
Abstract
Abstract
Counter, S. Allen, Leo H. Buchanan, Fernando Ortega, Anthony B. Jacobs, and Göran Laurell. Assessment of the brainstem-mediated stapedius muscle reflex in Andean children living at high altitudes. High Alt Med Biol. 18:37–45, 2017.—This study examined the physiological thresholds, amplitude growth, and contraction duration of the acoustic stapedius reflex (ASR) in Andean children aged 2–17 years living at altitudes of 2850 m (Altitude I Group) and 3973 m (Altitude II Group) as part of a general medical assessment of the health status of the children. The brainstem-mediated ASR reveals the integrity of the neuronal components of the auditory reflex arc, including the cochlea receptors, eight cranial nerves, and brainstem neural projections to the cochlear nuclei, bilateral superior olivary nuclei, facial nerve nuclei, and facial nerve and its stapedius branch. Uncrossed (ipsilateral) and crossed (contralateral) ASR thresholds (ASRT), ASR amplitude growth (ASRG) function, and ASR muscle contraction duration (decay/fatigue) (ASRD) were measured noninvasively with 500, 1000 Hz and broadband (bandwidth = 125–4000 Hz) noise stimulus activators using a middle ear immittance system. Oxygen saturation (SaO2) level and heart rate were measured in a subsample of the study group. Statistical analyses revealed that the Altitude I and Altitude II groups had ASRT, ASRG function, and ASRD rates comparable to children at sea level and that the two groups were not significantly different for any of the ASR measures. No significant association was found between SaO2 or heart rate and ASRT, growth, and muscle fatigue rate. In conclusion, the assessment of the ASR in children in the high-altitude groups revealed normal function. Furthermore, the results indicate no adverse oto-physiological effects of altitude on the brainstem-mediated ASR at elevations between 2850 and 4000 m and suggest normal middle ear and auditory brainstem function.
Introduction
O
The middle ear contains two small skeletal muscles, the tensor tympani and the stapedius. In humans, the tensor tympani responds reflexively to tactile stimuli, whereas the stapedius muscle responds reflexively to moderate and higher intensity level acoustic stimuli, which is the basis for clinical acoustic stapedius reflex (ASR) testing. The brainstem-mediated ASR provides noninvasive clinical diagnostic information on the status of the tympanic membrane, middle ear, cochlea, eight cranial nerves, and bilateral brainstem neural projections to the cochlear nuclei, superior olivary complex, facial nerve nuclei, seventh cranial nerve and its stapedius nerve branch, and the contractile mechanisms of the stapedius skeletal muscle (Borg, 1973; Borg and Counter, 1989).
A number of otological and neurological conditions may affect the functioning of the ASR, including tympanic membrane dysfunction, middle ear pathology, cochlear impairment, retrocochlear pathology, hypothyroidism, and neuromuscular diseases, such as, Bell's palsy, myasthenia gravis, multiple sclerosis, amyotrophic lateral sclerosis, and myotonic dystrophy type 1. In middle ear disorders and cochlea impairment, the ASR characteristically shows elevated or absent ASR thresholds (ASRT) and abnormal diminution in the ASR amplitude. In the oto-neurological condition of Bell's palsy, for example, the ASR may be absent or hypersensitive. In eighth nerve tumors and multiple sclerosis, the ASR typically shows a diminished amplitude of the uncrossed (ipsilateral) and crossed (contralateral) brainstem physiological responses and a rapid fatigue or adaptation of the stapedius muscle contraction from its maximum onset amplitude. In neuromuscular diseases, such as the autoimmune disorder myasthenia gravis, the ASR may reveal weak and reduced contractions as a result of circulating antibodies that block acetylcholine receptors at the postsynaptic neuromuscular junction in the stapedius muscle (Mattle et al., 1986; Smith and Brezinova, 1991; Shimizu et al., 1992; Ardiç et al., 1997; Goulis et al., 1998; Tóth et al., 2000; Osanai et al., 2001; Dursun, 2008; Cingi et al., 2010; Counter et al., 2011; Counter et al., 2012). These studies indicate that the ASR is a useful diagnostic tool for the assessment of auditory system and neurological pathophysiology.
Investigations of the effects of altitude on ASR function have been limited. Laurikainen et al. (1990) used experimentally-induced hypoxic hypoxia conditions that mimic high altitude physiological states to assess the effects of altitude on the ASR. These investigators found that hypoxia modulates the amplitude of the ASR, causing apparent reductions in the velocity and amplitude of the ASR associated with oxygen saturation (SaO2) levels.
While persons living at high altitudes have adapted to low oxygen conditions, otological disorders may still be a cause for concern, particularly in children. The ASR procedure was used in this study as part of a clinical battery for objective evaluation of the otological status of the middle ear, inner ear, and auditory brainstem function in children living at high altitudes in the Andes Mountains. The specific purpose of this study was to investigate the physiological integrity of the ASR in children living at high altitudes in the Andes Mountains of Ecuador.
Materials and Methods
Participants and location
This study consisted of 97 children (54 females and 43 males) aged 2–17 years (mean age: 9.9; standard deviation [SD]: 3.4; median age: 10 years) that comprised two groups living at high elevations in La Victoria at 2850 m and in the Guangaje Andean villages of Curingue and Tingo Pucara at 3973 m (3500–4200 m) in Ecuador, South America. All children were examined using the ASR as part of an audiological clinical battery during general medical examinations. The children included in this study had normal hearing and normal middle ear functioning, as assessed by pure-tone hearing screening and tympanometry, respectively. Any participant exhibiting hearing impairment was not included in the ASR data analysis.
This study was conducted in the field in a nonstandard clinical situation and was thus subject to the variables of fieldwork. Field investigations, unlike controlled clinical environments, involve unmanageable variables and constraints, such as participants' school schedules, time limitations, and availability. Because of some of these constraints, all standard clinical ASR measures could not be performed on every participant in the study groups.
Altitude I Group
The Altitude I Group comprised 52 children residing at 2850 m above sea level in the Andes Mountains of Ecuador. Two children were excluded from the data analysis because one was found to have a bilateral mild conductive hearing loss, and a second child had high tympanic membrane compliance (3.3–3.5 cm3), both conditions that could potentially influence the middle ear immittance change related to the contraction of the stapedius muscle. The remaining 50 children in this group ranged in age from 3.0 to 17.0 years, with a mean age of 9.8 years (±3.4 years) and a median age of 10.0 years. The children in the Altitude I group consisted of 22 males (age range: 3.0–17; mean age: 10.4; SD: 3.9; median: 10.8 years) and 28 females (age range: 5.0–15.08; mean age: 9.4; SD: 2.8; median: 10.0 years). The participants in the Altitude I group consisted mainly of children of indigenous Quechua Indian ethnicity and mixed Spanish/indigenous Indian backgrounds, who had lived all of their lives at Altitude I.
Altitude II Group
The ASR in the Altitude II Group was investigated in 47 children residing at 3973 m (3500–4200 m) above sea level in the village community of Guangaje. The 47 children ranged in age from 2.0 to 17.0 years, with a mean age of 9.9 years (±3.5 years) and a median age of 10.0 years. The higher altitude group consisted of 21 males (age range: 2.0–15.67; mean age: 9.7; SD: 3.8; median: 10.0 years) and 26 females (age range: 4.6–17.0; mean age: 10.1; SD: 3.3; median 9.8 years). In a subsample of 22 children in the Altitude II Group, who ranged in age from 2 to 17 years (mean age: 10.1 ± 3.7; median: 10.6 years), SaO2 levels and heart rate data were obtained. The children in the Altitude II group were of indigenous Quechua Indian background and have lived all of their lives at Altitude II.
Informed consent was obtained from the parents or guardians of all children before testing. This study was conducted under the auspices of the Universidad San Francisco de Quito Medical School in Quito, Ecuador, and was approved by the Human Studies Committee (Comité de Bioética) of the Universidad San Francisco de Quito.
ASR test procedures
Each participant underwent a conventional audiological/otological examination, including otoscopy, tympanometry, and pure-tone threshold measurements. The specific parameters of the ASR that were assessed included ASRT, ASR amplitude growth (ASRG), and duration of the ASR peak contraction before decay/fatigue (ASRD). In addition, onset contraction time and offset relaxation time were calculated. The ASR was measured indirectly with a middle ear impedance/immittance instrument by recording the attendant change in the impedance/immittance of the middle ear system induced by contraction of the stapedius muscle.
The ASRT, ASRG, and ASRD were recorded by the TympStar immittance system (Grason-Stadler, Eden Prairie, MN) using a probe tone frequency of 226 Hz. The probe was hermetically sealed in the external ear canal, and tympanometry was performed to examine the status of the tympanic membrane and the middle ear system. The stimulus activators used to elicit the ASR were 500, 1000 Hz and broadband noise (BBN) (bandwidth = 125–4000 Hz). To measure the uncrossed ASR (ipsilateral ASR), the probe stimulus and the activator stimuli were delivered to the same ear. To measure the crossed brainstem reflex arc (contralateral ASR), the probe tone was presented to one ear and the reflex activator stimuli were delivered to the contralateral ear. The ASRT, ASRG, and ASRD were measured on each participant where possible. The ASRT was defined as the lowest stimulus intensity level at which a detectable and reproducible ASR deflection from the recording baseline (representing a minimum of 0.01 cm3 change in immittance/impedance) could be detected. Stimulus presentations of increasing intensity increments of 5 dB were used to evoke the ASR to measure amplitude growth over a range of 5–15 dB above threshold (sensation level) to evaluate the dynamic range or magnitude of the stapedius muscle contraction. The ASRD was defined as the time in seconds at which the stapedius muscle contraction amplitude declines or decays to at least 50% of its initial peak contraction amplitude over a 10-second stimulation period. Thus, a reduction in response amplitude of 50% or greater at any point during a 10-second period of acoustic stimulation (i.e., ASR half-life of <10 seconds) was considered abnormal fatigue or abnormal muscle adaptation. Figure 1A and B shows ASRT and ASRD recordings from two participants illustrating uncrossed (A) and crossed (B) physiological responses for a 1000 Hz activator stimulus and a BBN activator stimulus, respectively.

ASRT and ASRD recordings from two participants illustrating uncrossed
Before deployment in the field in Ecuador, the TympStar system was calibrated in the laboratory for pressure, volume, compliance, activator frequency and sound pressure level, activator noise band and sound pressure level, and probe tone frequency and sound pressure level in accordance with the American National Standards Institute (ANSI, 1987) specifications. Daily volume calibrations of the TympStar system were performed with a calibrated test cavity, and biological calibrations were made in the field on normal hearing staff to continually monitor the status of the equipment.
To determine if contraction (onset) and relaxation (offset) properties of the stapedius muscle were related to altitude, contraction and relaxation parameters of the ASR were determined in a manner similar to that of Shimizu et al. (1992) and Osanai et al. (2001). The contraction parameters of the ASR were examined by analyzing the time at which the response amplitude increased to 50% of the maximum amplitude following activator stimulus onset. The muscle relaxation time was examined by measuring the time from peak amplitude to a reduction of 50% of the maximum amplitude following activator stimulus offset.
Statistical analysis
For statistical analysis, the means, SDs, medians, ranges, and percentiles were calculated for the ASRT, ASRG, and ASRD. Because some of the variables were found to have skewed distributions, nonparametric statistical tests were used for data analysis. The Mann–Whitney U test was used to evaluate the difference between the Altitude I and the Altitude II groups for the ASRT, ASRG, and ASRD for the uncrossed and crossed ASR muscle responses. The Mann–Whitney U test also was used to probe for differences between the Altitude I Group and sea level Reference Group II, and between the Altitude II Group and sea level Reference Group II for ASRT and ASRG. The Wilcoxon Signed Rank Test for paired comparisons was used to assess the difference between the uncrossed and crossed ASRT within the groups. Associations of altitude with ASRT, ASRG, ASRD, SaO2, and heart rate were probed with the Spearman correlation coefficient. An alpha level of ≤0.05 was accepted as statistically significant.
Results
Acoustic stapedius reflex thresholds
The uncrossed and crossed ASRT for the Altitude I and the Altitude II groups are compared in Tables 1–3 for 500, 1000 Hz and BBN reflex activator stimuli, respectively. An analysis using the Mann–Whitney U revealed that the mean uncrossed ASRT (the smallest signal intensity eliciting a muscle contraction) for 500 Hz (p = 0.864), 1000 Hz (p = 0.961), and BBN (p = 0.639) were not significantly different for the Altitude I and the Altitude II groups. The crossed ASRT also was not statistically different for the two altitude groups for 500 Hz (p = 0.245), 1000 Hz (p = 0.997), and BBN (p = 0.855) reflex activator stimuli. Furthermore, it can be seen from Tables 1–3 that the mean ASRT for both groups tended to be better (lower ASRT) for the uncrossed condition than the crossed condition. For the Altitude I group, the uncrossed ASRT was significantly better by ∼4.5–7 dB (Tables 1–3) than the crossed ASRT (Wilcoxon Signed Rank Test) for 500 Hz (p = 0.0006), 1000 Hz (p ≤ 0.0001), and BBN (p ≤ 0.0001) reflex activator stimuli.
SD values are shown in parentheses.
ASRT, acoustic stapedius reflex thresholds; HL, hearing level; SD, standard deviation.
SD values are shown in parentheses.
SD values are shown in parentheses.
BBN, broadband noise.
The higher altitude group residing at 3973 m (Altitude II Group) showed no significant difference between the uncrossed and crossed ASRT for the 500 Hz (p = 0.122) reflex activator stimulus. However, the Altitude II Group showed significantly better uncrossed than crossed ASRT for the 1000 Hz (p = 0.0002) and BBN (p ≤ 0.0001) reflex activator stimuli, with the differences ranging from 4.8 to 7.5 dB.
Figure 2 compares the mean uncrossed and crossed ASRT for the Altitude I and Altitude II groups to sea level data obtained on young adults for 500 Hz, 1000 Hz, and BBN activator stimuli by Wiley et al. (1987) (Reference Group I). The broken lines in the graphs represent the ±2 SD range of Reference Group I. The ASRT for the Altitude I and Altitude II Groups tended to be higher than Reference Group I, but both groups were within the ±2 SD range of Reference Group I. The crossed ASRT data and ASRG function for the Altitude I Group and the Altitude II Group for 1000 Hz were also compared to results obtained by the authors on a sea level group of 22 children (Reference Group II, age range: 7–17.6 years; mean: 12.5; SD: 3.1; median: 12.2 years). The mean crossed ASRT at 1000 Hz for this sea level group was 85.4 dB (SD: 5.1; median: 85 dB), which was significantly better by 6 dB than the crossed ASRT of 91.7 dB for the Altitude I Group (Mann–Whitney U, p = 0.0002) and the Altitude II Group (p = 0.0004). Nevertheless, the crossed ASRT for the activator stimulus of 1000 Hz, which was identical (91.7 dB) for the Altitude I Group and the Altitude II Group, were within ±2 SD of the sea level Reference Group II.
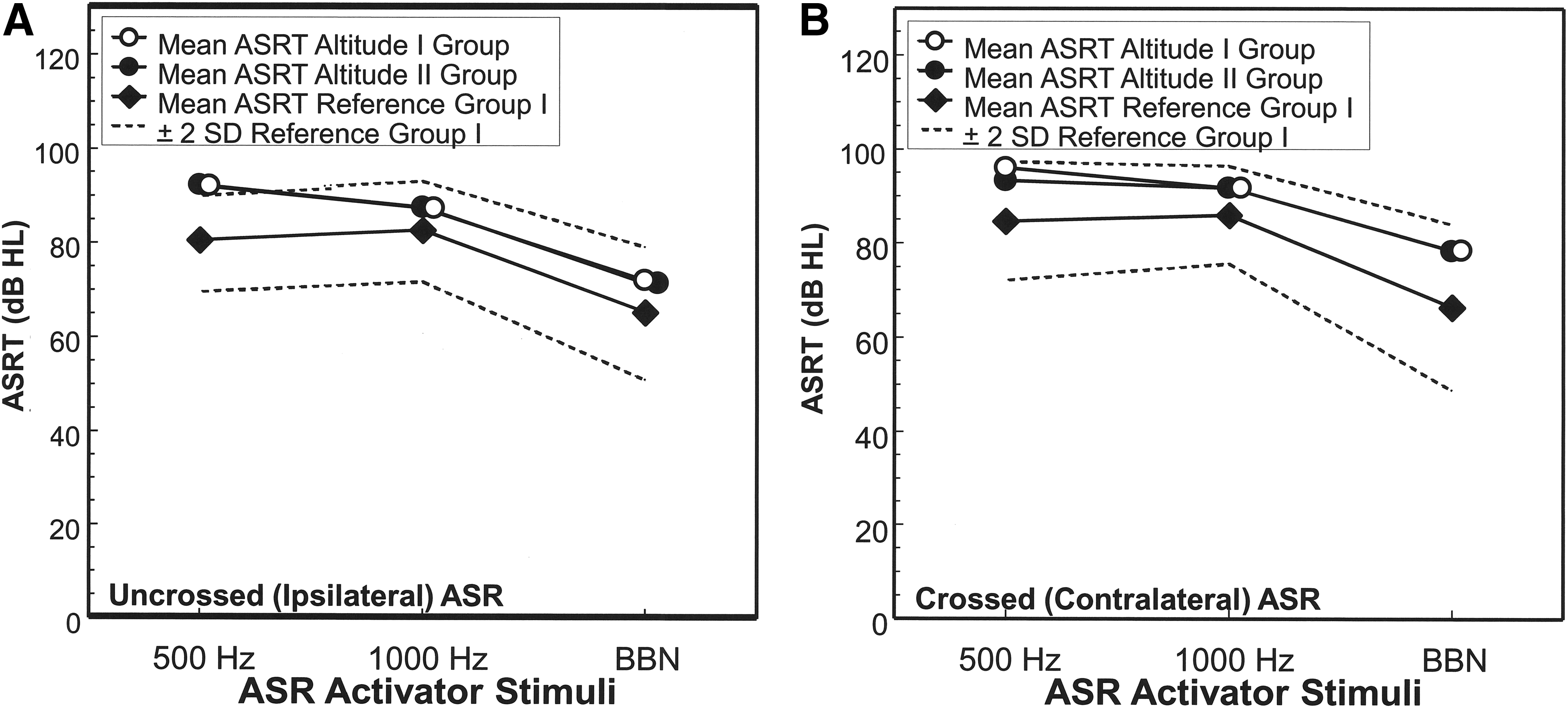
Mean ASRT in dB HL for the Altitude I and Altitude II groups for three ASR activator stimuli (500, 1000 Hz and BBN, bandwidth = 125–4000 Hz). The results for activation of the uncrossed (ipsilateral) ASR are shown in graph
ASRG function
The ASRG function of the Altitude I and the Altitude II groups is displayed in Figure 3A–C for the uncrossed reflex as a function of the three activator stimuli (500, 1000 Hz and BBN). The ASRG function for the crossed reflex profile of the two altitude groups is displayed in Figure 4A–C for the 500, 1000 Hz and BBN activator stimuli, respectively. Figure 3 revealed that the uncrossed ASRG for the Altitude I and the Altitude II groups is similar and both show essentially a linear growth function. Statistical analyses (Mann–Whitney U) revealed no statistically significant differences (p ranged from 0.221 to 0.920) between the Altitude I and the Altitude II groups for the ASRG for the three reflex activator stimuli during activation of the uncrossed ASR. Figure 4 shows that the crossed ASRG function tends to be less steep compared with the uncrossed ASRG, but the crossed ASRG is similar for the Altitude I and Altitude II groups for each of the three activator stimuli (p ranged from 0.213 to 0.997). Comparing the crossed ASRG function at 1000 Hz for the Altitude I Group and the Altitude II Group with that of the sea level Reference Group II participants yielded no significant differences.
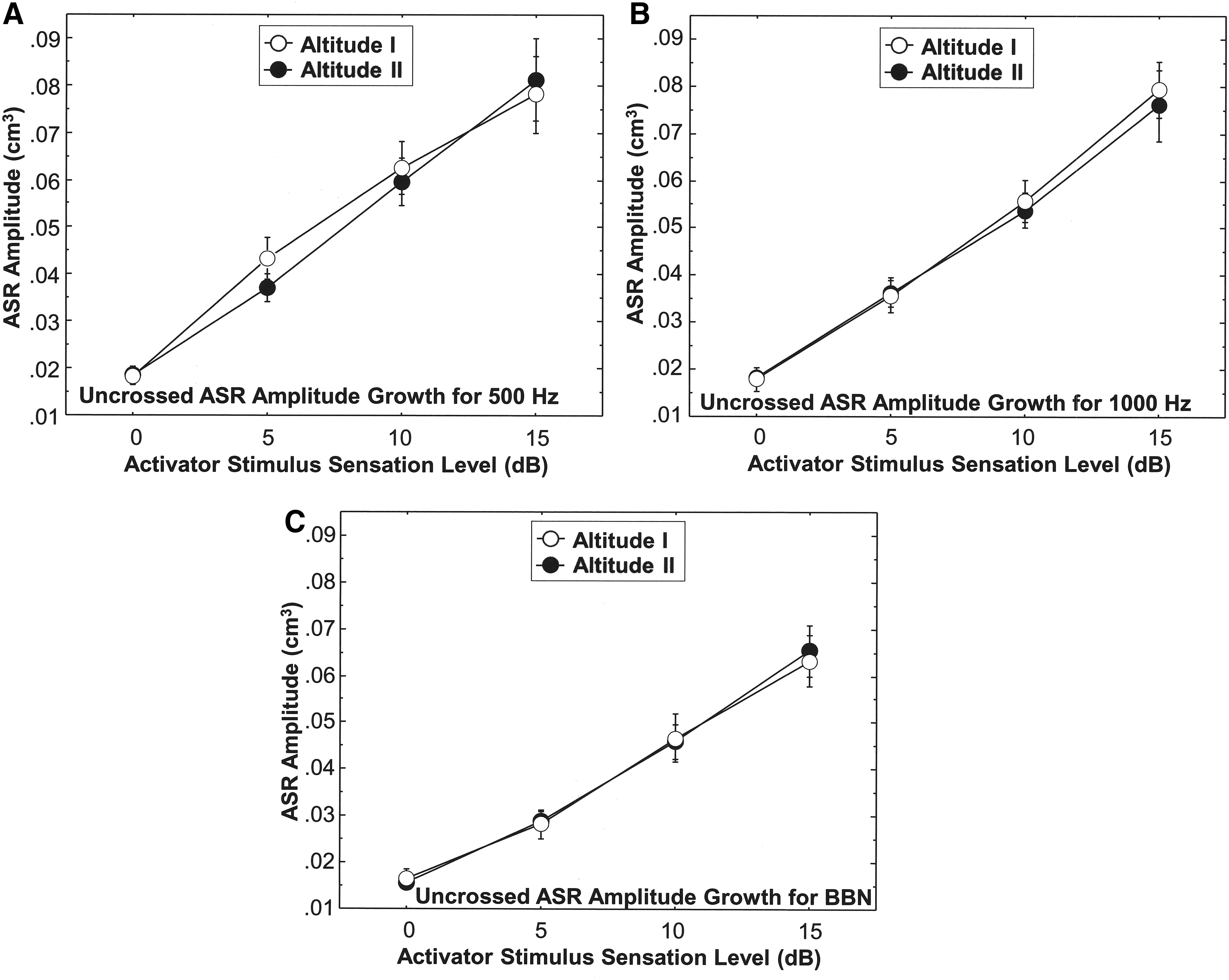
Uncrossed (ipsilateral) ASR ASRGs for three stimulus activators (
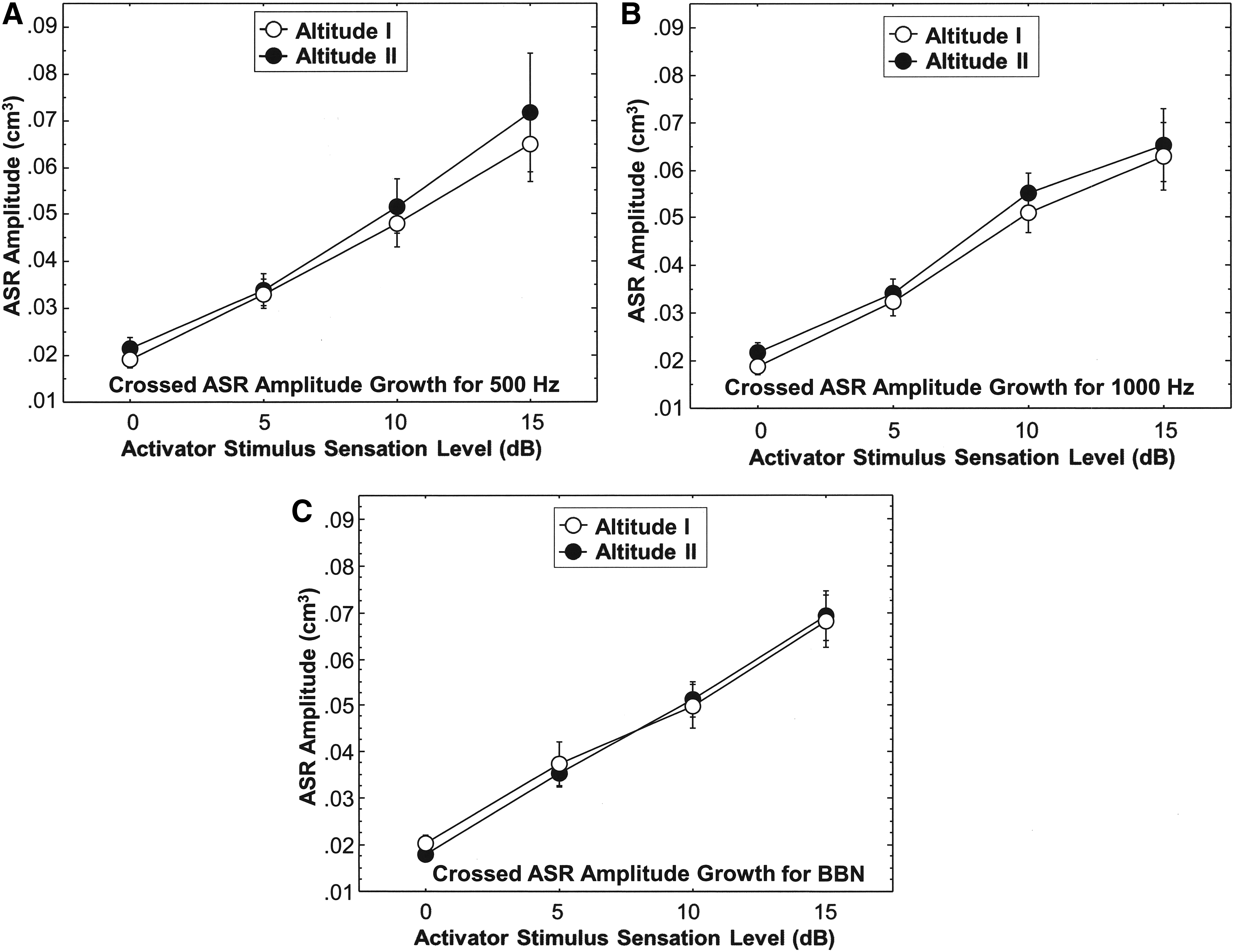
Crossed (contralateral) ASR ASRGs for three stimulus activators (
ASR muscle contraction decay/fatigue (ASRD)
The ASRD is defined clinically as the time in seconds it takes for the amplitude of the stapedius muscle contraction to diminish to 50% of its initial peak contraction amplitude during a 10-second recording period or an ASR half-life of less than 10 seconds. The ASRD is recorded in seconds, and any muscle contraction duration significantly less than 10 seconds is considered abnormal fatigue/decay. The ASRD times for the Altitude I and the Altitude II groups for uncrossed and crossed ASR stimulation for the three activator stimuli (500, 1000 Hz and BBN) are displayed in Table 4. Although the higher Altitude Group (Altitude II Group) tended to show a faster (7.7 seconds vs. 8.9 seconds) uncrossed ASR decay time than Altitude I Group for a 500 Hz activator stimuli, the difference did not reach statistical significance (Mann–Whitney U, p = 0.115). The crossed ASRD did not show a statistically significant difference between the two altitude groups at 500 Hz (p = 0.492). The difference in ASRD between the Altitude I and Altitude II groups for a 1000 Hz reflex activator stimulus was not statistically significant for the uncrossed reflex (p = 0.359) or the crossed reflex (p = 0.820). The crossed ASRD for BBN also was not significantly different (p = 0.667) for the Altitude I and Altitude II groups.
The mean and median data in the table indicate the number of seconds the ASR maintained a contraction over a 10-second sustained stimulation period. SD values are shown in parentheses, and median values are shown in brackets.
SaO2 level and heart rate
In a subsample of 22 children (age range: 2–17; mean age: 10.1 ± 3.7; median: 10.6 years) living at Altitude II (3973 m), SaO2 levels and heart rate data were obtained. Figure 5 shows the distribution of SaO2 data for the 22 participants from the Altitude II group. The mean SaO2 in the children was 85.2% (SD: 6.5; range: 67–93; median: 86.5), and the mean heart rate was 91.0 (SD: 21.2; range: 49–134; median: 93.0). There was no significant association between SaO2 and heart rate, or between SaO2 and ASRT, ASRG, or ASRD.
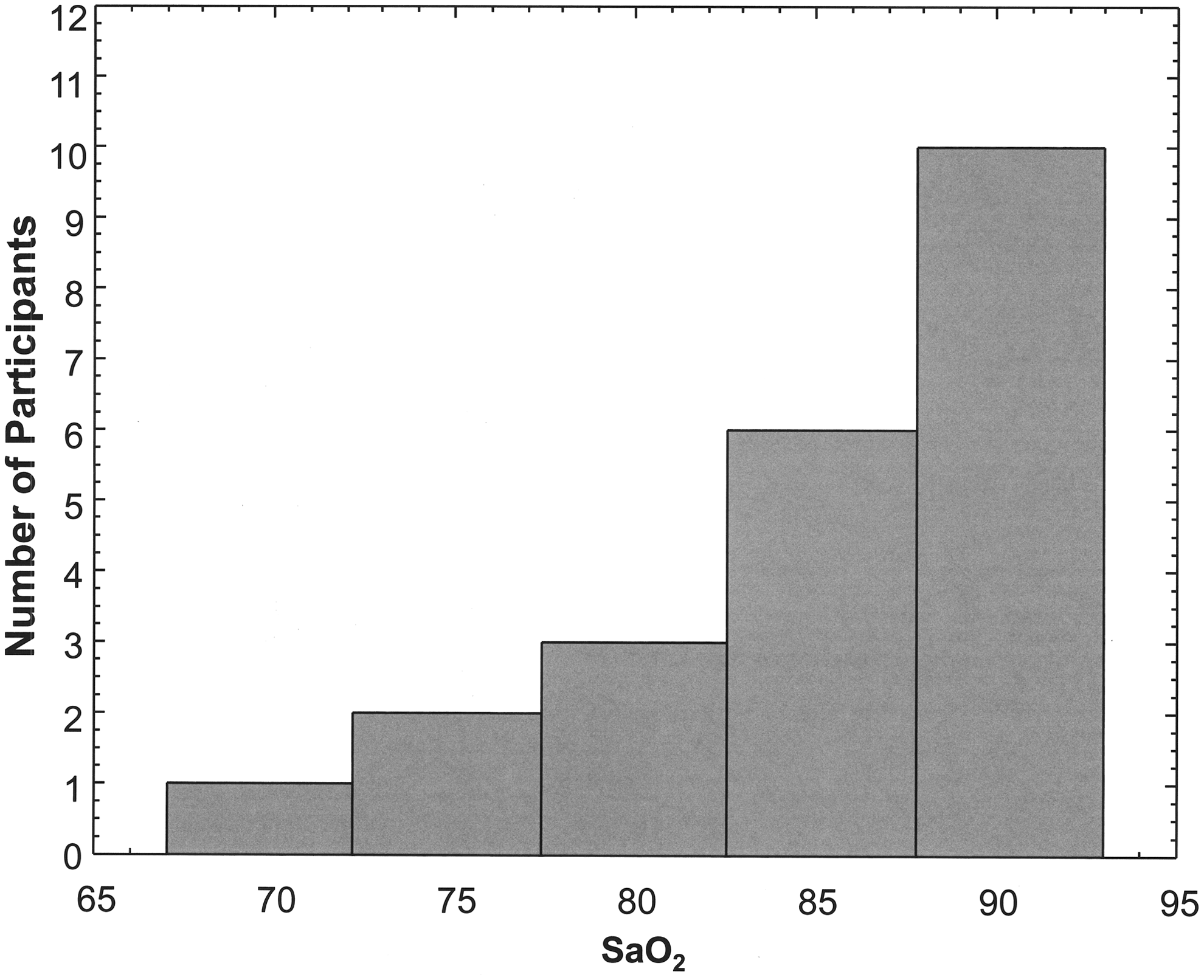
Distribution of SaO2 levels in children living at Altitude II in the villages of Guangaje, Ecuador. SaO2, oxygen saturation.
Contraction and relaxation dynamics of the ASR
The contraction and relaxation dynamics of the ASR were examined for the crossed condition by determining the time at which the muscle achieved 50% of its peak contraction following stimulus onset and the time required for the muscle contraction to diminish to 50% of its peak amplitude (relaxation time) following activator stimulus offset, respectively. The results showed that for both the Altitude I and Altitude II Groups, the stapedius muscle 50% contraction time was significantly longer than the muscle 50% relaxation time for the 500 Hz (Wilcoxon Signed Test, p ≤ 0.0001), 1000 Hz (p ≤ 0.0001), and BBN (p ≤ 0.0001) activator stimuli. The ASR 50% contraction time was 1.8–1.9 times longer than the 50% relaxation time.
As indicated above, both altitude groups had faster 50% relaxation times than 50% contraction time, but the Altitude I Group showed a significantly (p = 0.027) faster muscle 50% relaxation time than the Altitude II Group for the BBN activator stimulus. However, there were no significant differences between the two altitude groups and muscle relaxation times for 1000 and 500 Hz. The two groups did not differ in muscle 50% contraction dynamics for any of the three activator stimuli. The ASR muscle contraction and relaxation times did not correlate with SaO2 level.
Discussion
Adverse medical and oto-physiological effects of high altitude environments have been reported at altitudes of 2200–2500 m and above (Cingi et al., 2010; Dursun, 2008). Studies of the oto-physiological effects of low oxygen environments have reported a number of anomalous biomedical conditions, including otological disorders affecting the middle ear, inner ear, and auditory brain (Laurikainen et al., 1990; Fowler and Lindeis, 1992; Fowler and Prlic, 1995; Cingi et al., 2010; Dursun, 2008). Most of these studies investigated high altitude physiological effects on subjects in simulated low oxygen environments and on non-native high altitude dwellers who ascended from sea level to higher elevations. Although indigenous people who are native high altitude dwellers may exhibit adaptations to low oxygen conditions, their otological status and the physiology of their auditory system may be comprised.
The current study investigated the integrity of the auditory system in children living at high altitudes, using the ASR as a neurodiagnostic metric. The ASR is a noninvasive clinical neurophysiological measure, which yields diagnostic and physiological information about the middle ear, inner ear, and the ascending auditory pathways, including the multisynaptic brainstem arc of bipolar afferent neurons of the auditory nerve projecting to the cochlear nuclei, the uncrossed and crossed superior olivary nuclei, axonal projections to neurons in and around the nucleus of the seventh cranial nerve, and the efferent motoneurons of the facial nerve branch to the stapedius muscle. This study examined the threshold, amplitude growth, and contraction duration of the ASR in a cohort of children living at high altitudes (∼2800 and 4000 m) in the Andes Mountains.
The findings of this study showed that the children living at 2800 m (Altitude I Group) and at around 4000 m (Altitude II Group) above sea level had ASRT within the normal range, and there were no statistically significant differences between the two groups. Furthermore, there were no differences in the ASRT data for the two altitude groups of the current study and normative data from subjects tested at sea level. The results also showed sustained contraction of the stapedius muscle without substantial degradation or fatigue in the study groups using clinically established norms. There were no significant differences between the Altitude I and Altitude II groups in uncrossed and crossed stapedius reflex growth functions (ASRG), and the ASRG increase was consistent with normal growth patterns at sea level.
For the BBN activator stimulus, the Altitude I Group showed a significantly faster stapedius muscle relaxation time, that is, from peak contraction amplitude to a 50% reduction in amplitude, than the Altitude II Group. Because BBN activates frequency tuned receptors in the entire cochlea and a greater number of neurons in the tracts and nuclei of the auditory brainstem, it may be a better acoustic stimulus for evaluating the stapedius muscle reflex than pure-tone activator stimuli.
A previous study (Laurikainen et al., 1990) found that experimentally induced hypoxia in apparent sea level dwellers modulated the amplitude of the ASR and was associated with SaO2 levels. In the current study, threshold, amplitude, and decay parameters of the ASR did not correlate with SaO2 levels. The Laurikainen et al. (1990) study, however, differed in important ways from the current study in that it used experimentally induced hypoxia in sea level dwellers, whereas the current study investigated the ASR variables in native high altitude dwellers. Individuals who have lived all their lives at high altitudes have developed high-altitude adaptive physiological mechanisms or acclimatization and, thus, are expected to have different responses to low oxygen environments (Scheinfeldt and Tishkoff, 2010; Simonson, 2015). In addition, the participants in the Laurikainen et al. (1990) investigation experienced lower SaO2 levels than the participants in the current study.
The mean SaO2 level was 85.2% in the present study, which is lower than the 91.3% SaO2 level found by Huicho et al. (2001) in a population comprised mostly of indigenous Quechua Indians who reside in the Peruvian Andes at 4100 m. The ethnic background and the altitude of the Peruvian Quechua Indians are similar to that of the participants in the current study. Although the current study showed relatively low SaO2 levels compared to the Peruvians, there were no significant adverse effects on the physiology of the ASR, suggesting that the small skeletal muscle can tolerate relatively low SaO2 levels. The heart rate of 91.0 in the current study was higher than that in the Huicho et al. (2001) study of 84.8, but similar to the SaO2 results, heart rate had no significant effect on the functioning of the ASR. Using the formula P = antilog10 [(53012.2 − h)/18401.8], where P = atmospheric pressure in mm Hg and h = elevation above sea level in meters (Lilly and Shanks, 1981), we calculated the atmospheric pressure at 2850 m to be 532.03 mm Hg and at 3973 m to be 462.29 mm Hg. Atmospheric pressure differences between the Altitude I and Altitude II groups did not appear to have any effect on the measured ASR parameters.
Hypobaric hypoxia is associated with high altitude and cellular hypoxia in oxidative tissues, including skeleton muscles. The stapedius, the smallest human skeleton muscle, is at the end of an afferent–efferent motoneuron tract. It may be hypothesized that oxidative stress, which is associated with decreased mitochondrial function, is less apparent in persons living at high altitude from birth who have acclimatized to these conditions than persons living at sea levels (Levett et al., 2012).
This noninvasive study utilized the contraction parameters of the stapedius muscle, including threshold, amplitude growth, and muscle contraction duration as indices of cellular and physiological function in a pediatric population living at high altitudes in the Andes Mountains of Ecuador. The findings showed no adverse effects of altitude on the ASR up to 4000 m. Additional studies of the stapedius reflex in persons residing at altitudes higher than 4000 m may provide further information on the physiological functioning of the stapedius muscles at high altitudes. In addition, performing repeated measures of the ASR in participants before and after traveling to high altitudes may yield additional information on ASR physiology and participants' time course of altitude acclimatization.
Footnotes
Acknowledgments
The authors thank the administration of Universidad San Francisco de Quito, Fundacíon Capacitar of Ecuador, and the Ecuadorian health authorities for continued support of this project. The authors are grateful to the Harvard David Rockefeller Center for Latin American Studies for support of this project. The authors thank Professor Erik Borg for helpful comments and suggestions. The authors thank Dr. Jeremy Bloxham, Dean of Science at Harvard, the Harvard Biological Laboratories, and the Harvard University Health Services for support.
Author Disclosure Statement
No competing financial interests exist.