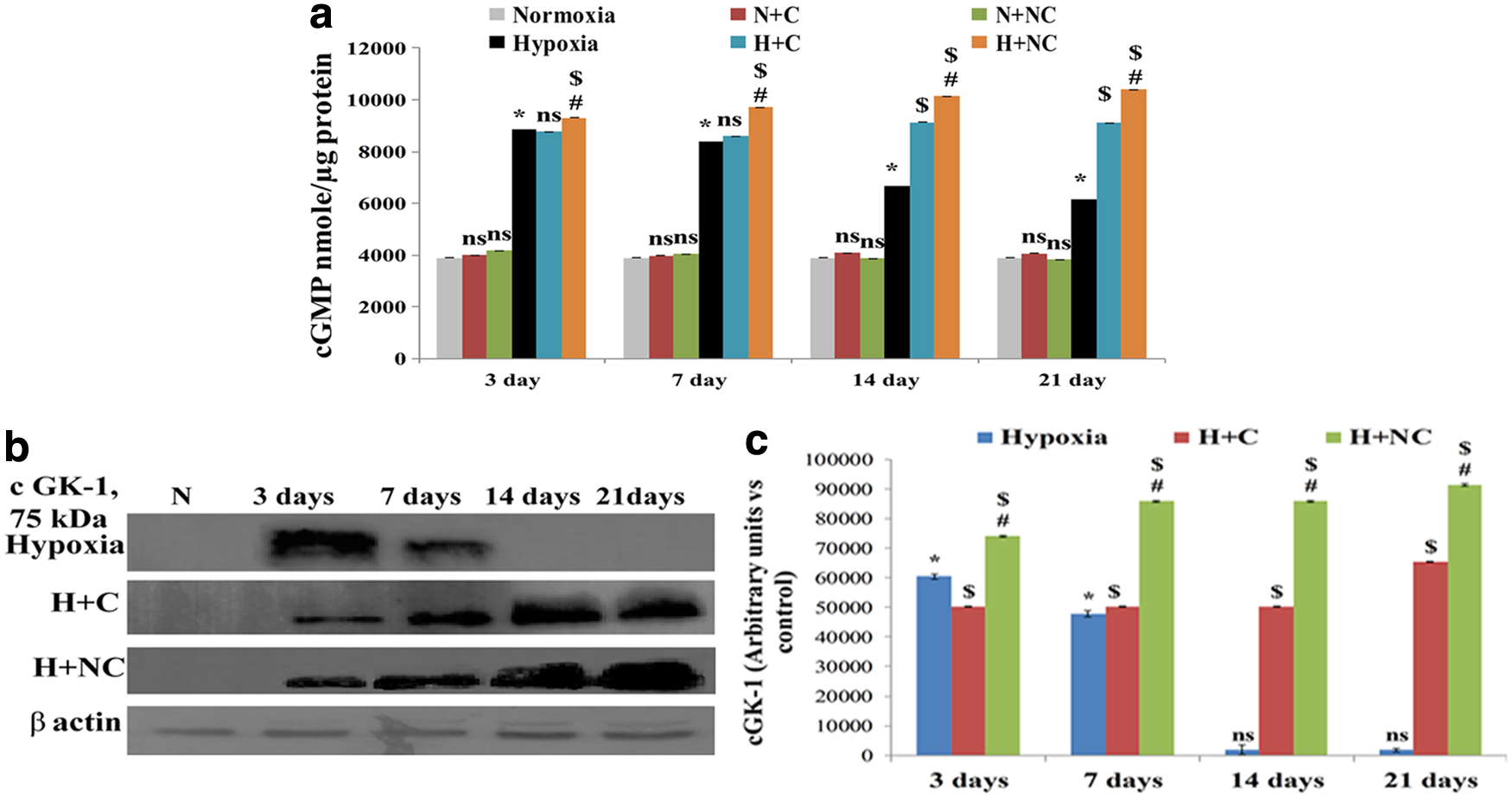
Figure showing changes in cGMP levels
The changes in circulating ANP levels were highly correlated with RV/LV+S (r2 = 0.907) and RV/BW (r2 = 0.981). A strong correlation of ANP increment was observed with CK-MB (r2 = 0.908) and LDH (r2 = 0.911) activity. No significant correlation was observed between ANP and glucose levels with chronic HH.
Chronic HH-induced RVH is associated with apoptosis in right heart
The chronic HH-induced pathological damages were analyzed in rats with assessment of apoptosis in cardiac tissues. It was found that HH induced apoptosis in rat hearts by 14 days by cytochrome-c release and caspase-3 activation (Fig. 6a–d). This was further confirmed by appearance of TUNEL-positive nuclei in hearts of animals exposed to HH for 21 days (Fig. 6e, f). These findings suggest that caspase activation occurs before DNA damage under HH, and prolonged HH stress activates nuclear signaling for apoptosis promoting DNA damage. The data clearly demonstrate that sustained deprivation of oxygen induces potentially pathological damage to the myocardium by initiating myocardial programmed cell death.
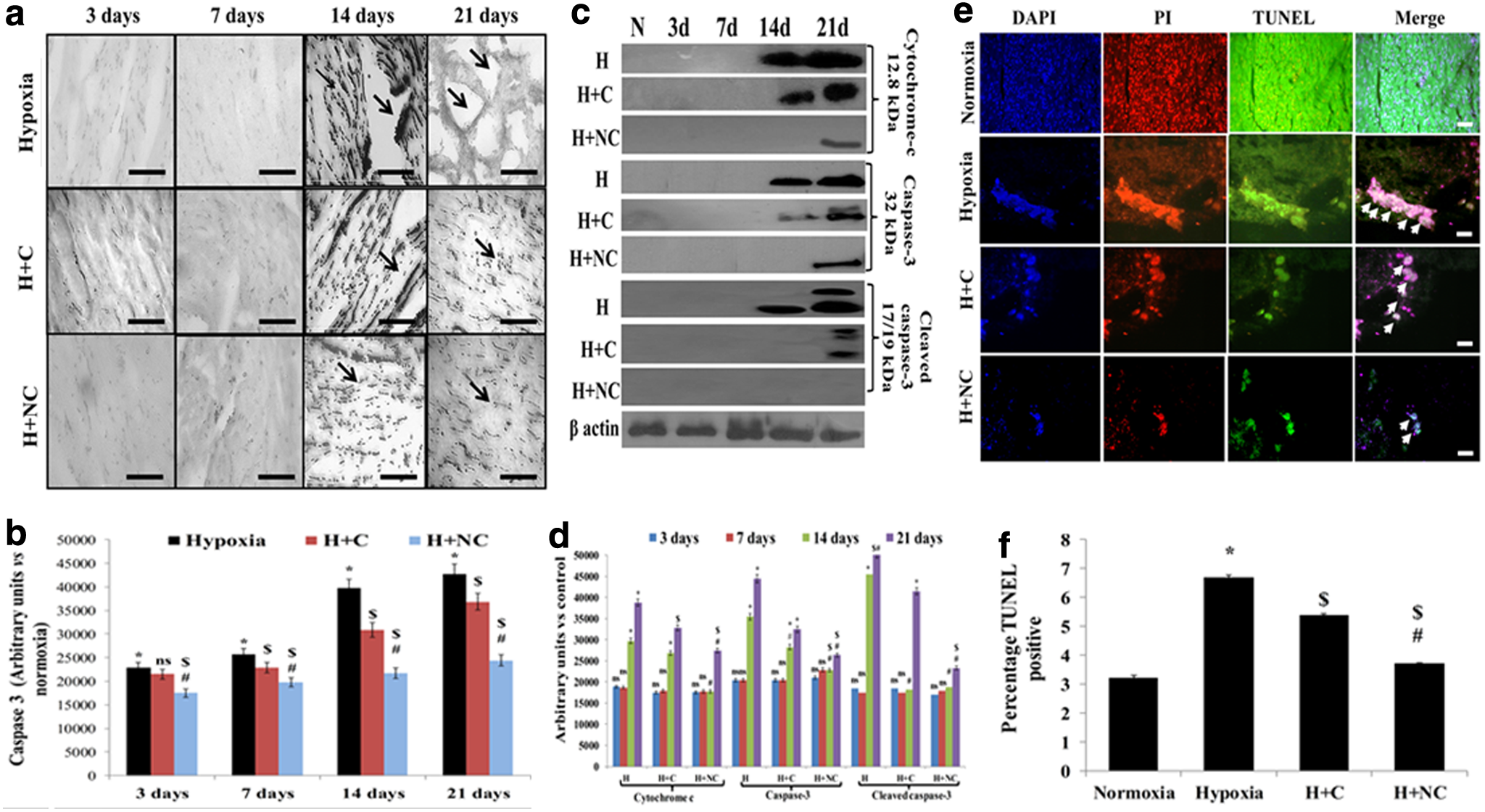
Figure showing tissue expression of caspase-3
Importantly, although both nanocurcumin and curcumin supplementations reduced apoptotic signaling in rat hearts, the effects were more pronounced in nanocurcumin-supplemented animals clearly depicting the improvement in cardioprotective efficacy of nanocurcumin by reducing programmed cell death.
Oxidative balance and cytokine response under HH
Oxidative damage remains an important process, which further promotes pathological damage in the myocardium. In addition, cytokine responses play a major role in transition from adaptive hypertrophy to dilation and RV dysfunction (Sano et al., 2000; Sriramula et al., 2008). We assessed tissue ROS leakage and lipid peroxidation as markers of oxidative damage in animals with HH. We also estimated circulating levels of TNFα, IL-6, and VEGF to assess immunological response in animals under HH (Supplementary Table S3).
It was found that excessive ROS leakage (Fig. 7a) and lipid peroxidation (Fig. 7b) were increased in animals exposed to HH within 3 days of exposure (p ≤ 0.05) compared to normoxia. The circulating levels of TNFα and IL-6 (p ≤ 0.05) increased up to 7 days of HH compared to normoxia, beyond which no significant change was observed. Nanocurcumin treatment significantly reduced oxidative damage (p ≤ 0.05) and lowered cytokine levels (p ≤ 0.05) more than curcumin at all time points during HH and in comparison to vehicle-treated animals.

Graphical representation of changes in ROS formation
Discussion
An excessive work load during chronic HH results in strain on the right heart, which causes discrete morphological, functional, and molecular changes of RVH that in the short term can be considered adaptive responses (Hart, 2003; Hainsworth and Drinkhill, 2007). However, prolonged stress eventually promote right heart dilation and gradually transition toward disease progression and decompensation. This decompensatory RVH appears to be an initial signaling event, which might further evoke more serious clinical deterioration (Diwan and Dorn, 2007). Studies have shown that apoptotic cell death appears as a pathophysiological mechanism underlying many disorders that affect the heart (Kang et al., 2004; Foo et al., 2005). In the present study, we demonstrated that chronic HH-induced RVH in adult male Sprague-Dawley rats was accompanied with apoptosis in the right heart as early as 14 days of exposure along with sustained redox and metabolic imbalance, in addition to an inflammatory cytokine response. We also assessed the improvement in therapeutic potential of nanotized curcumin, that is, nanocurcumin, in ameliorating HH-induced RVH and apoptosis and compared it to curcumin.
In the present study, increased levels of ANP were observed within 3 days of HH along with collagen accumulation and an increase in RV/BW and RV/LV+S, indicative of RVH. Corroborating this, slight increases were observed in BP and HR of the animals (7 days), which normalized after prolonged exposure (14 days), but the results were not significant. These data indicate that the in vivo experimental model of RVH developed in the present study was consistent with the previous findings suggesting induction of RVH without the development of systemic hypertension (Sharma et al., 2004).
ANP remains a key marker of hypertrophy in cardiomyocytes. ANP binds to cellular receptors GC-A present on the outer surface of plasma membrane and promotes cytosolic accumulation of cGMP and activation of downstream signaling cascades. An important secondary messenger and negative regulator of hypertrophy, cGMP also modulates vascular tone to improve contractibility under stress by regulating intracellular calcium levels. Importantly, the cGMP and cGK-1 levels initially remained stable and decreased with subsequent increase in HH exposure (21 days), but the levels of ANP continued to increase. These comprehensive findings indicate that HH-induced RVH is regulated by ANP and cGMP levels in a time-dependent manner. While acute HH did not affect cGMP levels, a chronic HH-induced decrease in cGMP may promote RVH with gradual increase in ANP levels. These data collectively suggest that acute exposure to HH initiates compensatory RVH in animals, whereas chronic HH causes a decompensatory transition, at least in part by modulating ANP-cGMP levels.
Since cardiomyocytes are considered terminally differentiated cells, induction of apoptosis may cause nonreversible cardiac damage, which can be lethal, if untreated (Soonpaa and Field, 1998). Nanocurcumin supplementation clearly reduced apoptosis as evident by the decreases in tissue expression of cytochrome-c and caspase-3 along with TUNEL-positive nuclei with HH. These findings were closely associated with reduction in ROS leakage and maintenance of normal lipid peroxidation in the hearts of nanocurcumin-supplemented animals compared to curcumin. Collectively, these findings suggests that cellular apoptotic events are inevitably associated with free radical damage with HH and play a critical role in largely irreversible pathological damage to cardiomyocytes. We have recently showed the protective efficacy of nanocurcumin under hypoxia in vitro (Nehra et al., 2015a, 2015b). The findings in the present extend these findings to rodent hearts during chronic HH. Importantly, curcumin supplementation had considerably less protective efficacy than nanocurcumin, suggesting that nanotization of curcumin significantly improves the pharmacological properties of curcumin.
Importantly, CaMkinase II expression levels increased in right hearts along with the total intracellular calcium pool. Our previous studies have shown that intracellular calcium regulation is crucial for maintaining normal physiological function in H9c2 cardiomyoblasts during hypoxia (Nehra et al., 2014). The data in present findings further demonstrate that rodent cardiac muscles experience calcium imbalance under chronic hypoxic stress. Knowing that maintenance of intracellular calcium is critical for normal contractile function, the data suggest that chronic HH might evoke hypercontractibility in the right heart initially, but eventually leads to decompensation and pathological damage. However, further investigations dedicated toward quantitative assessment of cardiac muscle contractibility need to be done. Taken together, the calcium imbalance and induction of apoptosis in right heart strongly indicate that although RVH is initiated as an adaptive response to combat increased work load during HH, prolonged stress induces pathological damage and apoptosis in the right heart.
We assessed the improvement in the efficacy of nanocurcumin in ameliorating HH-induced cardiac damage compared to curcumin. We found that nanocurcumin supplementation decreased ANP levels in animals exposed to HH and restored cGMP levels. This was corroborated with reduced RV/LV+S and RV/BW ratios, collagen accumulation, and modulations in tissue architecture. In addition, BP and sHR, along with total cardiac intracellular calcium pool and CaMkinase II levels, were effectively preserved in animals supplemented with nanocurcumin compared to curcumin during chronic HH. The data strongly suggest that nanocurcumin treatment protects the right heart from chronic HH-induced damage by regulating ANP/cGMP levels better than curcumin in rats, thus illustrating a therapeutic potential of curcumin.
Oxidative stress has an important impact on cardiomyocyte apoptosis. Studies have previously reported the substantial role of redox damage in disease progression (Nakanishi et al., 1995; Sorescu and Griendling, 2002; Valko et al., 2007). To check whether HH-induced RVH was associated with redox damage, we quantified the excessive ROS leakage and lipid peroxidation levels in the RV. We found that ROS leakage increased in the RVs as early as 3 days of HH accompanied with lipid peroxidation. These data support the findings of an increase in caspase activation along with TUNEL positivity confirming that HH-induced oxidative stress might be the initiating signal for pathological changes in the right heart.
We observed an increase in plasma LDH activity, suggesting a decline in metabolic adaptation during chronic HH. This finding encouraged us to assess whether the cardiac muscles specifically underwent metabolic insufficiency and injury. We assessed CK-MB activity as a marker of cardiac metabolic efficiency and injury in rat hearts under chronic HH. Our previous report suggested that hypertrophied cardiomyocytes experience a switch of preferred metabolic substrates from fatty acids to glucose along with severe metabolic damage (Nehra et al., 2015a). In the present study, we observed an increase in plasma CK-MB clearly demonstrating metabolic damage and injury to cardiac muscles with hypoxia. These data accord with previous studies demonstrating the critical relevance of CK-MB as a biomarker for metabolic damage and cardiac injury (Ye et al., 2001; Brancaccio et al., 2010). However, the underlying molecular mechanisms behind these maladaptive metabolic changes under chronic HH remain unknown.
Activation of inflammatory cytokines is a critical pathophysiological response, which has been shown to further promote myocardial pathological damages under stress (Nian et al., 2004). Stress-induced activation of cytokines such as TNFα and IL-6 is well known to have crucial effects on cardiac function (Damås et al., 2001). In the present study, increments in circulating TNFα (Feldman et al., 2000) and IL-6 (Sano et al., 2000) levels during chronic HH illustrated induction of immune response under hypoxic stress, which further promoted pathological effects along with RVH. Interestingly, nanocurcumin supplementation effectively reduced TNF and IL-6 in animals exposed to HH compared to curcumin- and vehicle-treated rats. Importantly, and in contrast, curcumin treatment did not have any effect in restoring the cytokine activation in animals during hypoxia. Collectively, the data indicate that nanotization of curcumin to nanocurcumin effectively combats damaging responses during HH and protects the heart better than curcumin.
Limitations of the Study
Since direct invasive/noninvasive measurements to analyze changes in pulmonary artery pressure were not performed, the direct implications of hemodynamic changes on RV function and remodeling could not be assessed in the present study. Another limitation is the absence of measurements of the extent of muscularization of small pulmonary vessels. However, we show indirect evidence of reduced RV function with chronic hypoxic stress in rats in terms of apoptotic cell death and redox damage.
Footnotes
Acknowledgments
The present work was funded by Defence Research and Development Organization (DRDO), Government of India. Sarita Nehra is a recipient of University Grants Commission fellowship. Varun Bhardwaj is a recipient of Council for Scientific and Industrial Research fellowship. The authors sincerely thank Director, DIPAS, for providing necessary facility to carry out the research work. The authors also thank Mr. Bhagwat Singh (Experimental Animal Facility, DIPAS) for providing necessary help in carrying out animal experiments.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.