
Abstract
Abstract
Lüneburg, Nicole, Patricia Siques, Julio Brito, Juan José De La Cruz, Fabiola León-Velarde, Juliane Hannemann, Cristian Ibanez, and Rainer Böger. Long-term intermittent exposure to high altitude elevates asymmetric dimethylarginine in first exposed young adults. High Alt Med Biol. 18:226–233, 2017.—Hypoxia-induced dysregulation of pulmonary and cerebral circulation may be related to an impaired nitric oxide (NO) pathway. We investigated the effect of chronic intermittent hypobaric hypoxia (CIH) on metabolites of the NO pathway. We measured asymmetric and symmetric dimethylarginine (ADMA and SDMA) and monomethyl-L-arginine (L-NMMA) and assessed their associations with acclimatization in male draftees (n = 72) undergoing CIH shifts at altitude (3550 m) during 3 months. Sixteen Andean natives living at altitude (3675 m) (chronic hypobaric hypoxia [CH]) were included for comparison. In CIH, ADMA and L-NMMA plasma concentrations increased from 1.14 ± 0.04 to 1.95 ± 0.09 μmol/L (mean ± SE) and from 0.22 ± 0.07 to 0.39 ± 0.03 μmol/L, respectively, (p < 0.001 for both) after 3 months, whereas SDMA did not change. The concentrations of ADMA and L-NMMA were higher in CH (3.48 ± 0.07, 0.53 ± 0.08 μmol/L; p < 0.001) as compared with CIH. In both CIH and CH, ADMA correlated with hematocrit (r2 = 0.07, p < 0.05; r2 = 0.26; p < 0.01). In CIH, an association of ADMA levels with poor acclimatization status was observed. We conclude that the endogenous NO synthase inhibitors, ADMA and L-NMMA, are elevated in hypoxia. This may contribute to impaired NO production at altitude and may also be predictive of altitude-associated health impairment.
Introduction
M
In populations permanently living at high altitude (chronic hypoxia [CH]), HAPH and pathological erythrocytosis (Monge's disease) are relevant problems, with prevalences varying between 15% and 20% (Leon-Velarde et al., 2005).
The vasodilator nitric oxide (NO) plays an important role in the regulation of oxygen delivery by local vasomotor control and in the central cardiovascular and respiratory responses to hypoxia (Semenza, 2011). Circulating NO has been shown to be higher in the Ladakhis, a high-altitude population of the Tibetan plateau, as compared with lowlanders (Ahsan et al., 2006). NO is produced by NO synthase (NOS), of which three isoforms are known (Michel and Feron, 1997). In the vasculature, the endothelial NOS (eNOS) plays a crucial role. It is regulated by several mechanisms and/or endogenous substances, such as methylarginines (Kittel and Maas, 2014).
Methylation of L-arginine residues in proteins (Nicholson et al., 2009) is a mechanism of post-translational protein modification. (Bedford and Richard, 2005). The products of such protein arginine methylation, monomethyl-L-arginine (L-NMMA), asymmetric and symmetric dimethylarginine (ADMA and SDMA), are endogenously present in the human body. ADMA and L-NMMA competitively inhibit NO synthesis from L-arginine, whereas SDMA does not (Vallance et al., 1992). ADMA has been identified as an important regulator of NO production in vivo (Böger et al., 2009). There is little published information on the possible role of ADMA in high altitude-induced hypobaric hypoxia in humans and its association with altitude diseases and acclimatization. Recently, one study in yaks demonstrated a low plasma ADMA concentration as a result of increased expression and activity of dimethylarginine dimethylaminohydrolase (DDAH), the enzyme that degrades ADMA (Mizuno et al., 2015). We have recently shown that ADMA is also increased in rats that are exposed to chronic intermittent hypoxia (Lüneburg et al., 2016). Moreover, a study in humans found that the occurrence of high-altitude pulmonary edema is associated with an enhanced vasoconstrictor activity due to an impaired NO pathway involving ADMA (Ali et al., 2012). However, changes over time of ADMA, L-NMMA, and SDMA, especially in long-term chronic intermittent hypoxia, have not yet been reported. For the first time, we performed a longitudinal study with young adults, non-natives and never exposed to high altitude (3550 m) undergoing CIH shifts, and followed them during a study period of 3 months. To compare different regimens of hypobaric hypoxia, we also included native Andean residents permanently living at high altitude as a group of people exposed to chronic hypobaric hypoxia (CH).
Methods
Study population
Chronic intermittent hypobaric hypoxia (CIH) group
One hundred male army draftees were included in our longitudinal study of CIH. From these participants never exposed to high altitude earlier, 28 were lost at follow-up visits for non-medical, that is, emotional and socioeconomic, reasons. The subjects' mean age was 18.9 ± 0.9 years, and 63% were smokers. Further characteristics describing the CIH group are provided in Table 1.
Values are mean ± SE.
p < 0.01 M3 versus M0.
p < 0.01 CH versus all the CIH periods.
p < 0.01 CH versus CIH M3.
p < 0.001 M3 versus M0.
CH, chronic hypobaric hypoxia; CIH, chronic intermittent hypobaric hypoxia.
Chronic hypobaric hypoxia (CH) group
A group of 16 male Andean natives living in Cariquima and Colchane (Chile; 3675 m) were studied as a chronic hypobaric hypoxia (CH) comparison group. The mean age of this CH group was 43.9 ± 1.2 years, and 19% were smokers. Further characteristics describing the CIH group are provided in Table 1.
Study protocol
CIH protocol
The army draftees were in a CIH shift regimen of 5 days at altitude (3550 m) and 2 days at sea level (SL) during the whole study period of 3 months. All army recruits had initially undergone physical examination elsewhere, the results of which had been considered healthy. Study participants adhered to a controlled diet (3000 kcal daily) and a defined daily exercise regimen (half an hour of mild effort walking at altitude and a 1-hour running exercise at SL) during the whole study period.
Measurements were taken on three defined study days: at baseline (before exposure to high altitude; M0), at month 1 (after 1 month of exposure to CIH; M1), and at month 3 (after 3 months of exposure to CIH; M3). All measurements were performed early in the morning, after 8 hours of fasting. Baseline measurements (M0) were taken at SL, except the Lake Louise Score (LLS), which is a self-assessment test describing the extent of AMS at altitude (Roach et al., 1993). For altitude conditions (M1 and M3), physiological measurements and LLS were taken after one night's sleep on the first day of the corresponding shift, 18 hours from arrival at altitude.
CH protocol
All parameters were measured on one random study day at altitude, early in the morning after 8 hours of fasting. Study participants consumed their usual diet, and most of them were sedentary according to WHO criteria (Bull et al., 2009). Only healthy subjects after a medical assessment were included.
Written informed consent was obtained from all study participants. This study was approved by the Ethical Committee of Universidad Arturo Prat, Iquique, Chile.
Variables measured
Age (years), weight (kg), height (m), and smoking habits were recorded. Body mass index (BMI) was calculated as weight (kg) divided by height (m2). On each study day, systolic blood pressure (SBP, mmHg), diastolic blood pressure (DBP, mmHg), heart rate (HR, beats/min), and hemoglobin oxygen saturation (SaO2, %) were measured. SBP and DBP were measured on the right arm of each participant in the seating position after a 5-minute rest, using appropriately sized cuffs and calibrated standard mercury sphygmomanometers. HR was measured with an HR-100C Omron device (Omron®, Healthcare Inc., Bethesda), and SaO2 was measured by using a pulse oximeter (POX050; Mediaid®, Cerritos). The mean of two measurements separated by a 3-minute interval was taken as the valid determination for BP, HR, and SaO2 according to international guidelines.
At altitude, the LLS was assessed on each of the three study days (Roach et al., 1993); AMS was diagnosed clinically when headache and one or more other symptoms occurred (i.e., gastrointestinal symptoms, insomnia, dizziness, lassitude, or fatigue) and an LLS of ≥3 was achieved. Severity was assessed according to the following score: mild (3–4), moderate (5–10), and severe (11–15) (Roach et al., 1993).
Venous blood samples were taken without blood stasis. At M0, M1, and M3, the following laboratory parameters were measured in the CIH group: hematocrit, hemoglobin, ADMA μmol/L, NNMA μmol/L, and SDMA μmol/L. The same parameters were assessed at one single occasion in the CH group. For the measurement of dimethylarginines, EDTA plasma was kept frozen at −80°C until analysis. Hematocrit and hemoglobin were measured on the day of blood withdrawal by using an automatic hematological counter (Cell-dyn 3700®, Tecnigen, Santiago, Chile).
Measurement of dimethylarginines
Mass spectrometric determination of ADMA, L-NMMA, and SDMA was performed as described elsewhere (Schwedhelm et al., 2005, 2007). In brief, 20 μL aliquots of EDTA plasma were analyzed by using 96-well 0.22-μm microfiltration plates (Multiscreen HTS™, Millipore, Molsheim, France) after addition of 100 μL aliquots of a methanolic solution of 40 pmol [2H6]-ADMA (Santa Cruz, Dallas). After conversion to their butyl ester derivatives, analytes were quantified with a Varian 1200L Triple Quadrupole MS (Agilent Technologies, Santa Clara) in the positive electrospray ionization (ESI+) mode. For all biomarkers, the imprecisions for inter- and intra-assay variations were <5.6% and <5.4%, respectively.
Data analysis
Data are given as mean ± SE or %, unless indicated otherwise. All variables except LLS were normally distributed (Kolmogorov–Smirnov test). Statistical differences were tested by analysis of variance (ANOVA) of repeated measures in the follow-up CIH group or by Student's t-test, except for not normally distributed variables, for which the Wilcoxon test was used. The chi-square test was used for the observation of proportional differences for independent samples, and the McNemar test was used for related samples. Pearson's correlation analyses were performed between quantitative variables. A combined variable “Poorly Acclimatized” was created for subjects who had altitude outcomes, AMS presence, plus SaO2 <89% at month 3. Subsequently, a receiver operating characteristic (ROC) curve analysis was performed to test the ability of ADMA to discriminate and correctly classify, by the area under the curve (AUC), those poorly acclimatized, with its confidence intervals and sensitivity and specificity. A significance level was established at p < 0.05. Data were analyzed by using the SPSS 17.0 statistical package (SPSS, Inc.®, Chicago, IL).
Results
Time course of physiological variables in CIH
We observed a slight reduction of SaO2 over time (p < 0.001). Hematocrit and hemoglobin increased from M0 to M1 and were maintained until M3 (p < 0.001) (Table 1). SBP was increased at M3 compared with baseline (p < 0.01, Table 1), whereas DBP did not change. HR continuously increased over time at high altitude (p < 0.001).
AMS assessment
The mean LLS at M0 was 2.7 ± 0.3. The LLS as well as the incidence of AMS decreased over time (p < 0.01 for both, Table 2), and no severe cases of AMS were observed in M3. No correlation with other variables was found.
Values for LLS are mean ± SE.
Differences among periods versus M0 p < 0.01.
AMS, acute mountain sickness; LLS, Lake Louise score.
Altitude native residents (CH group)
Mean SaO2 was slightly, but significantly lower (90.3% ± 0.3%; p < 0.01) than CIH M3. Hematocrit and hemoglobin were higher compared with CIH on all study days (p < 0.001, Table 1). Systolic and DBP in CH were comparable to CIH at M0 and slightly lower than CIH at M3 (p < 0.01, Table 1). HR was lower in CH than in CIH at M3 (p < 0.01).
Dimethylarginines
Plasma concentrations of both ADMA and L-NMMA were significantly elevated in CIH subjects at M1 and M3 compared with baseline (Fig. 1). ADMA progressively increased, reaching 80% higher levels at M3 (1.11 ± 0.04, 1.26 ± 0.05, 1.95 ± 0.09 μmol/L; Fig. 1A). Also, with prolonged exposure time, an increasing proportion of subjects with ADMA levels that were higher than 2.0 μmol/L was observed (M0: 2%; M1: 7%; M3: 38% of study participants had ADMA levels higher than 2.0 μmol/L). Likewise, L-NMMA increased by 60% at M3 (0.22 ± 0.08; 0.24 ± 0.03; 0.39 ± 0.03 μmol/L; Fig. 1B), whereas SDMA concentrations remained unchanged (0.45 ± 0.02 μmol/l vs. 0.50 ± 0.05 μmol/L; p = NS) (Fig. 1C). In the CH group, ADMA and L-NMMA plasma concentrations were twofold higher compared with CIH at M0 (Fig. 1A, B). SDMA was only slightly higher in CH compared with CIH at M0 and M1, with no significant difference between CH and CIH at M3 (Fig. 1C).
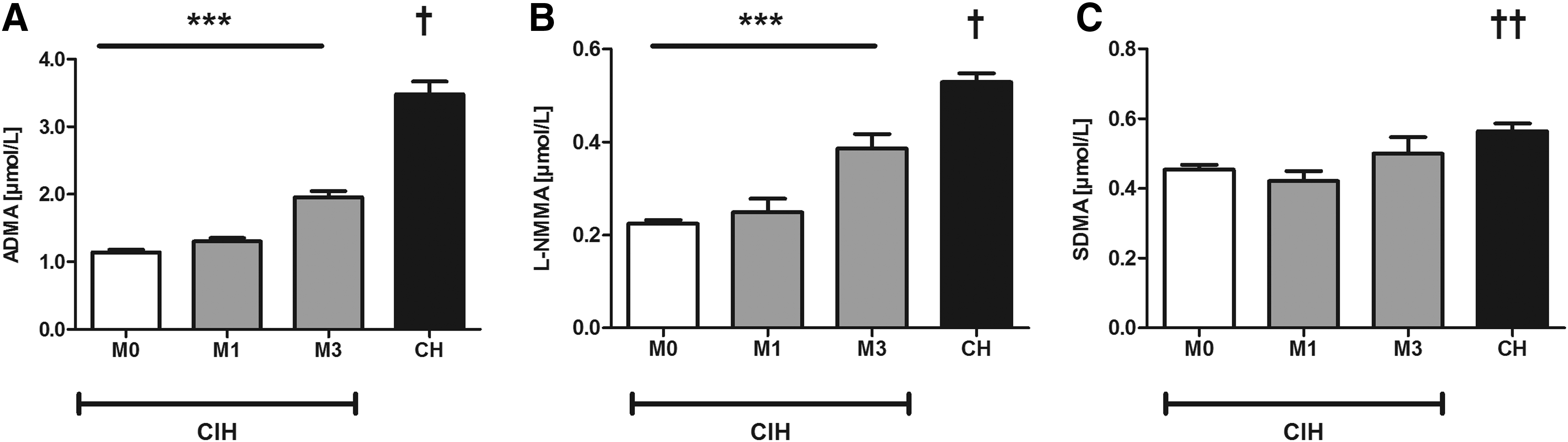
Plasma concentrations of dimethylarginines in the CIH group (n = 72) at month 0 (M0), month 1 (M1), and month 3 (M3), and in the CH group (n = 16);
In CIH, there was a correlation between ADMA and hematocrit at altitude M3 (R = 0.28, p < 0.05) (Fig. 2A). In the CH group, there was a strong correlation between ADMA and hematocrit (R = 0.66; p < 0.01) (Fig. 2B). A positive correlation between hematocrit and SBP was found in both groups: CIH at M1 (R = 0.34, p < 0.001); CH (R = 0.51; p < 0.01). Neither ADMA nor SDMA nor L-NMMA plasma levels correlated significantly with SBP, DBP, or SaO2.
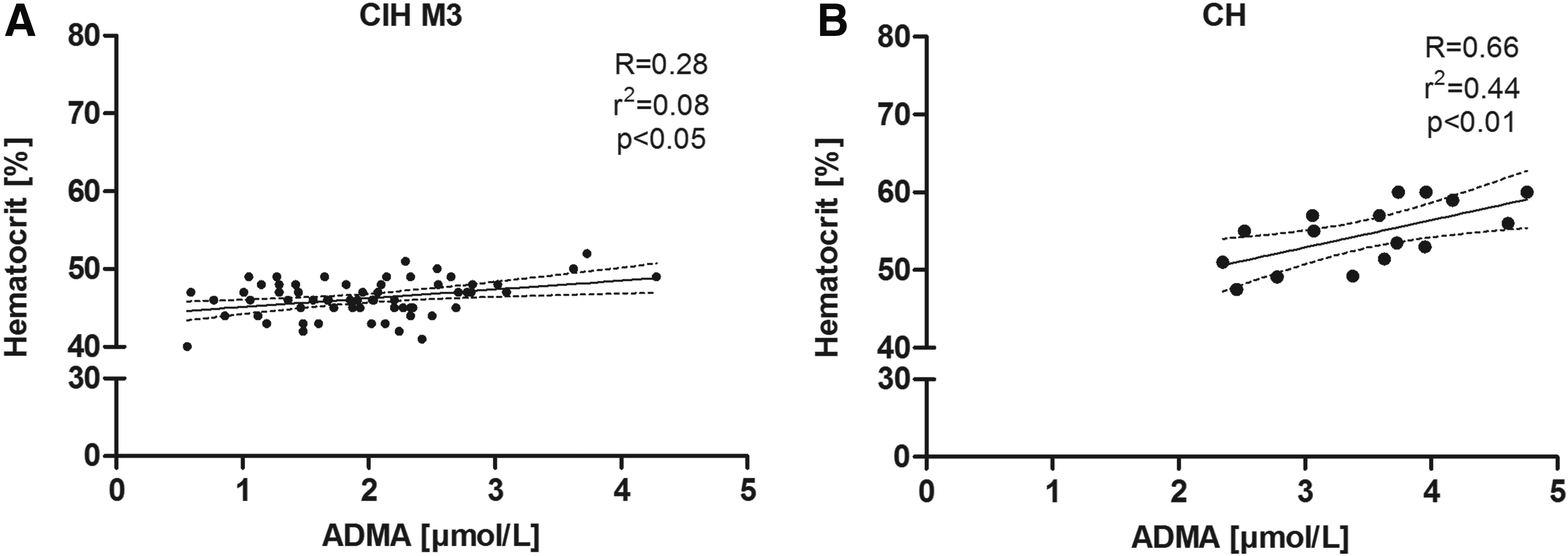
Pearson's correlation analyses of ADMA (μmol/L) and hematocrit (%) of
At M3, ROC curve analysis (Fig. 3A) showed a strong association between ADMA concentration and poor acclimatization status (AUC 0.78, 95% CI 0.60/0.95, p < 0.05, Fig. 3B) showing that having ADMA >2.7 μmol/L could be predictive of poor acclimatization.
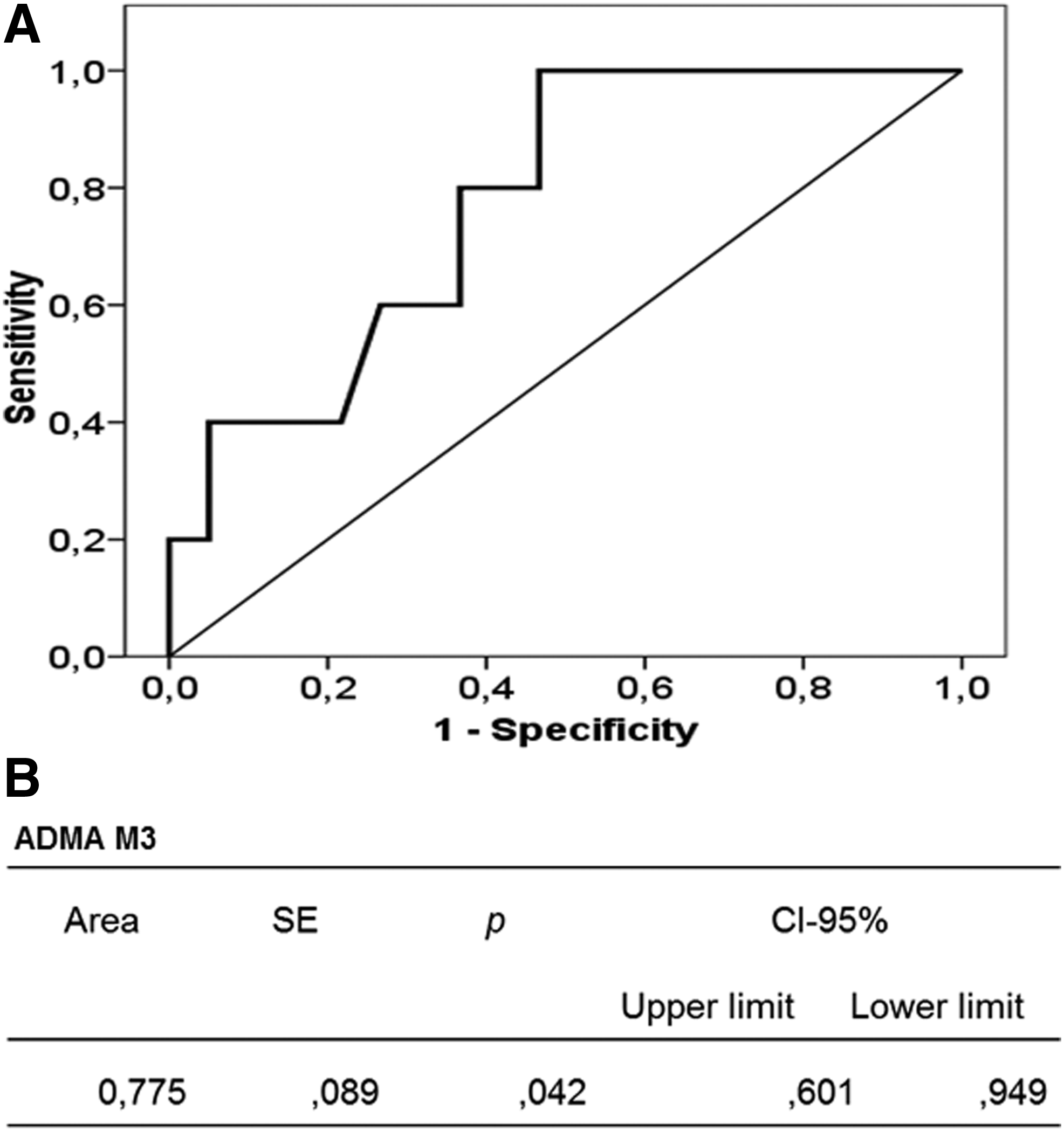
ROC curve
Discussion
Our study has three major findings: (1) Three months of exposure to CIH causes a significant elevation of ADMA and L-NMMA plasma concentrations, whereas SDMA levels do not change significantly. Likewise, Andean native residents at high altitude have very high ADMA levels. (2) ADMA plasma concentration is positively correlated with hematocrit in CIH as well as CH. (3) In CIH, high ADMA concentration is related to poor acclimatization status.
For the first time, our study shows that altitude-associated hypoxia selectively increases the plasma concentrations of ADMA and L-NMMA, but not SDMA. High ADMA concentration is a strong independent predictor of mortality in patients with idiopathic pulmonary arterial hypertension (Kielstein et al., 2005). Increased ADMA and L-NMMA concentrations are associated with pulmonary hypertension (PH) in patients with sickle cell disease (Kato et al., 2009). Reduced bioavailability of NO under pathophysiological conditions may be caused by decreased expression of eNOS, inhibition of eNOS activity, or early oxidative degradation of NO by superoxide radicals (Millatt et al., 2003). As both ADMA and L-NMMA competitively inhibit eNOS, the studies cited earlier (Kielstein et al., 2005; Kato et al., 2009) suggest that impaired production of NO may be a major cause of endothelial dysfunction in PH and may contribute to the progression of this disease. Our study adds important information by demonstrating that high altitude significantly elevates ADMA and L-NMMA, but not SDMA. This effect was independent of the regimen of exposure (CIH or CH). Interestingly, mean ADMA plasma concentration was even higher in Andean natives who had spent their whole lives at high altitude than in the CIH subjects during their relatively short exposure during 3 months, although one might expect a higher level of adaptation in high-altitude residents. It might be interesting to know whether ADMA concentration is also high in other high-altitude populations, for example, Tibetan natives, in whom different adaptation mechanisms have been reported (Beall, 2006).
The results of our present clinical study are in line with our previous study in rats, in which we showed that ADMA, but not SDMA, is elevated in experimental CIH and CH (Lüneburg et al., 2016).
The main difference in the metabolism of ADMA, SDMA, and L-NMMA is that ADMA and L-NMMA are enzymatically cleared by dimethylarginine dimethylaminohydrolase (DDAH), whereas SDMA is not a DDAH substrate (Tran et al., 2003). Rather, SDMA is cleared by the enzyme AGXT2 (Lüneburg et al., 2014) and by renal excretion.
In our previous study in rats, we found decreased expression and activity of DDAH in hypoxia, which explains the selective elevation of the DDAH substrates, ADMA and L-NMMA. The pattern of changes in biomarkers measured in the study presented here (elevation of ADMA and L-NMMA, no significant change in SDMA) suggests that dysregulation of DDAH may also occur in high altitude-induced hypoxia in humans. This hypothesis is supported by the study of Mizuno et al. (2015), who demonstrated reduced ADMA and enhanced DDAH activity in lungs of yaks—animals adapted to high altitude—as opposed to bovine. Furthermore, Ali et al. (2012) recently reported that elevation of ADMA, among other biomarkers, was associated with high-altitude pulmonary edema in first-time high-altitude sojourners. Although these authors measured ADMA at one single time point after first-time exposure to high altitude, we report here the time course of ADMA, NMMA, and SDMA plasma concentrations during 3 months of repeated exposure to intermittent hypoxia. In addition, Ali et al. (2012) reported a mean ADMA concentration of 1.9 ± 0.8 μmol/L in Tibetan high-altitude native residents. By contrast, mean ADMA concentration in the Andean high-altitude residents was as high as 3.5 ± 0.2 μmol/L, suggesting the possibility of different molecular adaptation mechanisms between different populations living at high altitude. This interesting hypothesis needs to be corroborated in future studies.
The association between ADMA and hematocrit at CIH M3 and CH, as found in our study, supports our hypothesis that the elevation of ADMA relates to hypoxia. An increase in hematocrit and a subsequent higher blood viscosity are expected phenomena that are related to hypoxia at high altitude (Zubieta-Calleja et al., 2007). Hematocrit has been demonstrated to be predictive of adaptation to altitude hypoxia (Reeves and Leon-Velarde, 2004; Siqués et al., 2006). However, this phenomenon is restricted to Andean populations. Other highland populations such as Tibetans or populations of Ethiopia do not show an increase in hematocrit at a similar altitude (Beall, 2006).
As opposed to acute hypoxia, where decreased systemic blood pressure is observed as a result of acute hypoxia-induced NO production in the endothelium (Cabel et al., 1994), we noted a tendency toward higher SBP in CIH. This observation is in line with the response to CIH caused by obstructive sleep apnea, where vasodilation by NO is overcome by other vasoconstrictor molecules such as endothelin-1 (Fletcher, 2001). An increase in blood pressure in individuals under chronic hypoxia at high altitude has repeatedly been demonstrated (Calbet, 2003; Siqués et al., 2006; Luks, 2009). Our results are consistent with these published data.
Insufficient adaptation to acute exposure to high altitude and persisting low arterial oxygen saturation are two major determinants of altitude-associated morbidity (Hackett and Roach, 2001; Richalet et al., 2002; Brito et al., 2007). We, therefore, defined “poor acclimatization” as the combination of persisting low SaO2 and the presence of AMS. For the first time, we describe here a tight association between high ADMA and “poor acclimatization,” pointing to a predictive role of ADMA as a biomarker for altitude-associated morbidity. This finding might help to identify individuals who are susceptible to poor acclimatization based on their basal ADMA plasma concentration before their first exposure to altitude. The ADMA/DDAH pathway is tightly controlled, because it plays a crucial role in regulating NO bioavailability. Dysregulation of the ADMA/DDAH pathway has been reported in different hypoxia-associated chronic respiratory diseases such as asthma (Scott et al., 2011), chronic obstructive pulmonary disease (Scott et al., 2014), and obstructive sleep apnea syndrome (Barceló et al., 2009). We, therefore, hypothesize that downregulation of DDAH expression and/or activity, resulting in elevated ADMA concentration and subsequent loss of NO-dependent vasodilation, contributes to pulmonary ventilation/perfusion matching. When hypoxia becomes global instead of regional, dysregulation of the ADMA/DDAH pathway elicits systemic symptoms (e.g., elevated pulmonary or systemic blood pressure), thereby contributing to the progression of disease. Future prospective studies are needed to validate this hypothesis.
Finally, to reverse the altitude-associated dysregulation of the ADMA/DDAH-pathway, L-arginine supplementation or pharmaceutical upregulation of DDAH activity appears to be a promising option. Animal research strongly supports the use of L-arginine. In humans, pre-treatment with intravenous L-arginine decreases AMS score and improves arterial oxygen saturation at 4350 m (Schneider et al., 2001). In addition, L-arginine is able to improve exercise efficiency and tolerance during high-intensity exercise (Bailey et al., 2010). Finally, it elicits an improved short-term pulmonary vasodilatory response in patients with PH of various origins (Mehta et al., 1995). Since there is currently no therapeutically useable DDAH activator available, L-arginine supplementation seems to be the only therapeutic option at present.
Limitations
Our study included only male study participants. It is known that premenopausal women have lower ADMA plasma concentrations than age-matched men (Schulze et al., 2005; Schwedhelm et al., 2009). Therefore, it is unclear whether our results can be extended to women. Another limitation is the small number of native altitude residents. However, although blood samples were drawn on one random occasion in this control group, variability of all variables was low. We, therefore, believe that these data are meaningful. As we previously demonstrated that ADMA increases only slightly with age in men and women (Schulze et al., 2005; Schwedhelm et al., 2009), we propose that the large difference in ADMA concentration between CIH and CH may be, in major part, related to the duration of exposure to high altitude. The significant difference in BMI between CIH and CH groups may also have contributed to differences in ADMA plasma concentrations. We previously reported associations of ADMA and BMI in obese women with a median BMI of 28 (Reimann et al., 2007). Since the CH group had a mean BMI of 25, it cannot be classified as obese. In addition, it must be considered that the proportion of smokers in both cohorts, especially in the CIH, is very high. Smoking impairs endothelial function; however, it tends to lower ADMA concentration, as previously demonstrated by our group (Maas et al., 2007). A combined effect of ADMA and smoking on endothelial function can, therefore, not be ruled out.
Conclusion
Chronic intermittent hypobaric hypoxia (CIH) and chronic hypobaric hypoxia (CH) lead to a large increase in ADMA and L-NMMA plasma concentrations, which may result in impaired NO availability. ADMA may play a dual, important role as (1) a predictor of poor acclimatization to hypoxia, and (2) a pathophysiological mediator involved in the incidence and progression of CIH-induced hypertension, AMS, and HAPH. As ADMA competes with L-arginine, the natural substrate of NO synthase, supplementation with L-arginine, as mainly demonstrated in animal experiments, may be one possibility to prevent diseases that are associated with high altitude.
Footnotes
Acknowledgments
The authors thank Mariola Kastner, Anna Steenpass, Sandra Maak, Stefany Ordenes, and Gabriela Lamas for their excellent technical assistance. This work was supported by grants from the FIC GORE-TARAPACA BIP 30434827-0, University Arturo Prat, and ALTMEDFIS-CYTED.
Author Disclosure Statement
No competing financial interests exist.