
Abstract
Abstract
Zhang, Jiaxing, Ji Chen, Cunxiu Fan, Jinqiang Li, Jianzhong Lin, Tianhe Yang, and Ming Fan. Alteration of spontaneous brain activity after hypoxia-reoxygenation: A resting-state fMRI study. High Alt Med Biol. 18:20–26, 2017.—The present study was designed to investigate the effect of hypoxia–reoxygenation on the spontaneous neuronal activity in brain. Sixteen sea-level (SL) soldiers (20.5 ± 0.7 years), who garrisoned the frontiers in high altitude (HA) (2300–4400 m) for two years and subsequently descended to sea level for one to seven days, were recruited. Control group consisted of 16 matched SL natives. The amplitude of low-frequency fluctuations (ALFF) of regional brain functional magnetic resonance imaging signal in resting state and functional connectivity (FC) between brain regions was analyzed. HA subjects showed significant increases of ALFF at several sites within the bilateral occipital cortices and significant decreases of ALFF in the right anterior insula and extending to the caudate, putamen, inferior frontal orbital cortex, temporal pole, and superior temporal gyrus; lower ALFF values in the right insula were positively correlated with low respiratory measurements. The right insula in HA subjects had increases of FC with the right superior temporal gyrus, postcentral gyrus, rolandic operculum, supramarginal gyrus, and inferior frontal triangular area. We thus demonstrated that hypoxia–reoxygenation had influence on the spontaneous neuronal activity in brain. The decrease of insular neuronal activity may be related to the reduction of ventilatory drive, while the increase of FC with insula may indicate a central compensation.
Introduction
S
The brain continuously maintains a high level of spontaneous activity, which is proved to be relevant for human behavior such as action, perception, and higher cognitive function (Sadaghiani and Kleinschmidt, 2013; Romano et al., 2015; Tan, 2015; Visintin et al., 2015). Injury in sensory afferent pathway contributes to the change of cortical spontaneous activity (Alonso-Calviño et al., 2016). Highly synchronous low-frequency fluctuations (0.01–0.08 Hz) of BOLD signals among motor cortices were first observed by Biswal et al. (1995) and then the ALFF analysis was developed by Zang et al. (2007). ALFF analysis has been widely used in the studies of mental disorders (Hoptman et al., 2010; Qi et al., 2015) and cognitive impairments (Fryer et al., 2015; Gao et al., 2015).
Recently, we have investigated the brain structures in 16 sea-level (SL) soldiers who have garrisoned the frontiers in HA for two years (Zhang et al., 2013). However, although none of these soldiers felt obvious mental discomfort [a sign of “HA deadaptation reaction,” suffering of some people shortly after their return to lowland from HA (Zhou et al., 2012)], we still found the changes of local synchronization of resting-state signals in these soldiers seven days within their return to SL using functional magnetic resonance imaging (fMRI) (Chen et al., 2016). In the present study, these soldiers were further examined ALFF to confirm the change of spontaneous brain activity. Resting-state functional connectivity (FC) of the brain regions was also detected. FC refers to temporal correlation between remote brain regions. Abnormality in FC is thought to reflect pathophysiological processes, and between-subject differences in FC correlate with individual traits and behavioral characteristics (Andrews-Hanna et al., 2007; Zuo et al., 2010). We hypothesized that the ALFF and FC in the brains of these soldiers could be influenced by both the increase of vascular density induced by long-term HA hypoxia exposure and the change of blood oxygen level resulting from hypoxia/reoxygenation.
Methods
Subjects
Sixteen male healthy soldiers (20.5 ± 0.7 years) (natives at Xiamen, China, <10 m), who garrisoned the frontiers in Qinghai-Tibet Plateau (2300–4400 m) for two years and subsequently moved to Xiamen for one to seven days, were studied. They all had no chronic mountain sickness at HA. Sixteen male subjects (19.9 ± 1.5 years), matched with age, education, and race, were recruited from Xiamen as controls. All subjects were right handed and nonsmokers and had no documented head injury or neurological disorders. The detailed information about subjects has been described in our previous study (Zhang et al., 2013). All subjects were given written informed consent to participate in the study, which had been approved by the Ethics Committee of Xiamen University.
Physiological assessments
Pulmonary function and hematological measurement were measured in Zhongshan Hospital Xiamen University on the same day with the MRI scanning. Blood samples were taken in the morning between 7:00 and 7:30. Hematological measurement was tested using a Blood GAS Analyzer (AVL compact 3, Switzerland). Pulmonary function was tested using a pulmonary function testing device (MasterScreen Body, JAEGER, Germany). Independent t-test was used to analyze between-group differences. Statistical significance was set at p < 0.05.
MRI data acquisition
Brain images were scanned on Siemens Trio Tim 3.0T (Erlangen, Germany) at Magnetic Resonance Center in Zhongshan Hospital Xiamen University. A 3D T1-weighted MPRAGE sequence was: TR/TE = 1900 ms/2.7 ms, NEX = 1, matrix = 256 × 246, FOV = 25 × 25 cm2. Conventional 2D T1 and T2 images were also acquired for examining any incidental findings.
Resting-state fMRI images were obtained using echo-planar imaging sequence with the following parameters: TR/TE = 3000 ms/30 ms, flip angle = 90°, matrix = 64 × 64, voxel size = 3.4 × 3.4 × 3.75 mm3, FOV = 24 × 24 cm2, slices = 38, and slice thickness = 3 mm. All subjects were introduced to lie still in the scanner with their eyes closed and keep awake. The data analyses were conducted by two researchers who were blind to the status of subjects.
Analysis of ALFF
Images were first processed using the data processing assistant for resting-state fMRI in SPM8 (www.fil.ion.ucl.ac.uk/spm), including the discard of the first four time points, head-motion correction, and the images smoothed using a 10 mm FWHM Gaussian kernel. Then the removal of linear trends of the time series and the temporal band-pass filtering (0.01–0.08 Hz) were performed to reduce the effects of the respiratory and cardiac rhythms. After temporally filtering for each subject, nuisance signals (white matter, cerebrospinal fluid, and six motion parameters) were regressed out. We derived the white matter/cerebrospinal fluid nuisance signals averaging the time courses of the voxels in each subject's white matter/cerebrospinal fluid masks. These masks were generated by the segmentation process of each subject's brain.
ALFF calculation was performed using REST (http://restfmri.net). Each filtered voxel's time series was transformed to the frequency domain by a fast Fourier transform to get power spectrum. The square root was calculated at each frequency of the power spectrum and averaged across 0.01–0.08 Hz at each voxel. This averaged square root was taken as the ALFF. The ALFF of each voxel was divided by the global mean ALFF value for the purpose of standardization. Two sample t-test was used to assess between-group difference, with age and education as covariates. Correction for multiple comparisons was done using AlphaSim program determined by Monte Carlo simulation in REST. A corrected significance level of p < 0.05 was obtained by combined individual voxel threshold with a minimum cluster size of 384 voxels (individual voxel p = 0.05, 1000 simulations, FWHM = 10 mm, within a gray matter mask generated from a gray matter probability image which set the threshold at 25%).
Seeding-based rest-state FC analysis
Given that the decrease of ALFF in the right insula had significant positive correlations with the pulmonary function measurements, the right insula was selected as a seed region.
Nuisance signals were regressed out for FC analysis; the procedures were the same as that in ALFF calculation. The mean time courses within the seed region were extracted by averaging over voxels and putting into REST to calculate the voxel-wise correlation coefficients of them with other regions in the brain. Each subject's correlation map was Fisher z-transformed and entered into group-level analysis. Statistical analysis was performed using SPM8 software for FC data, including one sample t-test and two sample t-test (controlling for age and education). For one sample t-test, probability maps were thresholded at p < 0.05, family-wise error corrected for multiple comparisons and combine cluster threshold k > 10 voxels in the native space; the two sample t-test resultant map was cluster level p < 0.05, corrected using Monte Carlo simulation in REST, and the parameters were the same as aforementioned in ALFF analysis.
Correlation analysis
Average ALFF value of all voxels in the right anterior insula was extracted using REST. In the HA group, correlations of pulmonary measurements with ALFF values were analyzed. The FC value in each of the regions that showed significant between-group difference with the seed was also extracted, and then the correlations of each FC value with the pulmonary measurements were analyzed. Statistical significance was set at p < 0.05 using Bonferroni's correction for multiple comparisons.
Results
Pulmonary and hematological measurements
The physiological values are shown in Table 1. Compared with controls, HA subjects had significant increases of heart rate, hemoglobin, and erythrocyte, while had significant decreases of pulmonary function in vital capacity, forced vital capacity (FVC), and forced expiratory volume in 1 second (FEV1).
Data are mean ± standard deviation.
FEV, forced expiratory volume; FEV1, forced expiratory volume in 1 second; FVC, forced vital capacity; HA, high altitude.
ALFF values
Compared with controls, HA subjects had significant increases of ALFF at several sites within the bilateral occipital cortex, including the superior, middle, and inferior occipital gyri, lingual gyrus, cuneus, and fusiform gyrus, and had significant decreases of ALFF in the right anterior insula and extending to the caudate, putamen, inferior frontal orbital cortex, temporal pole, and superior temporal gyrus (p < 0.05, corrected) (Fig. 1; Table 2). In addition, the ALFF regions showing group differences with statistical significance at p < 0.05 (uncorrected) and p < 0.01 (corrected) are shown in the supplementary figures (Supplementary Figs. S1 and S2). With statistical significance at p < 0.05 (uncorrected), the increases of ALFF in HA subjects expand beyond the occipital cortex and anterior insula to the bilateral temporal cortices and right anterior cingulate cortex (Supplementary Fig. S1). Moreover, the differences of ALFF at the bilateral occipital cortex and right anterior insula can survive more strict correction (p < 0.01, corrected) (Supplementary Fig. S2).
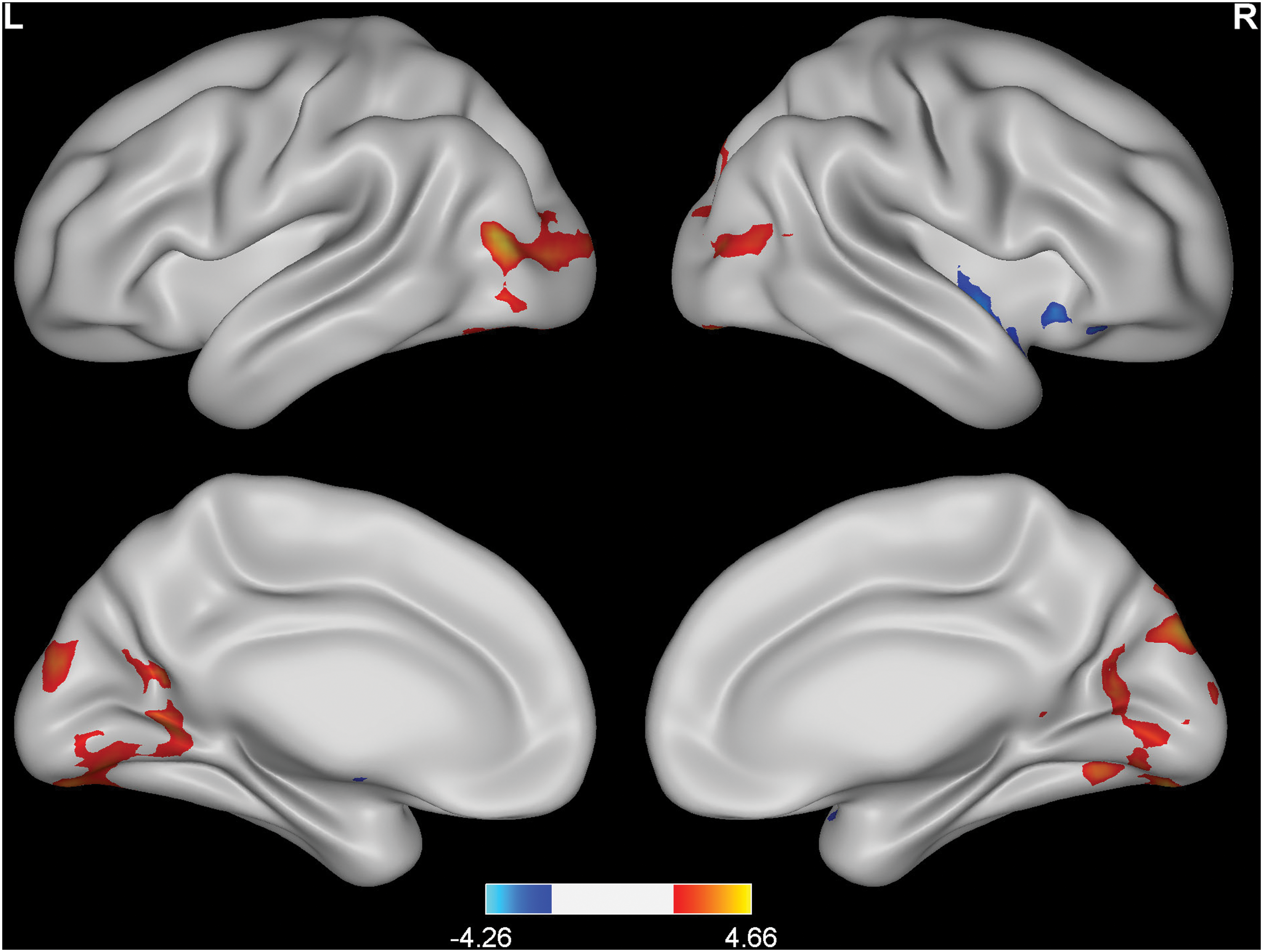
Changes in the resting-state ALFF in HA subjects compared with SL controls. p < 0.05 (corrected). Red to yellow indicates an increase; blue indicates a decrease. ALFF, amplitude of low-frequency fluctuations; HA, high altitude; SL, sea level.
In HA subjects, ALFF value in the right insula had significant positive correlations with predicted FVC and predicted FEV1 (Fig. 2). No significant correlations were detected between the ALFF values in the several occipital regions and the predicted FVC or predicted FEV1.
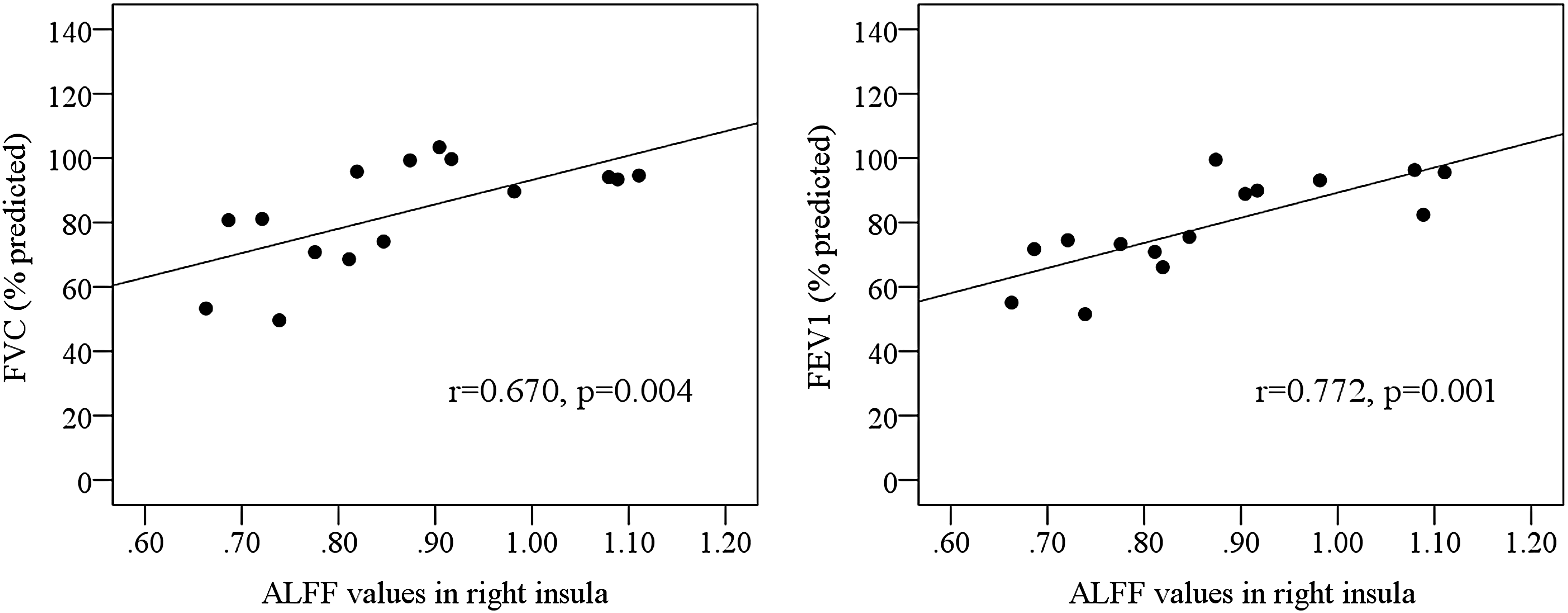
Correlations of the ALFF in the right insula with the forced vital capacity and forced expiratory volume in 1 second (FEV1).
FC of the right insula
In both SL controls and HA subjects, the right insula showed a bilateral pattern of connectivity (Fig. 3).
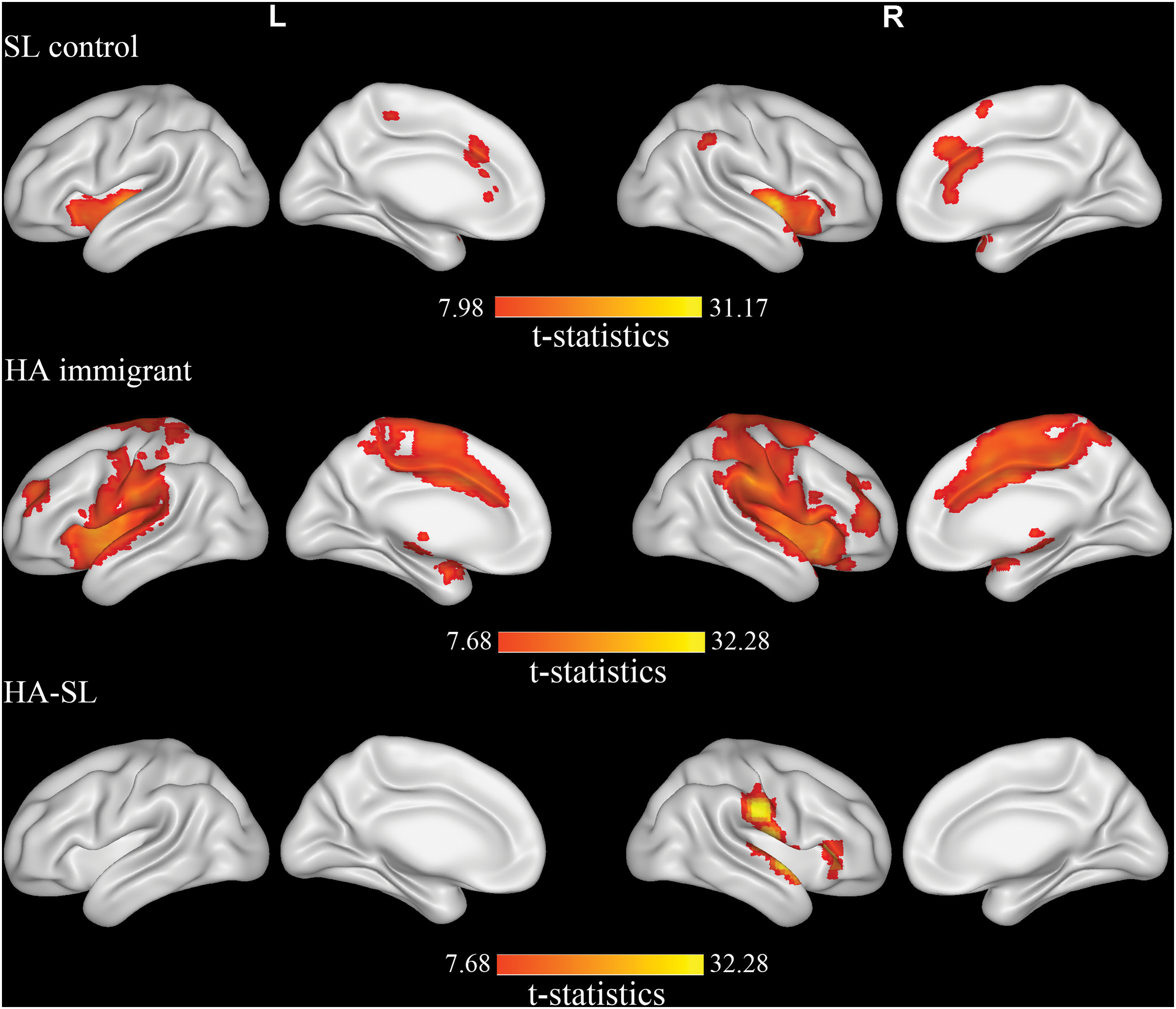
Regions showing the differences in functional connectivity of the right insula with other regions between HA subjects and SL controls. p < 0.05 (corrected), with cluster threshold k > 10 voxels. Red to yellow indicates an increase.
Compared with controls, the right insula in HA subjects had increases of FC with the right superior temporal gyrus, postcentral gyrus, rolandic operculum, supramarginal gyrus, and inferior frontal triangular area (Fig. 3).
The strength of FC of the right insula to the right superior temporal gyrus (Fig. 4a) and right rolandic operculum (Fig. 4b) showed significant positive correlations with the predicted FEV1.
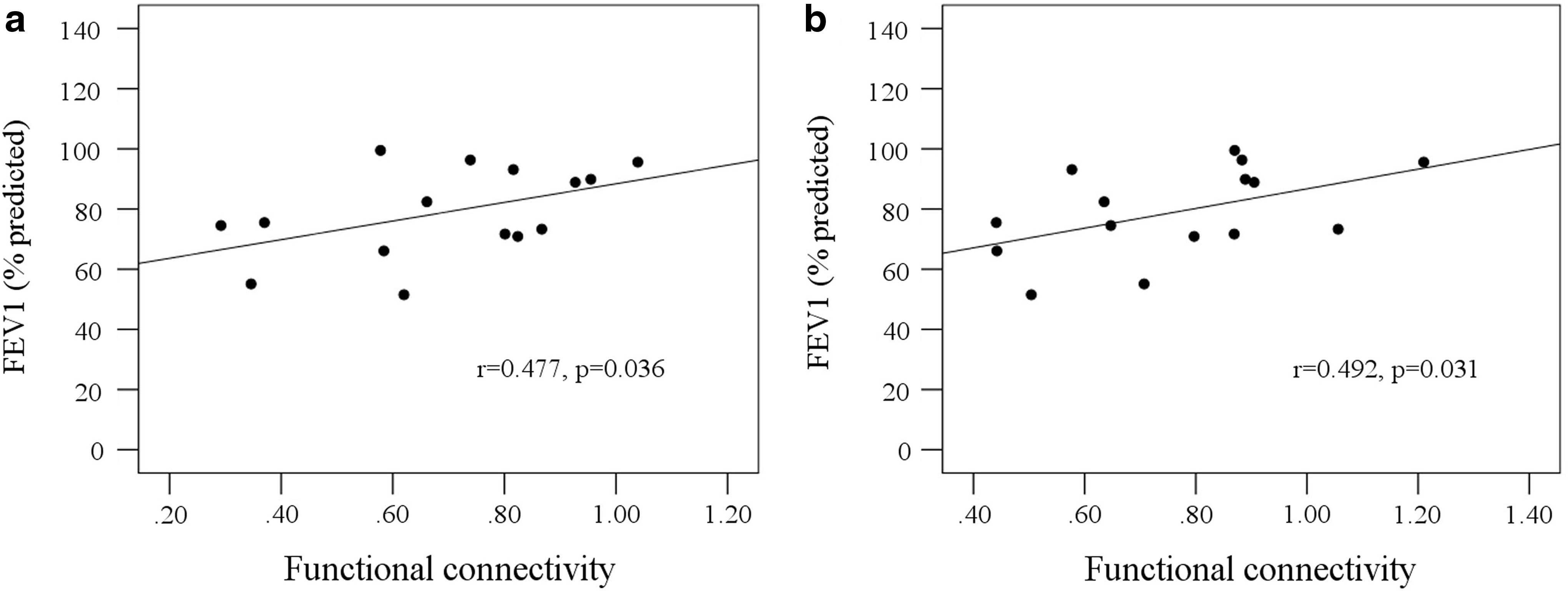
Correlations between FEV1 and the strength of functional connectivity.
Discussion
Our present study demonstrated that hypoxia–reoxygenation increased ALFF in several sites within the visual cortex, while decreased ALFF mainly in the right anterior insular cortex. Given that ALFF in the right insula had significant positive correlations with the lower pulmonary functions, we further analyzed FC of the right insula and found that the right insula in HA subjects showed increases of FC with the ipsilateral temporal, parietal, and frontal areas. Moreover, the strengths of this FC were significantly correlated with the pulmonary measurements.
Previous studies have found a significant positive correlation between ALFF and vascular density (Vigneau-Roy et al., 2014). Therefore, we speculated that the increase of ALFF within visual cortex may be related to the increase of vasculature. It is known that visual cortex is supplied by vertebral artery. However, vertebral artery is apt to be influenced by acute hypoxia. For example, Willie et al. (2012) found that with profound hypoxia (35 mmHg) the relative increase in vertebral artery blood flow was 50% greater than the other vessels; Ogoh et al. (2013) found that the internal carotid artery blood flow was unchanged, while vertebral artery blood flow significantly increased during acute hypoxia. After prolonged hypoxia, the adaptive increase of capillary occurs in visual cortex (LaManna et al., 2004). In fact, our previous findings of the increased cortical thickness in visual cortex on the same subjects may be due to the increases of vasculature (Zhang et al., 2013).
The change of gray matter induced by HA hypoxia may also contribute to the altered spontaneous neuronal activity. Previous multimodal neuroimaging studies have shed light on this function-structure association underlining cognition, aging, disease, and behavior (Sui et al., 2014; Yang et al., 2016). In our previous studies, the descendants of HA immigrants had reduced gray matter accompanied with longer delay in hemodynamic response to the maximum inspiration task in the bilateral anterior insula (Zhang et al., 2010; Yan et al., 2011). In the present study, we found the increase of ALFF within visual cortex, which has been shown the increase of cortical thickness in our previous study in the same subjects (Zhang et al., 2013). However, in the same subjects, we did not find an impairment of structure in insular cortex (Zhang et al., 2013).
Recently, we have recorded EEG activity on 25 lowland soldiers in eyes-closed resting condition at four time points: seven days at lowland before their ascending to HA, during the first 7 and 30 days at 3800 m altitude, and seven days after their return to lowland (Zhao et al., 2016). We found a persistent increase of beta power during HA acclimatization and after return to lowland, which was confined in the posterior parietal cortex and occipital cortex. Beta activity is an indicative of background excitation that involves a frequency potentiating mechanism at the level of synapse in a network (Whittington et al., 1997; Wróbel, 2000). Therefore, consistent with our previous EEG findings (Zhao et al., 2016), the increase of ALFF in visual cortex found in the present study suggested a sustained higher level of cortical excitation during hypoxic exposure and reoxygenation.
Insular cortex has fiber connections with the structures that related to respiratory sensation and control, including the rostral ventral respiratory cell group (ventrolateral part of the medulla oblongata) (Gaytan and Pasaro, 1998; Bowman et al., 2013), whisker barrel cortex (delta oscillations and gamma power in this area are linked to respiration) (Ito et al., 2014), and diaphragm (Lois et al., 2009). Injection of
The FCs of insula that showed in our study were also identified by Cauda et al. (2011) and Lacuey et al. (2016). These FCs have been detected in the previous cytoarchitectonics and tract tracing studies (Augustine, 1996). In the present study, the right insula in HA subjects enhanced its connectivity with the right superior temporal gyrus and right rolandic operculum. Superior temporal cortex was activated by acute hunger (Davenport and Vovk, 2009; Herigstad et al., 2011), while neuronal activity in it decreased in response to acute hunger in patients with congenital central hypoventilation syndrome (Macey et al., 2004) and, moreover, gray matter volume in it reduced in patients with asthma (Wang et al., 2014) and in patients with central hypoventilation syndrome (Harper et al., 2015). Animal study has shown that vagal stimulation produced response in the rolandic cortex, suggesting that this area involved in the processing of respiratory information (Davenport and Vovk, 2009). Taken together, the increased FC may be as a compensation of decrease in insular activation.
There are two limitations in our study. The first one is that we did not look at the brain before, during, and after HA exposure. The second one is that HA subjects had higher heart rhythms. The cardiac rate is normally in the 0.9–1.1 Hz ranges (Cordes et al., 2001). In our study, we only analyzed band-pass between 0.01 and 0.08 Hz to remove this higher frequency noise, and thus, the effects of very low-frequency drift and higher frequency noise were removed.
Conclusion
In summary, we demonstrated that hypoxia/reoxygenation changed the spontaneous neuronal activity and FC in brain. The present study, taken together with our previous findings that EEG beta power increased during HA acclimatization and after return to lowland (Zhao et al., 2016), suggests that hypoxia–reoxygenation can increase occipital excitation. The increased spontaneous neuronal activity within the visual cortex may be attributed to the increase of capillary induced by chronic hypoxia. Decrease of resting-state neuronal activity in the insular cortex may result in the reduction of ventilatory drive, while the increase of resting-state brain FC may indicate a central compensation.
Footnotes
Acknowledgments
This work was supported by National Science Foundation of China (Project No. 81171324; 81471630) and National Key Project (2012CB518200).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.