
Abstract
Abstract
Omrani, Hasan, Mohammad Reza Alipour, Fereshteh Farajdokht, Hadi Ebrahimi, Mehran Mesgari Abbasi, and Gisou Mohaddes. Effects of chronic ghrelin treatment on hypoxia-induced brain oxidative stress and inflammation in a rat normobaric chronic hypoxia model. High Alt Med Biol. 18:145–151, 2017.
Aim:
This study aimed to evaluate the probable antioxidant effects of ghrelin in the brain and serum and its effect on tumor necrosis factor-alpha (TNF-α) levels in the brain in a model of chronic systemic hypoxia in rats.
Methods:
Systemic hypoxia was induced by a normobaric hypoxic chamber (O2 11%) for ten days. Adult male Wistar rats were divided into control (C), chronic ghrelin (80 μg/kg/10 days) (Ghr), chronic hypoxia (CH), and CH and ghrelin (80 μg/kg/ip/10 days) (CH + Gh) groups. The activity of superoxide dismutase (SOD), glutathione peroxidase (GPx), catalase (CAT), and malondialdehyde (MDA), total antioxidant capacity, and TNF-α levels were assessed in the serum and brain tissue.
Results:
Our results showed that chronic ghrelin administration attenuated the CH-increased oxidative stress by decreasing MDA levels in the serum and brain tissue. Moreover, ghrelin enhanced the antioxidant defense against hypoxia-induced oxidative stress in the serum and brain tissue. Brain TNF-α levels in CH did not change significantly; however, ghrelin significantly (p < 0.001) decreased it.
Conclusion:
These results indicated that ghrelin promoted antioxidative and anti-inflammatory defense under chronic exposure to hypoxia. Therefore, ghrelin might be used as a potential therapy in normobaric hypoxia and oxidative stress induced by CH.
Introduction
H
Previous studies found that hypoxia results in an increased mitochondrial reactive oxygen species (ROS) generation and reduction of antioxidant defenses (Sabharwal and Schumacker, 2014). ROS can interact with biomolecules and damage various cellular components such as DNA, RNA, proteins, and lipids (Turrens, 2003; Erguder et al., 2005; Pajocvic and Saicić, 2008).
Combination of several factors makes the CNS particularly vulnerable to oxidative damage (Dringen, 2000), including the low-to-moderate activity of catalase and glutathione peroxidase (GPx), having high levels of unsaturated fatty acids (Cooper and Kristal, 1997), and high metabolic activity and oxygen consumption (Porter, 1984). Enzymatic antioxidant defenses including superoxide dismutase (SOD), thioredoxin, GPx, glutathione reductase, and catalase (CAT) are involved in the protection of cell components from oxidative stress and damage (Gemma et al., 2007; Valko et al., 2007).
On the other hand, the effect of hypoxia on the brain is intensified by the production of inflammatory cytokines from glia and neuronal cells (Mukandala et al., 2016). Hypoxia induces production of inflammatory cytokines such as interleukin 1 alpha (Shreeniwas et al., 1992), tumor necrosis factor-alpha (TNF-α), and nuclear factor kappa B, which can result in systemic inflammation and cell death (Kalpana et al., 2008). Several studies support a role for TNF-α in disruption of the blood–brain barrier, edema formation in cerebral ischemia (Yang et al., 1999), and brain trauma (Shohami et al., 1997b). TNF-alpha aggravates inflammation by stimulation of capillary endothelial cell proinflammatory responses and increases leukocyte adhesion and infiltration into the ischemic brain (Feuerstein et al., 1994). Agents that suppress TNF-alpha production or action reduce leukocyte infiltration into ischemic brain regions, and thereby diminish the extent of tissue loss (Feuerstein et al., 1994) and brain injury during transient cerebral ischemia (Yang et al., 1998).
Ghrelin is a 28-amino acid hormone that is principally released from the stomach (Date et al., 2000). Ghrelin has several functions in the CNS, including energy homeostasis (Tschöp et al., 2000), neuroprotection, learning and memory, reward and motivation (Andrews, 2011), and antinociceptive and antiphotophobia effects (Farajdokht et al., 2016, 2017). Also, anti-inflammatory and antiapoptotic properties of ghrelin have been proved (Bansal et al., 2010; Cheyuo et al., 2011; Azizzadeh et al., 2017). Ghrelin passes into the blood–brain barrier (BBB) following systemic administration (Banks et al., 2002) and exerts neuroprotective effects following cerebral ischemia and reperfusion by decreasing neuronal apoptosis, and reducing total infarct volume after the ischemic insult (Liu et al., 2006; Miao et al., 2007). Ghrelin also showed antioxidant properties in testis tissue of adult rats by decreasing levels of malonyldealdehyde (MDA) and increasing glutathione peroxidase (GSH-Px) (Kheradmand et al., 2009). Obay et al. also showed that ghrelin prevented reduction of antioxidant enzyme activities and lipid peroxidation induced by pentylenetetrazole in the erythrocytes, liver, and brain tissue of rats (Obay et al., 2008).
Therefore, the aim of this study was to evaluate the effect of chronic ghrelin treatment on lipid peroxidation levels (MDA), antioxidant enzyme activities (catalase, SOD, and GPx), and inflammatory cytokine (TNF-a) following chronic normobaric systemic hypoxia in rats.
Materials and Methods
Animals
Adult male Wistar rats (220–250 g) were housed in standard cages in a temperature-controlled room (22°C–24°C), with humidity of 40%–60% and a light period of 12-hour dark–12-hour light. Food and water were available ad libitum. All experiments were performed in agreement with guidelines of the Tabriz University of Medical Sciences for care and use of laboratory animals.
Experimental groups
After a week of habituation, animals (N = 28) were randomly divided into four (n = 8) experimental groups:
C: control
Ghr: Ghrelin (80 μg/kg/day for 10 days)
CH: chronic hypoxia
CH + Ghr: chronic hypoxia that received ghrelin for 10 days
Control animals received intraperitoneal (i.p) injection of saline (1 mL/kg/day). Ghrelin (Innovagen, Sweden)-treated groups received ghrelin with a dose of (80 μg/kg/i.p) for ten days (Hossienzadeh et al., 2013) in the same volume.
Hypoxia induction (normobaric)
Animals in chronic hypoxia were placed into the hypoxia chamber (GO2 Altitude-Biomedical Australia) (10.5%–11% O2) for 10 days and only emitted for 10 min in a day to clean the cages and receive injections (Schwenke et al., 2008; Bauer et al., 2010). The control group was placed in room air (21% O2) in the same room.
Sampling
Tail blood samples were used for blood glucose measurement using a commercial glucometer (Accu-chek Active, Roche, Germany). On the last day of experiments, animals were deeply anesthetized with a combination of 80 mg/kg ketamine and 12 mg/kg xylazine, blood samples (5 mL) were collected from the heart and centrifuged at 4000 rpm for 10 min, and serum aliquots were isolated. Then, animals were immediately sacrificed by decapitation and brain tissues were carefully removed. All samples were kept in −80°C for later analysis.
Determination of brain antioxidant enzyme activity (GPX, SOD, CAT), MDA and TNF-α levels
Brain samples were washed with ice-cold 0.9% NaCl to remove the blood, then homogenized in a lysis buffer containing a complete protease inhibitor, and centrifuged for acquiring the supernatant. The resulting supernatants were used for enzyme activity analysis. The GPx activity was measured according to Paglia and Valentine (Paglia and Valentine, 1967) using Randox (United Kingdom). Tissue SOD was evaluated by a spectrophotometric method based on the inhibition of a superoxide-induced reduced nicotinamide adenine dinucleotide oxidation (Paoletti and Mocali, 1990). The catalase activity was measured based on the change in absorbance at 240 nm at 25°C of the solution of 10 mM H2O2 in phosphate buffer, pH 7.0 (Luck, 1965).
The activities of GPx and SOD were expressed as IU/mg protein in tissue homogenate, respectively.
Malondialdehyde (MDA) levels in the brain were measured using the thiobarbituric acid reactive substances (TBARS) method (Kaya et al., 2004).
Brain TNF-α level was measured using a rat TNF-α enzyme-linked immunosorbent assay (ELISA) kit (Invitrogen). The assay was carried out according to the instruction provided by the manufacturer.
Determination of serum MDA and total antioxidant capacity
Malondialdehyde level was determined in the serum of animals using the thiobarbituric acid (TBARS) method. Results were obtained in terms of nmol/mL and determined using the colorimetric method. The total antioxidant capacity (TAC) in the serum was measured using a rat-specific ELISA Kit according to the manufacturer's protocols (Miller et al., 1997).
Statistical analysis
All results are expressed as the mean ± standard error of mean. SPSS 16.0 software was used for statistical analysis of data. Statistical significance difference was determined for the brain MDA, GPx, SOD, and CAT using the Kruskal–Wallis test for multiple comparisons, followed by the post hoc Mann–Whitney test. For other comparisons, one-way analysis of ANOVA followed by Tukey's test was used. The significant level was set at p < 0.05.
Results
Chronic ghrelin treatment decreased brain and serum levels of MDA induced by chronic normobaric hypoxia
The results showed that chronic hypoxia insignificantly increased brain MDA levels (Fig. 1A) and significantly increased serum (p < 0.05, Fig. 1B) MDA levels in comparison with the control group. Moreover, chronic treatment with ghrelin during hypoxia significantly decreased brain (p < 0.05) and serum (p < 0.001) levels of MDA, when compared to the chronic hypoxia (CH) group.
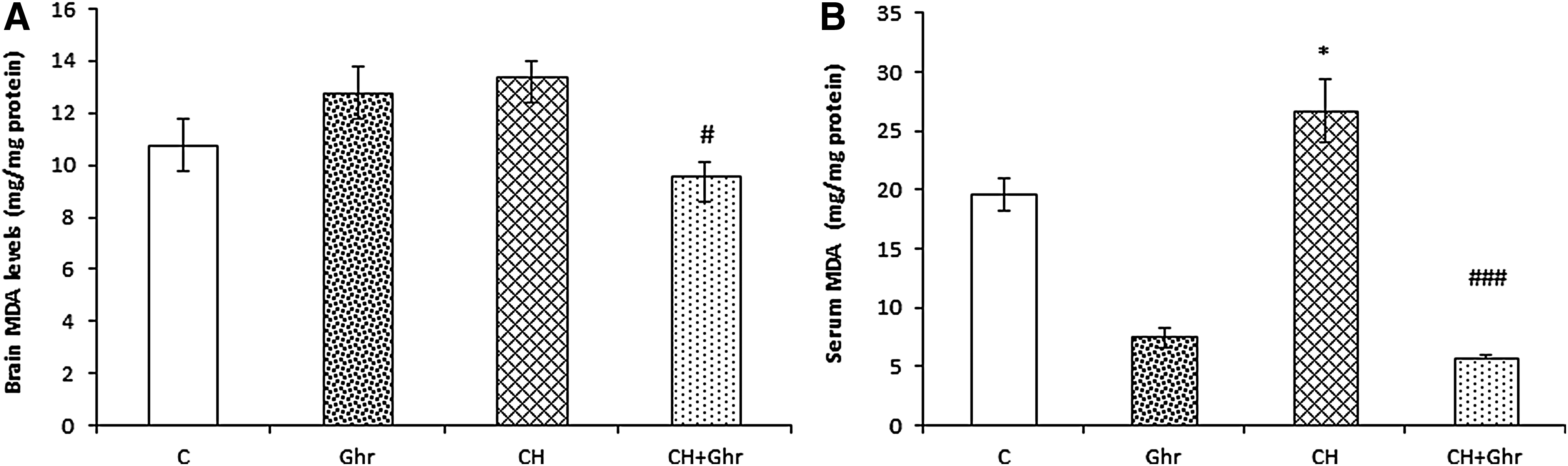
Brain
Chronic ghrelin treatment enhanced the antioxidant defense against hypoxia-induced oxidative stress in the brain
The results of Kruskal–Wallis analysis revealed that the SOD activity significantly (p < 0.05, Fig. 2A) increased in the CH group. Also, a significant (p < 0.05) elevation was observed in the SOD levels of CH + Ghr group when compared with the CH group. GPx levels, another indicator of antioxidant defense, presented a significant (p < 0.05, Fig. 2B) increase in the CH group compared to the control group. In addition, chronic administration of ghrelin significantly (p < 0.05) increased brain GPx levels in the CH + Ghr group compared with the CH group. The catalase activity of the brain in the CH group decreased insignificantly (p = 0.18) (Fig. 2C) compared with the control group. The mean of catalase activity levels in the ghrelin-treated rats was more than the CH group; however, the differences were not statistically significant.
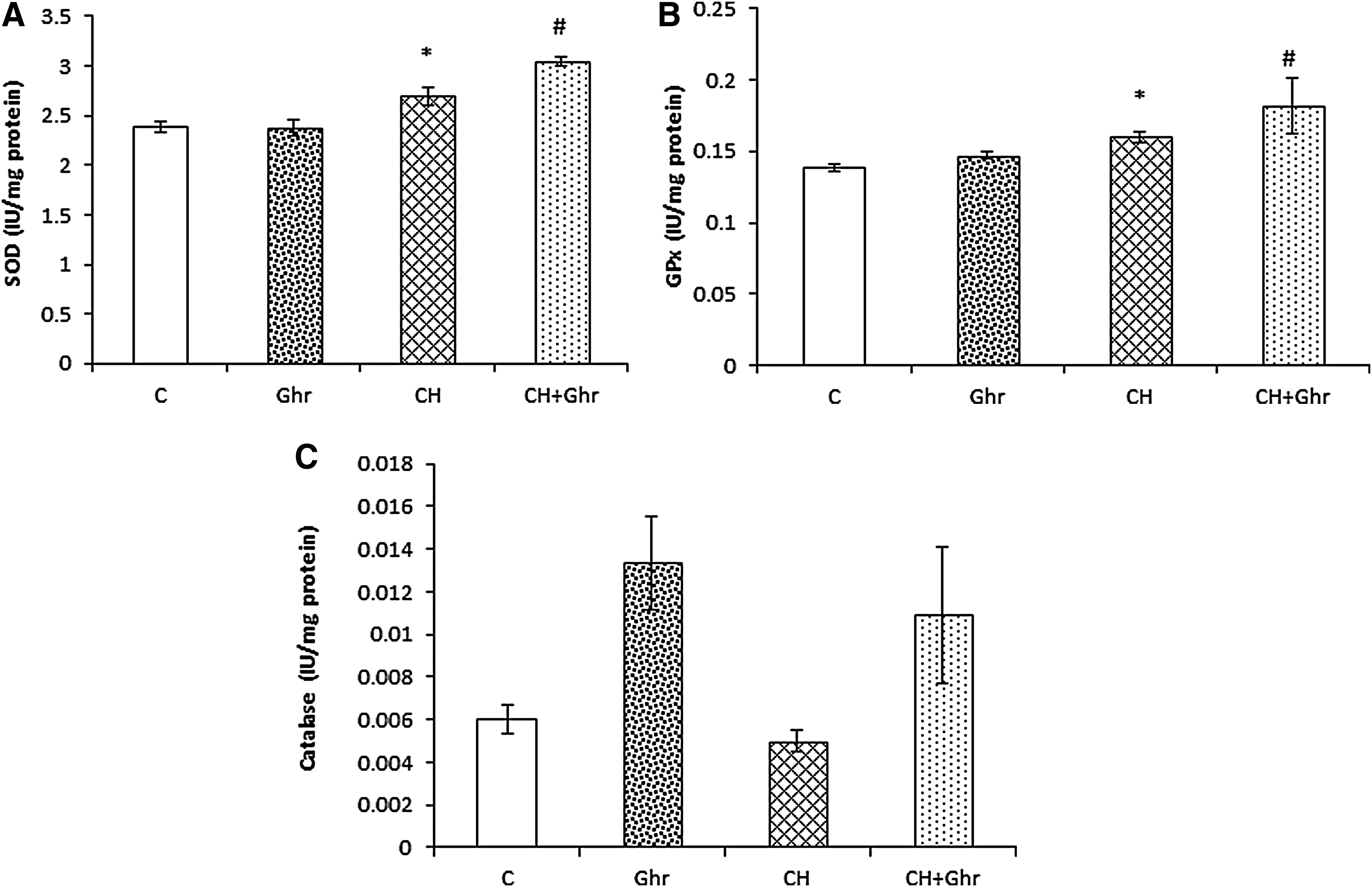
Brain levels of SOD
Chronic ghrelin treatment increased serum TAC
As Figure 3 shows, chronic hypoxia failed to change TAC levels in the serums when compared to the control group. However, mean TAC levels in the CH + Ghr group were significantly (p < 0.001) higher than the chronic hypoxia group.
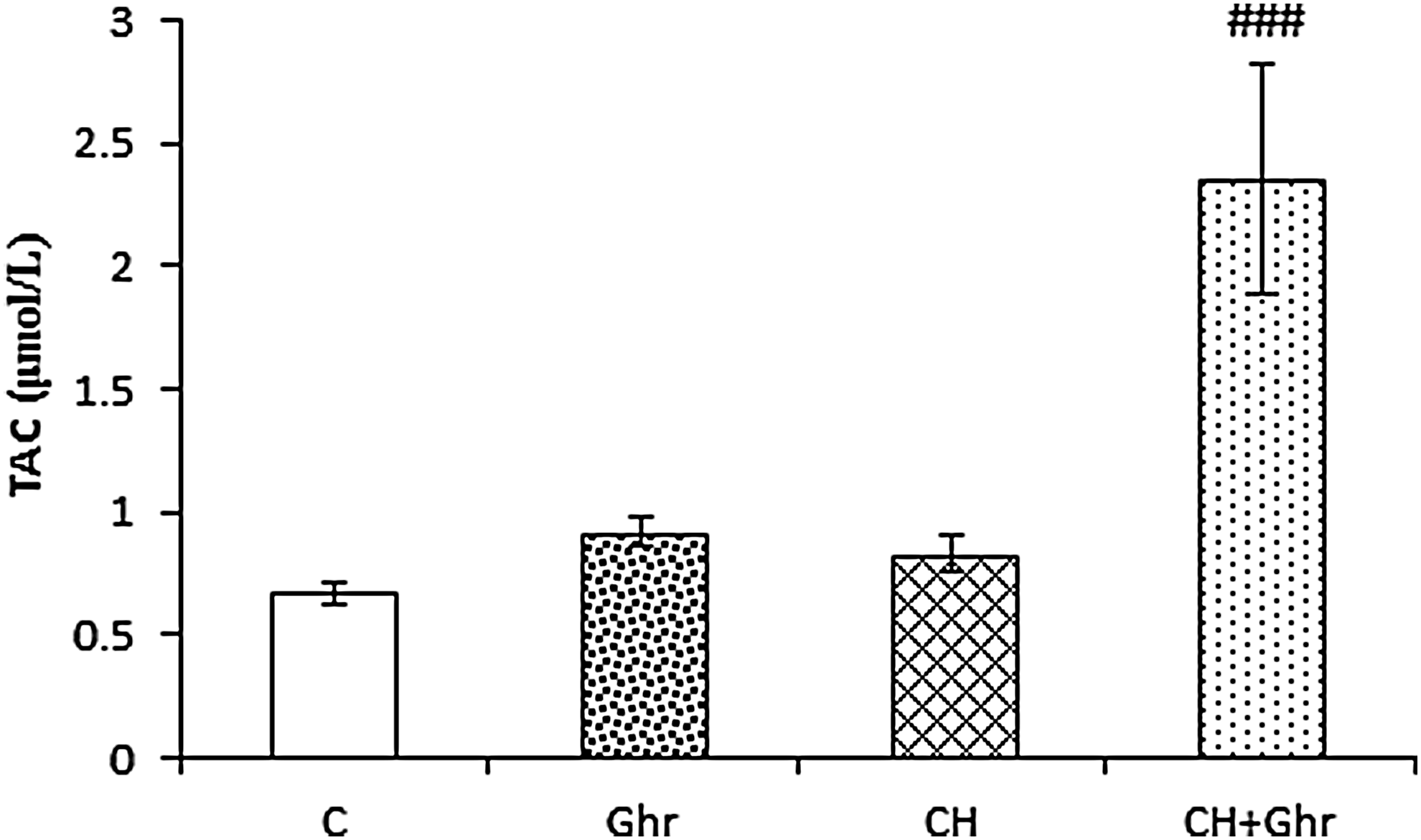
Serum total antioxidant activity (TAC) levels in the control (C), Ghr (Ghrelin), and chronic hypoxia groups that received saline (CH) or 80 μg/kg ghrelin (CH + Ghr) for 10 days. Data are expressed as mean ± SEM for n = 8 animals per group. ###p < 0.001 versus the CH group.
Chronic ghrelin treatment reduced brain TNF-α levels induced by chronic normobaric hypoxia
The results of one-way ANOVA revealed that chronic hypoxia did not change the brain TNF-α levels; however, chronic ghrelin treatment significantly attenuated TNF-α levels (p < 0.001, Fig. 4) compared with the CH group.
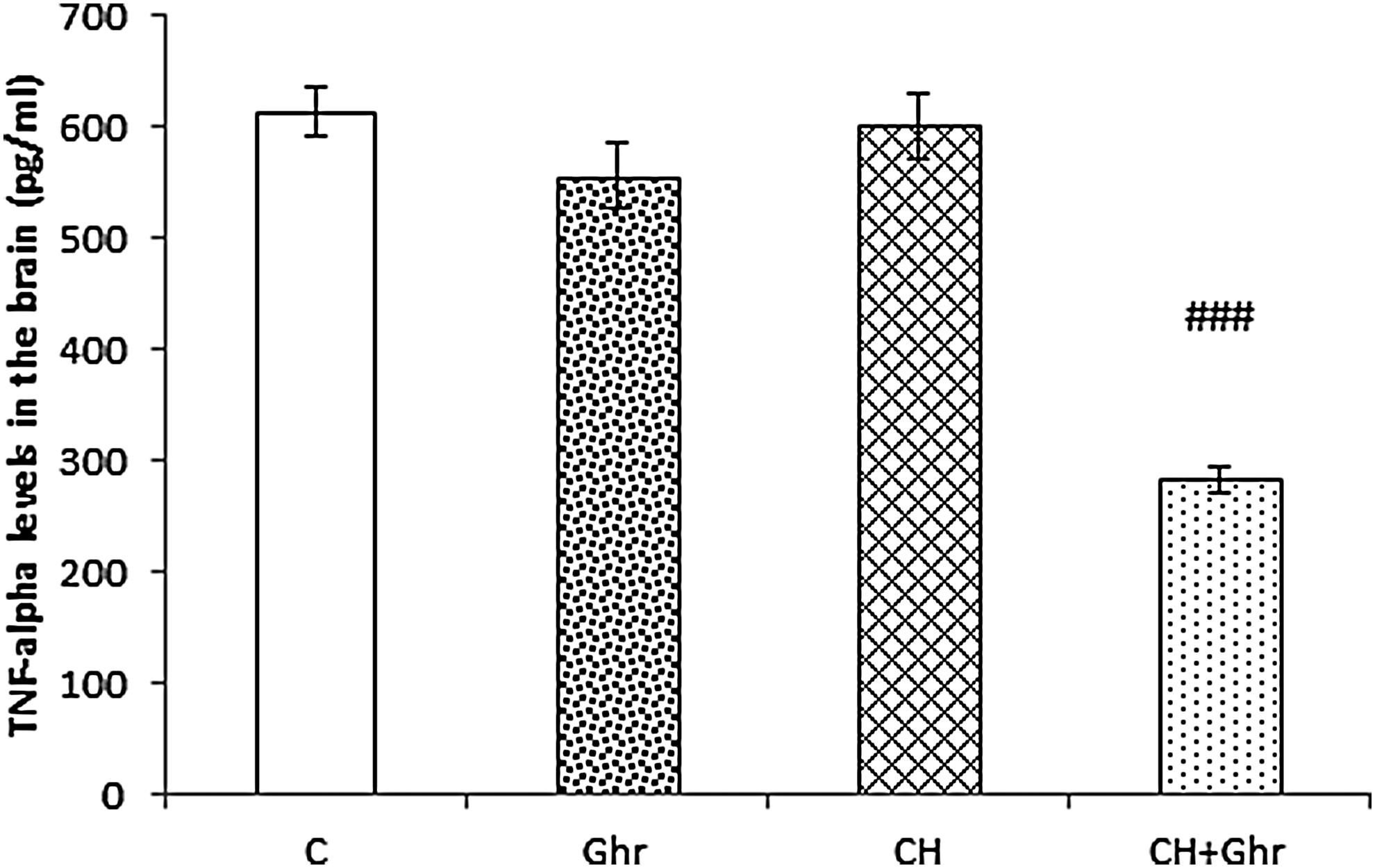
Brain TNF-α levels in control (C), Ghr (Ghrelin), and chronic hypoxia groups that received saline (CH) or 80 μg/kg ghrelin (CH + Ghr) for 10 days. Data are expressed as mean ± SEM for n = 8 animals per group. ###p < 0.001 versus the CH group. TNF-α, tumor necrosis factor-alpha.
Discussion
Our findings demonstrated that the induction of chronic normobaric hypoxia increased MDA levels both in the serum and brain tissue. However, chronic treatment with ghrelin (80 μg/kg/day) for 10 days in chronic hypoxic conditions significantly decreased serum and brain MDA levels, and increased the enzymatic antioxidant activity in the brain (SOD and GPx) and serum TAC levels. Moreover, ghrelin showed an anti-inflammatory property and decreased brain TNF-α level in chronic hypoxia.
In the normal brain, there is a balance between the generation and detoxification of ROS. Failures of balance between formation and removal of free radicals contribute to oxidative stress and brain injury (Pham-Huy et al., 2008; Rahal et al., 2014; Bhattacharya, 2015). Previous studies have shown that intermittent hypoxia is associated with increased oxidative stress (Rahal et al., 2014). Our recent study also revealed that acute hypoxia increased brain and serum levels of MDA in rats (Omrani et al., 2015). It has been shown that hypoxia induces ROS formation, which leads to oxidative stress and lipid peroxidation. Malondialdehyde (MDA) is the end product of lipid peroxidation (Zwirska-Korczala et al., 2007). Previous studies found that an increase in oxidative stress is associated with an antioxidant response (Aydin et al., 2010). Studies have also demonstrated that the intensity of oxidative stress is related to the duration of exposure and the severity of hypoxia (Solaini et al., 2010).
In this study, measurement of the product of lipid peroxidation demonstrated that chronic hypoxia increased the brain and serum levels of MDA. Moreover, administration of ghrelin for ten days effectively decreased MDA levels both in the brain and serum, which indicates decreased lipid peroxidation and reduced oxidative stress damage. The antioxidant and anti-inflammatory effects of ghrelin have been revealed before (Brzozowski et al., 2004; Dixit et al., 2004). Hypoxia induces an increase in mitochondrial ROS formation, which leads to an increase in serum MDA concentrations (Zwirska-Korczala et al., 2007; Sabharwal and Schumacker, 2014). Previously, we found that the administration of ghrelin decreased MDA levels in the brain and serum and increased TAC of serum in acute normobaric hypoxia (Omrani et al., 2015). Can et al. (2015) also revealed that the administration of ghrelin decreased MDA levels in an experimental glaucoma model (Can et al., 2015). In another study, Zwirska-Korczala et al. found that ghrelin significantly decreases MDA concentration in preadipocyte cell culture (Zwirska-Korczala et al., 2007).
In this study, chronic treatment with ghrelin failed to change the activity of catalase in the brain, which seems to be possibly due to low activity of catalase in the rat's brain (Nakano, 1990; Shohami et al., 1997a). Also, animals exposed to chronic hypoxia showed enhanced SOD and GPx activities, which represent an adaptive mechanism against oxidative stress (Gemma et al., 2007; Valko et al., 2007). SOD, a key enzyme in the protective mechanism against free radicals, catalyzes the conversion of O2_ to H2O2 (Fridovich, 1995). Hydrogen peroxide is further detoxified by catalase and/or GPx (Betteridge, 2000). GPx has an important role in the brain antioxidant defense system, although its activity in the rat brain is lower than other organs (Hayes et al., 1989; Galkina, 2013). Taken together, the elevation of SOD and GPx activities in this study seems to be due to enhanced generation of superoxide anion and hydrogen peroxide induced by hypoxia, which partially neutralize free radicals in the brain tissue.
Our findings also showed that ghrelin treatment increased the activity of SOD and GPx in the brain tissue of hypoxic animals and therefore antioxidant defense against hypoxia. It has been revealed that the administration of ghrelin increases the activity of antioxidant enzymes (Sibilia et al., 2003; Obay et al., 2008; Kheradmand et al., 2009). Zwirska-Korczala et al. (2007) verified that ghrelin significantly increased the activity of antioxidant enzymes such as SOD, catalase (CAT), and GPx and decreased the level of malondialdehyde (MDA) in preadipocyte cell lines (Zwirska-Korczala et al., 2007). Moreover, ghrelin pretreatment in exhaustive exercise-induced oxidative stress injury decreased MDA levels and increased SOD, CAT, and GSH activities both in the brain and liver (Samir and Mostafa, 2013). Yada et al. also showed that ghrelin increased mRNA levels of SOD in trout phagocytic leukocytes (Yada et al., 2006).
Our findings also showed that levels of TNF-α in the brain insignificantly increased in chronic hypoxia, and chronic treatment with ghrelin considerably decreased brain levels of TNF-α. TNF-α is a pleiotrophic polypeptide that plays important roles in brain immune and inflammatory activities (Feuerstein et al., 1994). TNF-α is one of the major proinflammatory cytokines (Bessis et al., 1998) that is produced by stimulation of monocytes, macrophages, T and B lymphocytes, neutrophils, and mast cells (Shohami et al., 1997b). In the CNS, it can be produced by activated astrocytes and microglia, as well as CNS-infiltrating lymphocytes and macrophages (Probert and Akassoglu, 2001). Production of TNF-α occurs in the brain in response to pathological processes such as ischemia and trauma (Feuerstein et al., 1994). TNF-α is also involved in the immune-mediated inflammation in the neurodegenerative disorders and brain edema formation following cerebral ischemia (Gregersen et al., 2000; Amantea et al., 2015). Previous study has demonstrated that TNF-α is one of the most important inflammatory cytokines related to the neuronal damage caused by the absence of oxygen (Amantea et al., 2015). It is also well known that oxidative stress and inflammatory cytokines play important roles in the development of brain injuries (Petronilho et al., 2010). Our recent study also showed that hypoxia induced brain edema and intraperitoneal injection of ghrelin reduced it in adult male rats (Hossienzadeh et al., 2013).
There are contradictory evidences showing that TNF-alpha has a dual role in brain injury in both processes of repair and damage of brain (Shohami et al., 1999; Mukandala et al., 2016). Although some research have provided a pathological role for TNF-α in traumatic and ischemic brain injury, some evidence indicates that TNF-α may play a role in the regulation of tolerance to chronic hypoxia (Rosenzweig et al., 2007). Recently, we showed that acute normobaric hypoxia increases serum TNF- α levels and ghrelin treatment ameliorates acute systemic hypoxia-induced brain edema and serum TNF-α levels (Hossienzadeh et al., 2013).
In addition, numerous studies have revealed that oxidative stress and inflammation are closely related pathophysiological processes, and one can activate another (Flohé et al., 1997; Collins, 1999; Ventura et al., 2009; Biswas, 2016). Therefore, selection of an appropriate agent that blocks some of the oxidative stress and/or inflammation pathway can provide valuable treatment for many chronic diseases. In this study, chronic administration of ghrelin effectively targeted both pathways attenuating MDA levels and brain TNF-α levels, and increasing antioxidant enzymatic defense.
Moreover, according to our previous study, ghrelin treatment increased food intake, reversed weight loss, and modulated blood glucose levels in the hypoxic condition (Babri et al., 2013). Furthermore, two recent studies from our laboratory indicated that ghrelin improved the integrity of the BBB in chronic hypoxia conditions (Mohaddes et al., 2017) and attenuated brain edema possibly through the downregulation of vascular endothelial growth factor expression (Mohaddes et al., 2015). Therefore, the administration of ghrelin may be effective for diminishing adverse responses to chronic hypoxia in the brain and serum through the reduction of oxidative stress and production of TNF-α. Further research is needed to understand the parameters that modulate brain cytokines and oxidative responses to hypoxia.
Conclusion
For the first time, our results indicated that exogenous ghrelin administration attenuated the CH-increased oxidative stress and enhanced the antioxidant defense against hypoxia-induced oxidative stress in the brain and serum. These results indicate the therapeutic benefits of ghrelin for impeding the pathogenesis of chronic hypoxia and may be effective for attenuating the adverse responses to CH.
Footnotes
Acknowledgment
This study was financially supported by the Drug Applied Research Centre of Tabriz University of Medical Sciences.
Author Disclosure Statement
No conflicting financial interests exist.