
Abstract
Abstract
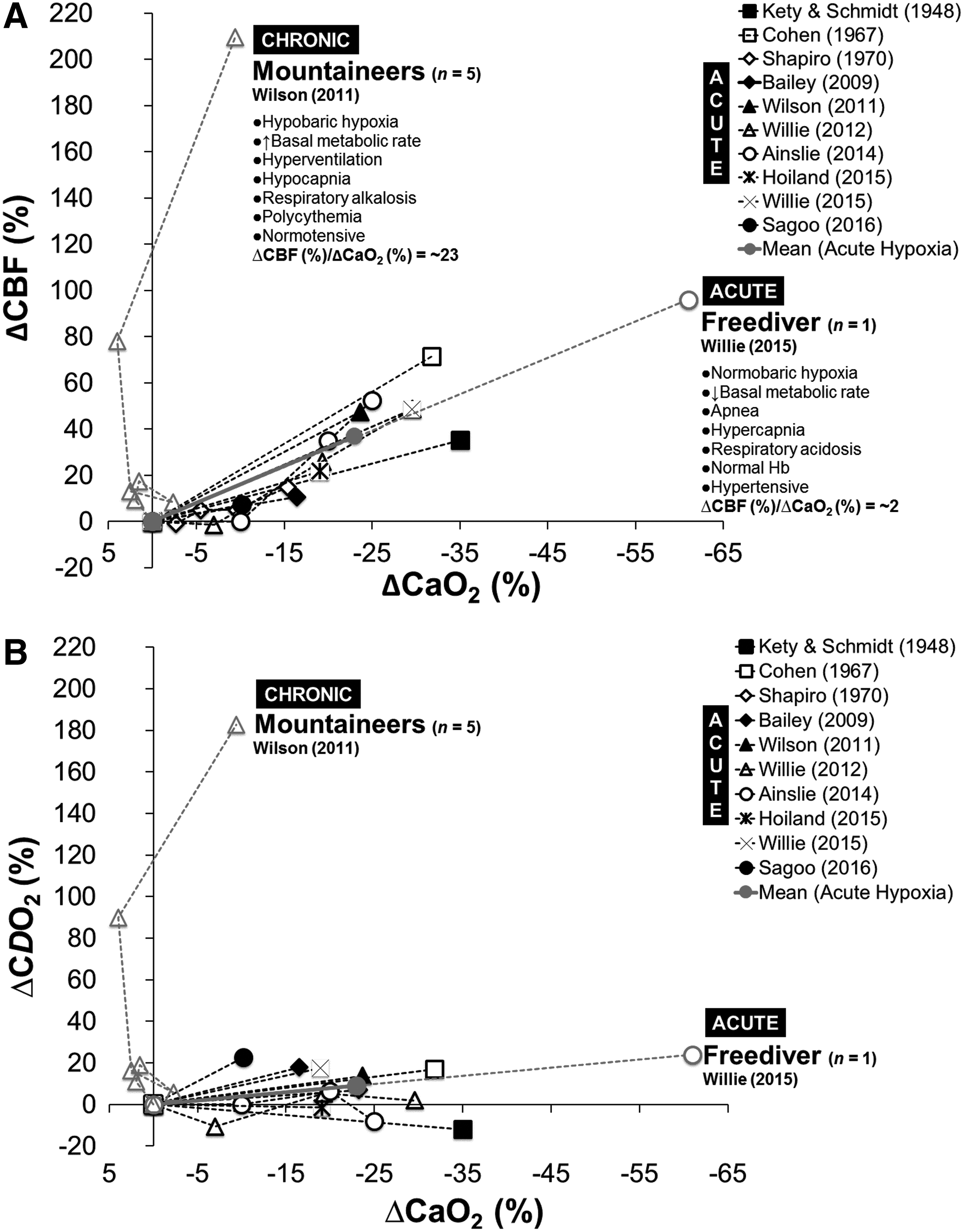
Bailey, Damian M., Christopher K. Willie, Ryan L. Hoiland, Anthony R. Bain, David B. MacLeod, Maria A. Santoro, Daniel K. DeMasi, Andrea Andrijanic, Tanja Mijacika, Otto F. Barak, Zeljko Dujic, and Philip N. Ainslie. Surviving without oxygen: how low can the human brain go? High Alt Med Biol 18:73–79, 2017.—Hypoxic cerebral vasodilation is a highly conserved physiological response coupling cerebral O2 delivery (CDO2) to metabolic demand with increasingly important roles identified for the red blood cell (sensor) and nitric oxide (effector). In the current article, we reexamine previously published cerebral blood flow (CBF) and arterial blood gas data obtained in freedivers and mountaineers, extreme athletes in whom the lowest arterial partial pressures of O2 (19–23 mmHg) and greatest extremes of carbon dioxide (16–61 mmHg) were recorded during (acute) maximal static dry apnea or (chronic) exposure to terrestrial high altitude. Data highlight compensatory increases in CBF (+96% in freedivers to +209% in mountaineers relative to normoxic baseline controls) that were sufficient to sustain CDO2 (+24% in freedivers to +183% in mountaineers) even in the face of the most severe reductions in arterial O2 content (−61% in freedivers to −9% in mountaineers) reported in the literature, consistent with the conservation of mass principle. These unique findings highlight to what extent cerebral vasodilation likely contributes toward these athletes' extraordinary abilities to survive in such harsh environments characterized by physiological extremes of hypoxemia, alkalosis, and acidosis helping define the human brain's remarkable limits of tolerance.
Introduction
Paleoclimate and evolution of the human brain
E
However, this obligatory high rate of O2 consumption is associated with high “vulnerability for failure” given the brain's paradoxically limited O2 reserves. Indeed, assuming an average brain tissue partial pressure of O2 (PO2) of ∼25 mmHg and lack of O2-binding proteins, its O2 content is a meagre ∼30 nmol/mL such that given an average cerebral metabolic rate of oxygen (CMRO2) of 30 nmol/mL/s, the O2 present would sustain metabolism for at best 1 second if blood supply were to be interrupted by anoxia (Leithner and Royl, 2014). Unable to compromise on its excessive energy budget, failure of ATP-dependent ion exchangers results in the breakdown of ionic gradients and membrane depolarization triggering a cytotoxic increase in intracellular Ca2+ concentration and uncontrolled release of excitatory neurotransmitters that ultimately converge in neuronal death (Lipton, 1999).
Yet, O2 is not the only atmospheric gas to have shaped life on Earth with an equally important role identified for carbon dioxide (CO2), the key substrate for oxygenic photosynthesis (Cummins et al., 2014). The human brain has since evolved heightened sensitivity to PaCO2/H+ (more so than PaO2) that extends throughout the cerebrovasculature, from the large extracranial and intracranial conduit and middle cerebral arteries through to the smallest pial arterioles and parenchymal vessels, prioritizing the buffering of brain tissue pH for stabilization of chemosensory and autonomic control at the level of the brainstem (Kety and Schmidt, 1948a; Willie et al., 2014). Given that the brain's O2-CO2 balance is so delicate, it would therefore seem intuitive for evolution to favor defense mechanisms capable of sensing “crosstalk” between these respiratory gases and orchestrating the coordinated transmission of vasoactive signals to the cerebrovasculature coupling local cerebral O2 delivery (CDO2) to tissue metabolic demand.
In the current article, we reexamine previously published cerebral blood flow (CBF) and arterial blood gas data obtained in freedivers and mountaineers, extreme athletes in whom the lowest PaO2's and greatest extremes of PaCO2 (from respiratory alkalosis in the mountaineer to acidosis in the freediver) to date have been documented in the literature. These unique datasets reveal that even at physiological, if not indeed pathological extremes of systemic hypoxemia and hypo/hypercapnia, adaptive increases in CBF and corresponding CDO2 are sufficient to compensate for any potential reductions in arterial O2 content (CaO2), such that cerebral homeostasis is preserved consistent with the conservation of mass principle.
Materials and Methods
Landmark performances
History reveals to what extent human tolerance to physiological extremes of hypoxia, alkalosis, and acidosis has traditionally been underestimated by the scientific community. The mountaineer Edward Norton got to within 300 m of the summit of Mt. Everest in 1924 without supplementary O2 and numerous failed attempts in the years that followed, including laboratory simulations led the physiological community to predict that an “oxygenless” ascent would prove physiologically impossible (West, 2009). However, the remaining 3% was successfully negotiated by Reinhold Messner and Peter Habeler, and the world's highest mountain finally conquered, albeit some 54 years later. Medical opinion was equally divided when the freedivers, Jacques Mayol and Enzo Maiorca, attempted to dive below 50 m without any breathing apparatus in the late 1960s. While both survived the challenge and later went on to exceed 100 m, precisely how modern-day world record holders, such as Herbert Nitsch (who descended to 214 m attached to a metal sled and fixed line during the “No Limits” discipline) and Stéphane Mifsud (who achieved an 11 minutes 35 seconds breath-hold with the respiratory tract immersed in water during the “Static Apnea” discipline), achieve such remarkable performances remains, at best enigmatic.
Landmark studies
We reexamined published data from three unique studies detailed below:
Mountaineers
Arterial samples were obtained from the femoral artery of four acclimatized mountaineers at an altitude of 8400 m on Mt. Everest following descent from the summit having spent 30 minutes breathing ambient air preceded by supplementary O2 (2–3 L/min while climbing, 0.5 L/min while sleeping above 7100 m) (Grocott et al., 2009). Individual data were available for all mountaineers in the published article. However, given that it was not possible to measure CBF at this elevation due to adverse weather, we interrogated a separate publication by the same group in a subset of five mountaineers (group not individual data available) incorporating transcranial doppler ultrasound of the right cerebral middle artery performed at the lower elevation of 7950 m, 30 minutes following removal of supplementary O2 (Wilson et al., 2011). These are the highest elevations where such measurements have been performed.
Freedivers
Fifteen elite freedivers (seven having placed top 10 in the world) were examined during a static dry (laboratory-based) apnea. Individual data were made available for all freedivers (Dr. C.K. Willie, personal communication). Volumetric flow was measured continuously using near-concurrent duplex vascular ultrasound measures of blood flow through the right internal carotid and left vertebral arteries complemented by serial samples obtained from the radial artery for blood gas analysis (Willie et al., 2015a).
Results and Discussion
Respiratory gas extremes and cerebral vasodilation: significance and mechanisms
A comparison of arterial blood gas results is outlined in Table 1, including an as of yet unpublished presentation of the most hypoxemic individuals in whom the lowest PaO2's (19–23 mmHg) and greatest extremes of PaCO2 (16–61 mmHg) were recorded. Despite exhibiting the lowest PaO2 and SaO2, polycythemia attenuated the reduction in CaO2 observed in the mountaineer such that the freediver was ultimately the most hypoxemic. Given that supplemental O2 is prescribed to patients when PaO2 falls <60 mmHg (consistent with the clinical diagnosis of respiratory failure) and risk of death increases exponentially <25 mmHg, combined with the increased mortality observed in hospitalized patients with PaCO2's of <35 or >45 mmHg (Laserna et al., 2012), puts their extraordinary tolerance to hitherto unchartered extremes of hypoxemia, alkalosis, and acidosis into clearer physiological perspective.
Freediver and mountaineer data based on publications of Willie et al. (2015a) and Grocott et al. (2009), respectively. Group data expressed as mean ± SD; PaO2/PaCO2, arterial partial pressure of oxygen/carbon dioxide; SaO2, arterial oxyhemoglobin saturation; Hb, hemoglobin.
Dr. C.K. Willie (personal communication).
Includes male and female values.
Different (p < 0.05) between disciplines assessed by independent samples t-tests following confirmation of distribution normality (Shapiro–Wilk W tests).
Calculated from raw data: TCO2, total carbon dioxide = [HCO3−] + αPCO2, where α = 0.226 mmol/L/kPa (kPa = mmHg ×
These unique datasets provide unique insight into the underlying mechanisms, notably the extent to which cerebral vasodilation likely promotes survival in such harsh environmental extremes. As illustrated in Figure 1A, this highly conserved physiological response compensates for any reduction in CaO2 even at the physiological extremes of arterial hypoxemia highlighting the brain's ability to defend against any potential threat to oxygenation. We observe a doubling of CBF in the freediver such that CDO2 (Fig. 1B) always remains preserved especially considering the reduction in CMRO2 recently documented in freedivers near apnea breakpoint, a consequence of the remarkable hypercapnia endured by these athletes (Bain et al., 2016). Thus, the combined benefits of cerebral hypometabolism and elevated capillary to mitochondrial O2 diffusive conductance, the primary resistance to vascular O2 transport (Wagner, 1996), likely optimize O2 demand–supply coupling to protect the brain from severe hypoxemia, part of a coordinated defense strategy commonly observed in (more) hypoxia-tolerant ectothermic species and diving mammals (Lutz et al., 2003; Bailey et al., 2009a).
Relative increases in CBF
Emerging evidence supports a key regulatory role for nitric oxide (NO) as a downstream signaling molecule with the capacity to control and coordinate hypoxic vasodilation, catalyzed by the allosteric transition of Hb from the relaxed (oxy) to tense (deoxy) state with separate roles for the stable metabolites, nitrite, and S-nitrosoHb widely contested given their ability to conserve and transfer bioactivity within the microcirculation (Stamler et al., 1997; Cosby et al., 2003). The emerging picture, at least in hypoxia-tolerant vertebrates, including humans indigenous to terrestrial high altitude (native highlanders), suggests that an adaptive increase in vascular and tissue NO bioavailability serves to reduce CMRO2, enhance cytoprotection against oxidative damage, and improve coupling of flow and metabolism (Umbrello et al., 2013; Fago and Jensen, 2015). In further support, dietary nitrate supplementation has been shown to exert an O2-conserving effect, elevate SaO2, and improve maximal apnea performance (Patrician and Schagatay, 2016). This contrasts with the opposing, generally maladaptive responses typically observed in (less hypoxia tolerant) native lowlanders during prolonged exposure to high altitude that include elevations in basal metabolic rate (BMR), free radical-mediated scavenging of NO, and vascular endothelial impairment (Bailey et al., 2010).
Atmosphere versus biosphere: battle of the gases
While both athletes share the common feature of severe arterial hypoxemia, the environmental challenges to which they are exposed and corresponding physiological demands are markedly different (Fig. 1A) suggesting that the mechanisms by which CDO2 is sustained may differ between groups. Cerebral vasodilation depends on the magnitude and duration of the hypoxic stimulus, extent of acid–base adjustment, intrinsic cerebral reactivity to changes in O2, CO2, pH, and systemic/local release of vasoactive metabolites (Ainslie et al., 2016). High-altitude mountaineering is characterized by long-term exposure to hypobaric hypoxia, increased BMR, hyperventilation-induced hypocapnia, respiratory alkalosis, normotension, and polycythemia, whereas freediving is characterized by short-term exposure to normobaric hypoxia during apnea, decreased BMR, hypercapnia, respiratory acidosis, and hypertension subsequent to chemoreflex activation of the sympathetic nervous system, with normal Hb concentrations. Freedivers also have to contend with the additional challenge of elevated hydrostatic pressure when competing “at depth” in select disciplines (e.g., 22.3 ATA during Herbert Nitsch's No Limits world record) and complications associated with pulmonary barotrauma, nitrogen narcosis, decompression sickness, and high pressure neurological syndrome (Lindholm and Lundgren, 2009). Indeed, it is intriguing to note that the lowest PO2's recorded in humans can arise from opposing ends of the atmospheric–biospheric pressure spectrum.
The overall CBF response to high altitude reflects the net balance of two opposing physiological stimuli, hypoxic vasodilation versus hyperventilation-induced hypocapnic vasoconstriction. Despite the brain's increased sensitivity to PaCO2 (Kety and Schmidt, 1948a; Willie et al., 2012), the balance is tipped in favor of hypoxic vasodilation during prolonged exposure to extreme altitude reflecting the overarching drive to preserve CDO2 as illustrated in Figure 1A, B. Indeed, it is remarkable that extreme hypoxemia in the mountaineers evoked an order of magnitude more vasodilation (technical differences acknowledged) compared to the synergism expected between extreme hypoxemia, hypercapnia, and hypertension in the freediver. Indeed, it is of interest to note that despite marked differences in the environmental challenges encountered and varying PaCO2's, the slope of the ΔCBF-CaO2 relationship for the single (most hypoxemic) freediver is identical to the average slope observed for the mountaineers during acute hypoxia.
The observed differences in acid–base balance are likely to exert opposing effects on the athletes' respective O2-Hb dissociation curves given the inverse relationship between O2-Hb binding affinity, pH, and pCO2 (Bohr effect) (Bohr et al., 1904). Respiratory alkalosis (mountaineer) will cause a leftward (increased O2-Hb affinity) facilitating greater O2 uptake in the lung for any given alveolar PO2, although this is likely to be countered at least to some extent by 2,3 diphosphoglycerate formation (Mairbaurl et al., 1993). In contrast, respiratory acidosis (freediver) will cause a rightward shift (decreased O2-Hb affinity) favoring O2 off-loading to the tissues. Evidence generally favors a leftward shift as being the most beneficial during severe hypoxia (Petschow et al., 1977) given that the slope of the O2-Hb dissociation curve at the prevailing alveolar PO2's is steeper than the slope at the tissue PO2's (Turek et al., 1984) rendering Nitsch's freediving record even more impressive. Furthermore, respiratory alkalosis blunts sympathetic tone, pulmonary vasoconstriction, and cerebral vasodilation, whereas the opposite occurs in response to hypercapnic acidosis further highlighting the complex relationship between acid–base balance and physiological responses to hypoxia (Swenson, 2016).
Pushing the limits of hypoxemia and hyperemia
Since the mid to late 1940s, the No-Limits freediving world record has improved by a remarkable 720% (from 30 to 216 m) compared with other sports, for example, a 6% improvement in the 100 m sprint world record observed over a similar time frame (from 10.20 to 9.58 seconds). The question thus arises as to what is the maximum depth or lowest PaO2 that the healthy human can ultimately tolerate before loss of consciousness (LOC) ensues, herein referred to as the critical PaO2 (PaO2CRIT)? Pioneering studies reported PaO2CRIT values in the range of 16–20 mmHg during instantaneous hypoxia induced by rapid decompression (1 second) or nitrogen hyperventilation (17 seconds) (Ernsting, 1963). While such challenges are clearly supraphysiological, LOC generally supervenes when the jugular venous (jv) PO2 falls to ∼20 mmHg (range 17–20 mmHg), which corresponds to an approximate SvO2 (upper limit) of 32% (Severinghaus, 1966) and calculated CvO2 of 6.8/100 mL (assuming a Hb concentration of 15 g/dL). From these figures, it is possible to approximate the PaO2CRIT assuming a global (average) CBF of 46 mL/100 g/min (36–56 mL/100 g/min) equivalent to 644 mL/min (504–784 mL/min) assuming an average brain mass of 1400 g (1300–1500 g) and average CMRO2 of 3.0 mL/100 g/min (2.6–3.4 mL/100 g/min) or 42 mL/min (36–48 mL/100 g/min) (Madsen et al., 1993). Using the average values cited, the CaO2-CvO2 can thus be derived using the Fick Principle:
This equates to an (uncompensated) CaO2 of 13.3/100 mL (6.8 + 6.5/100 mL) as the average minimum to avoid LOC or SaO2CRIT of ∼65% and PaO2CRIT of ∼35 mmHg, values that are far in excess of those recorded in the two most hypoxemic athletes documented in Table 1, suggesting that other compensatory factors, notably cerebral vasodilation (and acid–base adjustments) are contributing. In support, recalculation of the Fick Principle demonstrates that the doubling of CBF observed in the freediver has the effect of further lowering SaO2CRIT to ∼40% and PaO2CRIT to ∼23 mmHg (latter corrected for prevailing pH and base excess), almost identical to the values observed (Table 1). These values are likely to be even lower for the mountaineers given such pronounced hyperemia, although unlikely to fall below 20 mmHg, which appears to be the lower limit observed in patients with severe respiratory disease who also exhibit polycythemia and maximal cerebral vasodilation (Refsum, 1963).
Whether prolonged exposure to extreme hypoxia in the Wilson et al. (2011) study represents maximal dilation of the cerebral vessels is unknown. It would be interesting to examine if experimental manipulation of PaCO2 would incur further increases given the ∼5.5% increase in CBF per mmHg increase in PaCO2 reported during iso-oxic end-tidal forcing (Willie et al., 2012). However, the CBF response at physiological extremes of hypoxia–hypercapnea is not a simple summative dose–response. Emerging evidence suggests that severe hypoxia blunts the normal cerebrovascular reactivity to CO2 possibly due to exhaustion of the cerebral vasodilatory reserve (Fan and Kayser, 2013) or alternatively, to protect against severe blood–brain barrier disruption and complications associated with extracellular vasogenic cerebral edema (Bailey et al., 2009c).
Notwithstanding the uncertainties associated with acid–base adjustments and corresponding shifts in the O2-Hb dissociation curve as outlined, these calculations are at the very least hypothesis testing and demonstrate that these athletes are operating very close to, if not indeed at, the very limit of human consciousness, findings corroborated by the high incidence of LOC and loss of motor control (confusion, loss of postural control, faltering vision, speech problems) recorded among competitive freedivers, clinical signs that are the frustrating cause for disqualification. Furthermore, emerging MRI and immunochemical evidence suggests that extremes of freediving and high-altitude mountaineering may be associated with structural brain damage (Bailey et al., 2009a; Gren et al., 2016) with potential long-term neuropsychological consequences. This contrasts with the extraordinary hypoxia tolerance exhibited by diving mammals, including the seal that is capable of enduring PaO2's as low as 7–10 mmHg without cerebral integrity being compromised (Elsner et al., 1970).
While global CDO2 in humans may appear well preserved both at rest and even during maximal exercise in hypoxia, the overall reduction in jv (Ainslie et al., 2014) and (derived) cerebral mitochondrial PO2 (Bailey et al., 2011) reported in the literature suggests that it is heterogeneous and regional PO2 deficits likely coexist, which has important implications for regional susceptibility to damage or indeed adaptation. Regional differences in the CBF, and hence CDO2 response to hypoxia, have since been observed, complicated in part by differing analytical techniques, experimental designs, and fact that the entire cerebral arterial tree including large arteries of the neck exhibit differential reactivities to PaO2 and PaCO2 (Willie et al., 2014). Increased CBF reactivity has been reported in the putamen, brain stem, thalamus, caudate nucleus, nucleus accumbens, and pallidum using H215O positron emission tomography (Binks et al., 2008), whereas single photon emission computed tomography has identified predilection for the anterior cingulate cortex, right temporal lobe, sensory motor cortices, prefrontal cortex, and basal ganglia (Pagani et al., 2011). Collectively, these findings have been taken to reflect the selective preservation of CDO2 to phylogenetically older regions of the brain responsible for more vital aspects of homeostatic function, a feature that may have provided a selective advantage.
Conclusion
A reexamination of published data from freedivers and mountaineers, extreme athletes in whom the lowest PaO2's and greatest extremes of PaCO2 to date have been recorded, highlights to what extent compensatory increases in CBF are sufficient to preserve CDO2 consistent with the conservation of mass principle. While the molecular mechanisms underlying O2-CO2 sensing remain elusive, cerebral vasodilation likely contributes toward these athletes' extraordinary abilities to survive in such harsh environments characterized by physiological extremes of hypoxemia, alkalosis, and acidosis helping define the human brain's remarkable limits of tolerance.
Footnotes
Acknowledgments
Aspects of this work were supported by the Higher Education Funding Council for Wales (D.M.B.), Canada NSERC Discovery Grant (P.N.A.), and FP7-PEOPLE-2010-ITN-264816 PHYPODE Grant (Z.D.). D.M.B. is the Reichwald Family UBC Southern Medical Program Chair in Preventive Medicine, C.K.W. is a Vanier Canada graduate scholar, and P.N.A. is a Canada Research Chair in Cerebrovascular Physiology.
Authors' Contributions
D.M.B. drafted the article and revisions thereof; co-authors approved the final article for submission.
Author Disclosure Statement
No competing financial interests exist.