
Abstract
Abstract
Liu, Xiang-Wen, Jie Yin, Qi-Sheng Ma, Chu-Chu Qi, Ji-Ying Mu, Lang Zhang, Li-Ping Gao, and Yu-Hong Jing. Role of arcuate nucleus in the regulation of feeding behavior in the process of altitude acclimatization in rats. High Alt Med Biol. 18:234–241, 2017.—Highly efficient energy utilization and metabolic homeostasis maintenance rely on neuromodulation. Altitude exposure is known to stimulate neuroendocrine systems to respond to acute hypoxia and adaptive acclimatization. However, limited data on how the adaptive regulation of the arcuate nucleus performs in the process of altitude acclimatization are available. In the present study, male Sprague Dawley rats were transported to Huashixia, Qinghai (with an altitude of 4400 m) from Xian (with an altitude of 300 m) by air; rats were consistently raised in Xian as control. Food uptake and body weight were measured consecutively after being subjected to high-altitude condition. Contents of plasma leptin and ghrelin were analyzed by the Enzyme Linked Immunosorbent Assay (ELISA) Kits. Brain coronal sections were obtained, and neuropeptide Y (NPY), proopiomelanocotin (POMC), and c-fos immunoreactivity in arcuate nucleus were observed. Arcuate nucleus was isolated from the hypothalamus, and the mRNA of NPY and POMC were measured by quantitative real-time polymerase chain reaction. Our results showed both food consumption and body weight decreased in the high plateau compared with rats raised in the low-altitude condition. Plasma leptin increased at the early stage, and ghrelin decreased at a later stage after reaching the high plateau. The peak of c-fos immunoreactivity in the arcuate nucleus was at day 3 after reaching the high plateau. The expression level of NPY increased, and POMC decreased in the arcuate nucleus at day 7 after reaching the high plateau compared with the plain control group. These results indicate that the arcuate nucleus of hypothalamus performs an important function in regulating feeding behavior during altitude acclimatization. Our study suggested that altitude acclimation is regulated by the hypothalamus that received leptin and ghrelin signals to response by its microcircuit, including NPY- and POMC-neurons in the arcuate nucleus.
Introduction
P
In recent years, researchers have revealed that melanocortin and its analogues, that is, the derived product of hypothalamic proopiomelanocortin (POMC), perform an important regulatory function in energy metabolism and ingestion behavior (Haskell-Luevano et al., 1999; Fan et al., 2000; Butler et al., 2001; Cone, 2005). The melanocortin system (MS) in the central nervous system of mammals is mainly distributed in the arcuate nucleus of the hypothalamus (ANH), including the neuropeptide Y (NPY) neurons and their projection fibers as well as the POMC neurons and their projection fibers (Krampl et al., 2001; Kamal and Ragy, 2012; Kim and Wu, 2015). However, systemic research data on MS-ANH regulation in an actual high-altitude area are still rarely available. It is needed to explore how the MS-ANH regulates the sustainable energy supply and metabolic homeostasis in the altitude acclimatization process. Our present study aims to explore the roles and mechanisms of ANH in regulation of feeding behavior in the process of the altitude acclimatization.
Materials and Methods
Animals
Sixty male Sprague Dawley rats weighing 250 ± 10 g were purchased from the Laboratory Animal Center of the Fourth Military Medical University (Xian, China). All rats were divided into control group (altitude, 300 m) and high-altitude group. Twelve rats housed in the Animal Center of the Fourth Military Medical University were measured as control. Other rats were transported to Huashixia of Qinhai, China, (altitude, 4400 m) in Tibet Plateau by air. The animals were caged in a ventilated condition and consumed food and water ad libitum during transportation. All rats were housed under controlled temperature (23°C ± 2°C) and humidity (40% to 60%) with natural light and consumed food and water ad libitum. All experimental manipulations were undertaken in accordance with the Institutional Guidelines for the Care and Use of Laboratory Animals. Food consumption and body weight were measured consecutively, and blood was collected from the femoral artery before the rats were sacrificed. Six rats per group were perfused with 0.9% saline–4% paraformaldehyde solution through the heart under anesthetization (10% chloral hydrate, 100 mg/kg). Afterward, brains were removed and postfixed in 4% paraformaldehyde for 8 hours and transmitted to 20% sucrose–4% paraformaldehyde solution for sufficient saturation. Other rats were anesthetized and the arcuate nucleus was isolated at 4°C. The arcuate nucleus was frozen in liquid nitrogen, stored in cryogenic refrigerator at −80°C until use. Blood samples were collected through the tail vein from all animals before being sacrificed. The protocol of animal experiment was established according to the ethics approval by the Experiment Animal Ethics Committee of Lanzhou University.
Enzyme-linked immunosorbent assay
Plasma was separated by centrifuge; leptin and ghrelin were determined by using the respective Enzyme Linked Immunosorbent Assay (ELISA) Kit. Detection was carried out according to manipulation instructions. The rat leptin ELISA Kit and rat ghrelin ELISA Kit were purchased from R&D (R&D).
Immunohistochemistry
For c-fos immunostaining, five sections were selected at −2.16, −2.4, −2.64, −2.88, and −3.12 mm from the bregma of each rat. Sections were incubated in 0.3% H2O2 for 30 minutes, placed in blocking buffer containing 10% normal goat serum and 0.3% Triton X-100 in 0.01 M phosphate-buffered saline (pH 7.2) for 30 minutes at 37°C, and then incubated overnight with antibodies against mouse anti-c-fos (1:1000; Santa Cruz) at 4°C. The sections were then incubated with the corresponding biotinylated secondary antibodies (1:200) at 37°C for 1 hour and then with Strep-Avidin-HRP (1:200) at 37°C for 1 hour. Immunoreactivity was visualized with 0.05% 3,3′-diaminobenzidine as chromogen. Negative controls received the same treatment without the primary antibodies and showed no specific staining.
Immunofluorescence staining
Whole brains were cut into coronal sections at 30 μm thickness by cryotome. All sections were placed into 96-well plates with in situ hybridization buffer and stored at −20°C until use. Five sections were selected at −2.16, −2.4, −2.64, −2.88, and −3.12 mm from the bregma of each rat. Sections were respectively incubated with the antibodies of NPY (1:100; Bioworld) and POMC (1:100; Bioworld) at 4°C overnight. Then, the sections were rinsed with PBS and incubated with the corresponding secondary antibody-conjugated fluorescein isothiocyanate (FITC) at 37°C for 1 hour, respectively. Sections were mounted and observed under a microscope.
Western blotting
Total protein was extracted from the unilateral arcuate nucleus by using lysis buffer containing the protein inhibitor cocktail. Protein (50 μg) was fractionated on 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then transferred onto polyvinylidene fluoride membranes. The membranes were blotted overnight with anti-c-fos (1:400) or anti-GAPDH (1:2000) at 4°C, washed with Tris-buffered saline containing Tween 20, and then blotted with corresponding horseradish peroxidase-conjugated secondary antibodies (1:5000). Immunoblotting was visualized by enhanced chemiluminescence. All western blot experiments were conducted for a minimum of three times and with protein preparations from three independently treated animal groups.
Quantitative reverse transcription-polymerase chain reaction
Total RNA in the unilateral arcuate nucleus was isolated using an RNA Isolation Kit from TaKaRa. RNA quantity was measured by spectrophotometrical quantification (NanoDrop 2000; Peqlab, Erlangen, Germany). Total RNA (1.0 μg) was reverse transcribed to cDNA. Quantitative real-time polymerase chain reaction (PCR) was performed with Quanti-Fast SYBR Green PCR Master mix (Bio-Rad, Hercules) with a Rotorgene 3000 system (Corbett, Sydney, Australia). The primers for rat NPY were as follows: forward, 5′-GATCCAGCCCTGAGACACTGA-3′; reverse, 3′-CATGCATTGGGTGGGACA-5′. The primers for rat POMC were as follows: forward, 5′-GCCACTGAACATCTTCGTCCTC-3′; reverse, 3′-GACTGTAGCAGAATCTCGGCATC-5′. GAPDH was used as internal control. The primers of rat GAPDH were used as follows: forward, 5′-GCC ACA GTC AAG GCTGAG AAT G-3′; reverse, 3′-ATG GTG GTG AAG ACGCCA GTA-5′. The PCR conditions were as follows: 95°C for 10 s followed by 40 cycles of 95°C for 5 s and 60°C for 30 s. Gene expression was normalized to the expression of the reference gene GAPDH.
Cell counting
We performed total cell number counting in a double-blinded fashion. In these experiments, c-fos, NPY, and POMC-positive cells in arcuate nucleus from five sections (at −2.16, −2.4, −2.64, −2.88, and −3.12 mm from the bregma) of each rat were counted under 20 × objective manually. The mean number of positive cells were counted from five sections of each rat. Six rats from each group were performed in the present study.
Statistical methods
Experimental data were expressed as mean ± SD. Statistical analysis was performed using SPSS statistical program version 17. The difference between the two groups was analyzed by Student's t-test, whereas that among three or more groups was analyzed by one-way analysis of variance with the least significant difference test. The difference with p < 0.05 was considered statistically significant.
Results
Changes in food consumption and body weight during altitude acclimatization
Body weight was measured and results show that the growth rate is lower under high-altitude condition than that under low-altitude condition during 1 month (Fig. 1A). Food consumption was measured at days 3, 7, 15, and 30 after reaching the plateau, and result show that food consumption at day 3 (18 ± 3.5 g) decreased progressively compared with the control group (30 ± 2.68 g/24 hour, p < 0.01, Fig. 1B).
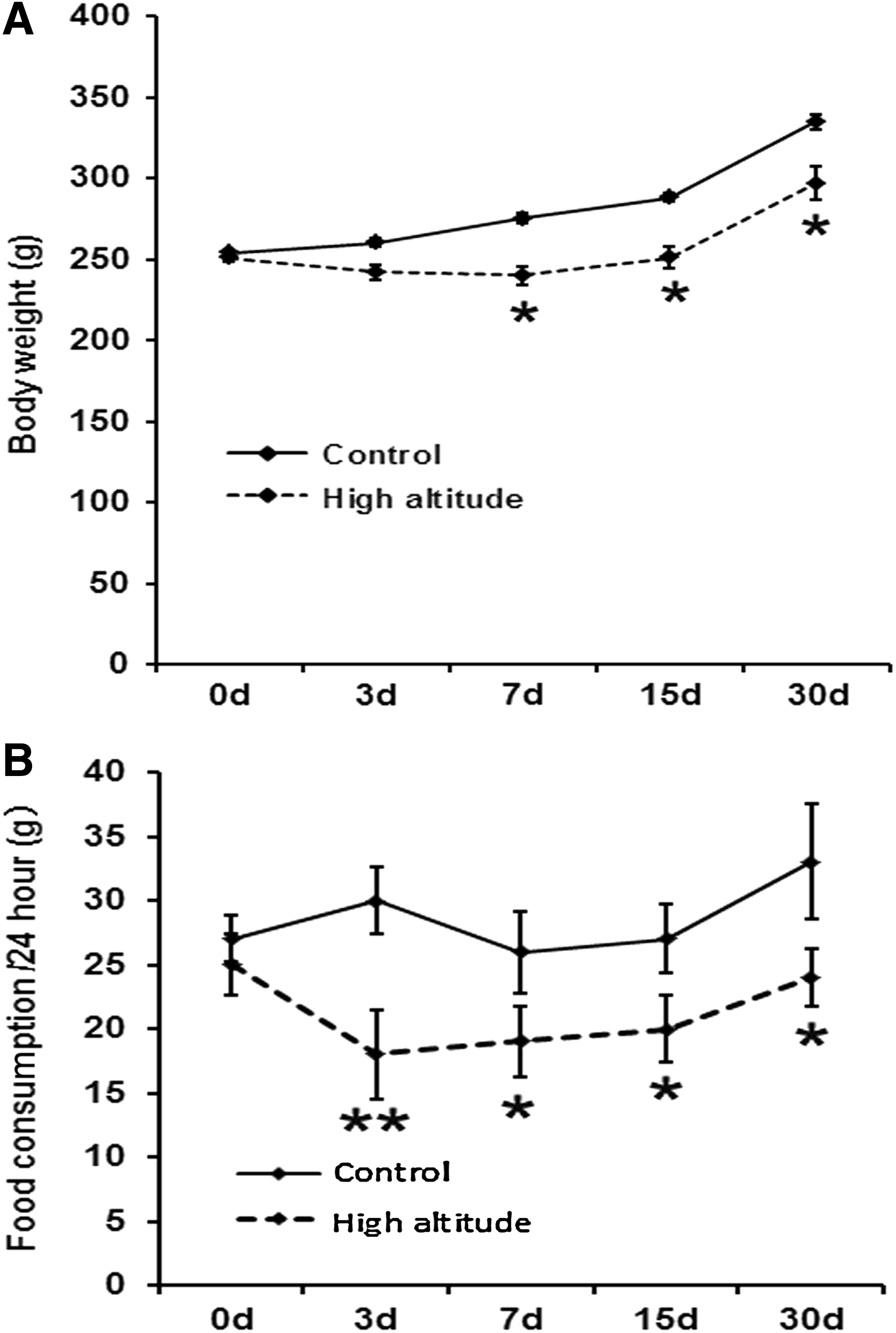
Changes in food consumption and body weight in the process of altitude acclimatization.
Changes of plasma leptin and ghrelin in the process of altitude acclimatization
Leptin, which is generated mainly from fat tissue, performs essential functions in the regulation of ingestion and metabolism. In this study, plasma leptin was measured at different time points after reaching the plateau. Results show that leptin at days 3 (3800 ± 128 pg/mL) and 7 (3817 ± 179 pg/mL) increased significantly compared with the control group (2367 ± 302 pg/mL, p < 0.05, Fig. 2A). At days 15 (2467 ± 122 pg/mL) and 30 (2387 ± 212 pg/mL), the leptin levels decreased significantly compared with day 3 (p < 0.05, Fig. 2A). Ghrelin, which is generated mainly from gut, can accelerate the ingestion, performing an opposing function to that of leptin. In this study, plasma ghrelin was measured at different time points after reaching the plateau. In the results shown in Figure 2B, ghrelin decreased significantly at day 3 (947 ± 133 pg/mL) compared with the control group (1434 ± 123 pg/mL, p < 0.05). The content of plasma ghrelin decreased significantly at day 30 (316 ± 44 pg/mL) compared with day 3 (947 ± 133 pg/mL, p < 0.01).
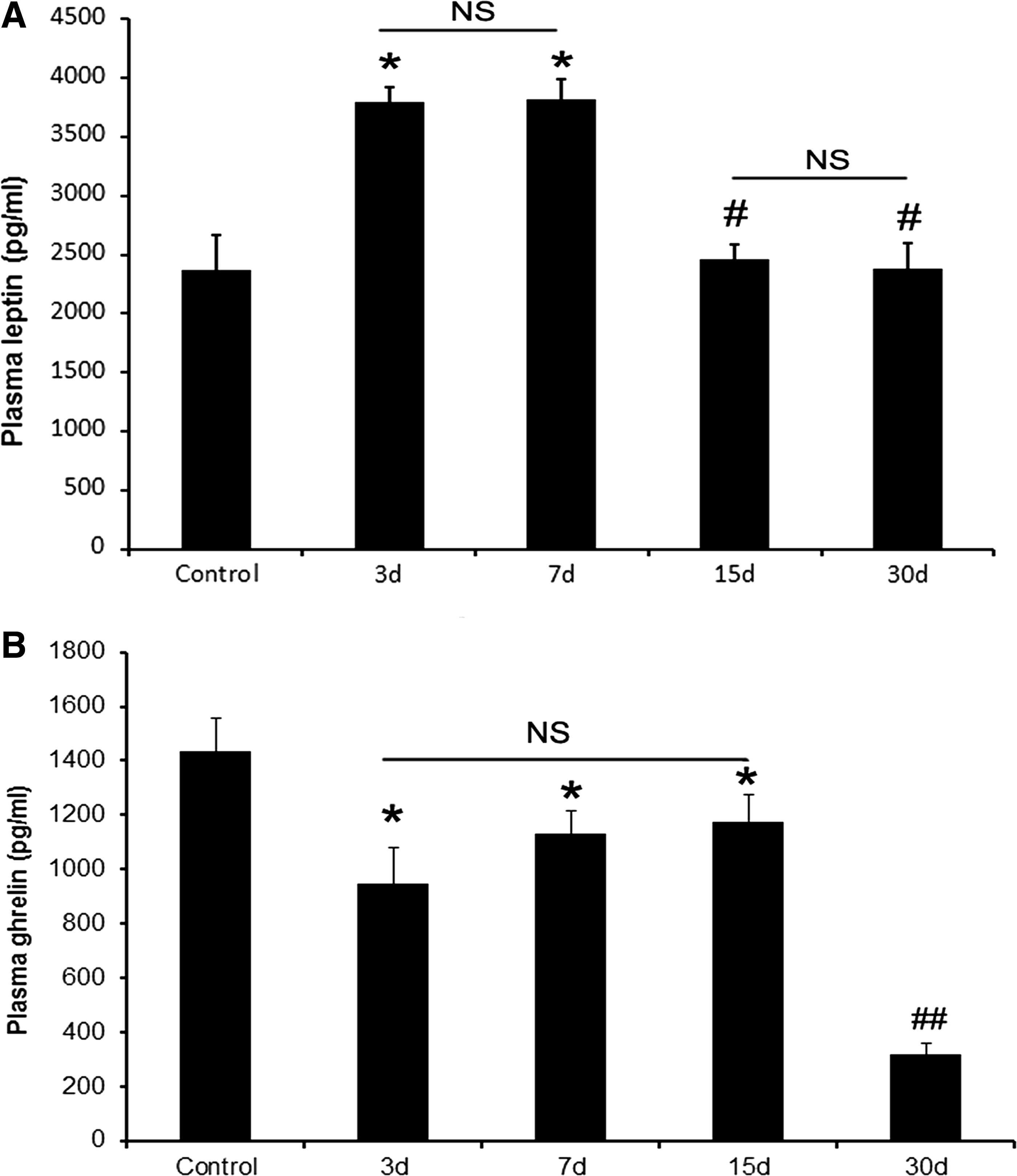
Changes in plasma leptin and ghrelin in the process of altitude acclimatization.
Time feature of c-fos-positive cell distribution in arcuate nucleus in the process of altitude acclimatization
C-fos can be considered as a molecular marker for active neurons that are fired by novel stimulation. Based on this method, immunohistochemistry and cell counting were used to evaluate the expression of c-fos in arcuate nucleus in the process of altitude acclimatization. The results show more c-fos-positive cells in the arcuate nucleus at day 3 compared with the control group, decreasing at days 7 and 15 relative to day 3. At day 30, c-fos-positive cells are few (Fig. 3A, B). In addition, c-fos level was quantified using western blotting in the isolated arcuate nucleus tissue. Similarly, c-fos expression in arcuate nucleus increased at day 3 compared with the control group and decreased at days 7, 15, and 30 compared with day 3 (Fig. 3C, D).
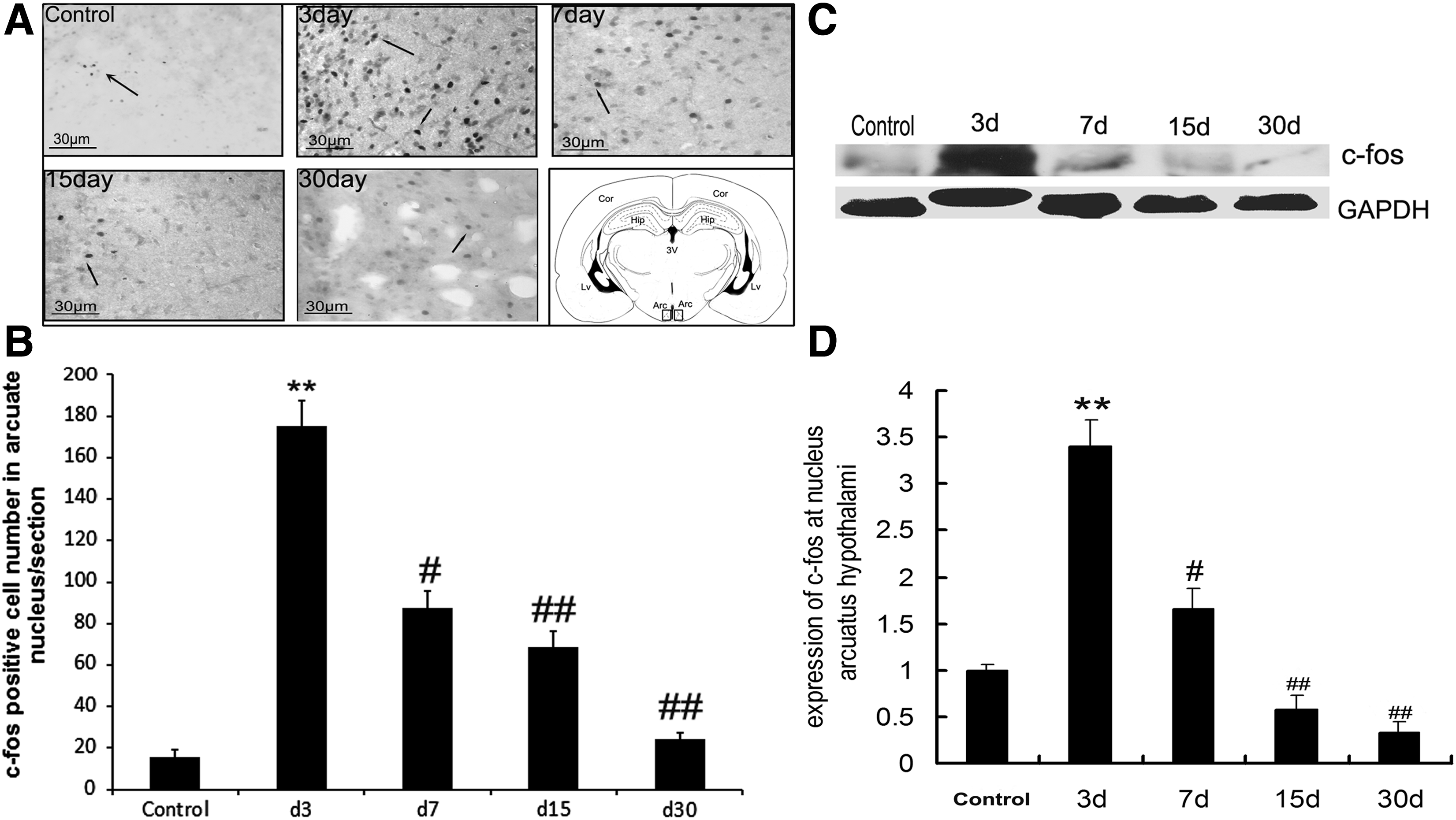
Activity of c-fos in the arcuate nucleus of rats at different time points after reaching the plateau.
Changes in NPY expression in arcuate nucleus during altitude acclimatization
Immunofluorescence and cell counting were used to evaluate the immunoreactivity of NPY in arcuate nucleus. The results indicate that the number of NPY-positive cells in arcuate nucleus increased significantly at day 7 compared with the control group and decreased at day 30 compared with day 3 (Fig. 4A, B). Similarly, the NPY mRNA in arcuate nucleus at day 7 was 10-fold higher than that of control and decreased at day 30 (Fig. 4C).
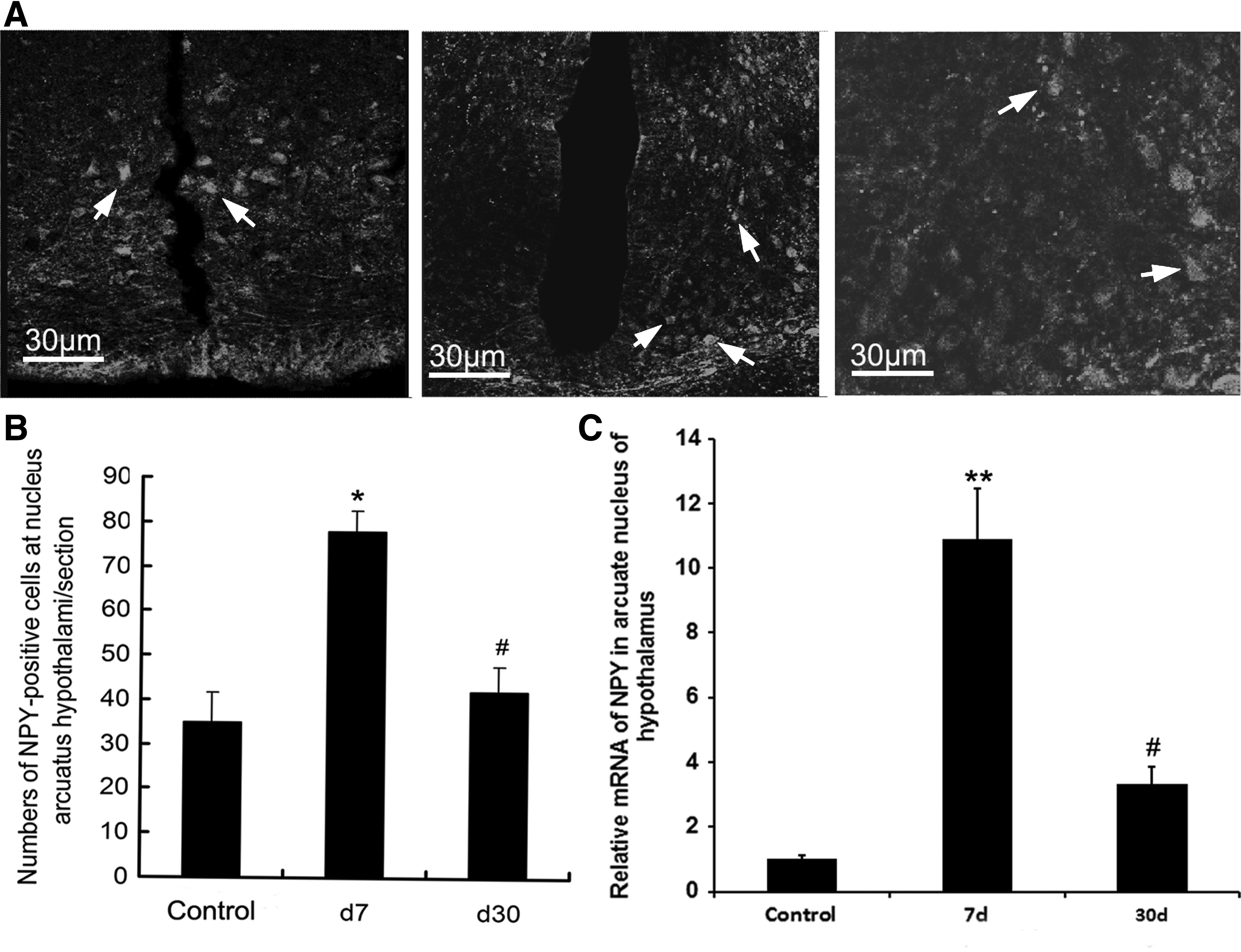
Changes in NPY in the arcuate nucleus of rats at days 7 and 30 after reaching the plateau.
Changes in POMC expression in arcuate nucleus during altitude acclimatization
POMC neuron is an inverse effect compared with NPY neuron in ingestion regulation. In this study, immunofluorescence and cell counting were used to evaluate the immunoreactivity of POMC in arcuate nucleus. Results show that the number of POMC-positive cells decreased at day 7 compared with the control group and further decreased at day 30 (Fig. 5A, B). The POMC transcript level in arcuate nucleus was determined to further evaluate the changes during altitude acclimatization. The POMC transcript level in arcuate nucleus at days 7 and 30 was ∼0.2-fold and 0.7-fold lower than that of control (Fig. 5C).
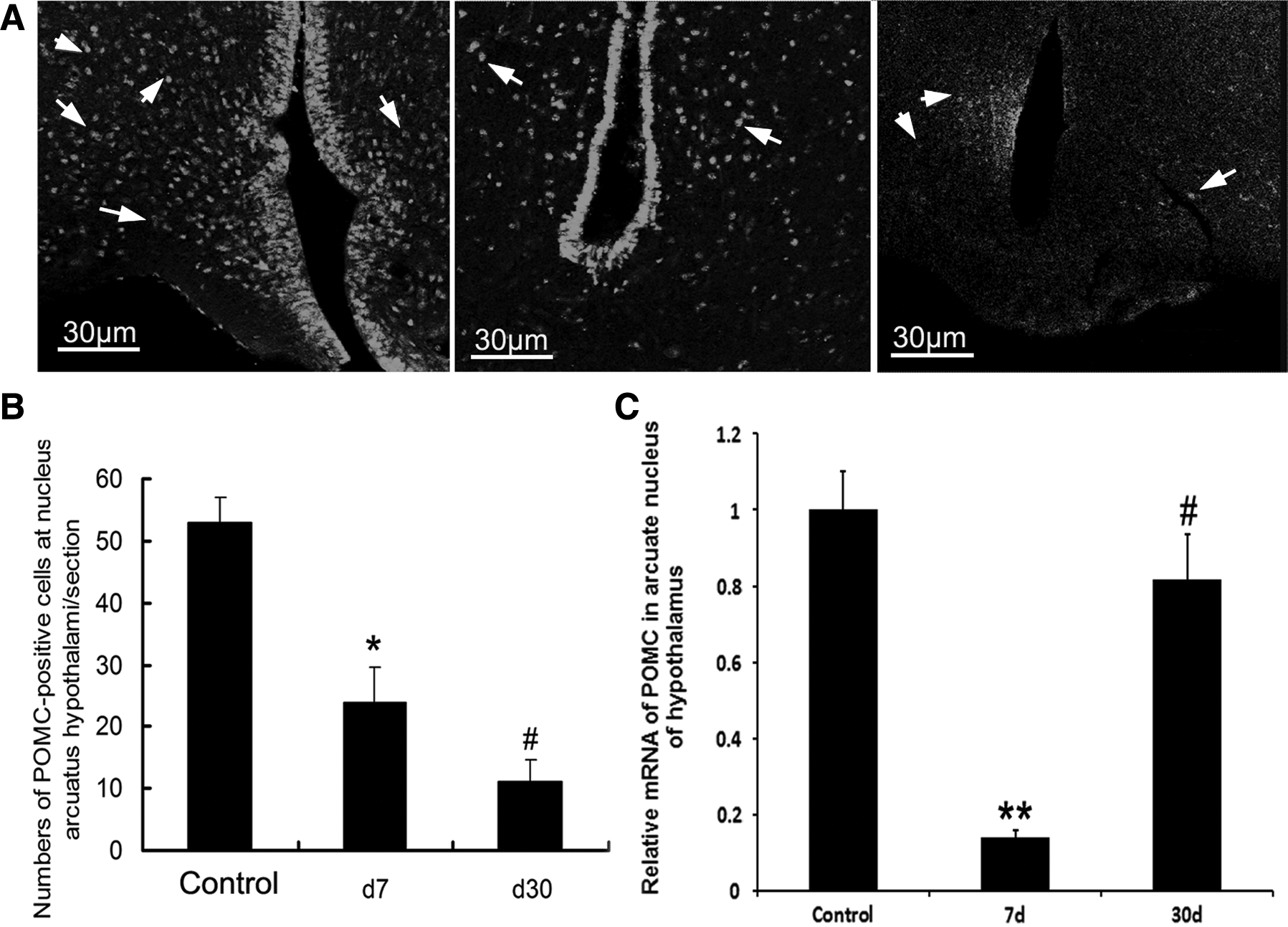
Changes in POMC in the arcuate nucleus of rats at days 7 and 30 after reaching the plateau.
Discussion
In the present study, we found that the plasma leptin and ghrelin fluctuated when the rats lived in a high-altitude place from low-altitude condition during 1 month. Likewise, the feeding behavior and neuron activity in ANH have been changed in this process. The adaptation of the organism to a critical environment can be accomplished by regulating the metabolic pattern and response (Larsen et al., 1997; Simonson, 2015). As a major component in the energy utilization, oxygen can be utilized by intracellular mitochondria through oxidative phosphorylation during the production of ATPs, which may provide energy for the body. During hypoxia, especially for the people who initially access a highland, the utilization of nutrients may be changed accordingly (McClelland et al., 1998; Sayers et al., 2010; Moza and Gujral, 2016). In addition, several signaling molecules may affect the requirement status of the body for food and subsequently lead to the change in the ingestion behavior in response to the alteration of the utilizing pattern of glucose, lipids, and proteins (Muller and Geisen, 1996; Bergen and Mersmann, 2005). Previous research has shown that decreased food consumption may be present when a man stays in a high-altitude region for ∼7 days, and these changes are associated with insulin and leptin (Wei et al., 2015). As shown in the present study, reduced food consumption may be initially observed at day 3 after entering a high-altitude region, and food consumption is associated with changes in body weight. We found that the body weight of animals may initially decrease at an early stage and gradually return to normal status.
Leptin, which is mainly secreted from the adipose tissue, is able to deliver the signal from the adipose tissue to the brain, particularly to the hypothalamus (Denroche et al., 2016). The hypothalamus functions in a modulating behavior and metabolic response by integrating the signals from leptin and other molecules, consequently modifying energy balance and weight (Luo et al., 2011; Kim et al., 2014). Leptin is a molecule that can reduce body weight by inhibiting the appetite and increasing energy consumption (Pelleymounter et al., 1995; Dalamaga et al., 2013; Moon et al., 2013; Park and Ahima, 2015). Meanwhile, the NPY and POMC-associated neurons are targets for leptin-based regulation. For appetite inhibition, the POMC-positive neuron is coexpressed with cocaine- and amphetamine-regulated transcript (CART); this neuron and the coexpression of POMC/CART may be activated by leptin (Ahima et al., 1999; Cowley et al., 2001; Abizaid et al., 2006; Robertson et al., 2008). Ghrelin, a hormone-like peptide that was first discovered in 1999, is produced by endocrinal cells in the stomach and the proximal intestine as a ligand of growth hormone secretagogue receptor. Ghrelin is further secreted in the circulating blood (Kojima et al., 1999). A recent study showed that intracerebroventricular administration of ghrelin to rats is associated with increased food intake and decreased oxygen consumption, indicating that this substance can improve appetite and reduce energy consumption (Nakazato et al., 2001). Two independent ghrelin-expressed cell populations were found in the arcuate nucleus: one, which is coexpressed with NPY, serves to improve appetite; and the other, which is coexpressed with POMC and CART, functions in appetite suppression (Morton et al., 2006). In the present study, plasma leptin of rats increased at an early stage after reaching a high plateau and decreased at a later stage. In addition, plasma ghrelin remained at a relatively high level and decreased at a later stage. Consistent with this fluctuation of plasma hormone, food consumption decreased at an early stage and gradually increased at a later stage after reaching a high plateau.
The arcuate nucleus is located in the periventricular zone of the hypothalamus, encircling the ventral aspect of the third cerebral ventricle, anteriorly in front of the infundibular recess and posteriorly to the ultimate end of the corpus albicans recess (Cone et al., 2001). Several neurons with different functions are present in the arcuate nucleus, including growth hormone-releasing factor neurons, endorphin neurons, P-substance neurons, NYP neurons, POMC neurons, melanocortin neurons, dopamine neurons, and GABA neurons (Coppari et al., 2005; Huo et al., 2009; German et al., 2011; Berglund et al., 2012). The functions of the arcuate nucleus are multiple and intricate in behavior modulation, including sleep regulation, reproductive behavior regulation, energy expenditure regulation, and feeding behavior regulation (Hunt et al., 2013; Teubner et al., 2015). However, few data are available on the involvement of arcuate nucleus in the process of altitude acclimatization. Our study revealed that adaptive regulation involves arcuate nucleus during altitude acclimatization. For instance, several neurons were fired and labeled by c-fos immunoreactivity in the arcuate nucleus under high-altitude condition, indicating that certain neurons, such as NPY-neuron and POMC-neuron, are implicated in the process of altitude acclimatization when animals access a high-altitude region. However, the effect of arcuate nucleus is marked in this process, and numerous detailed signal pathways need to be clarified in future studies. This study was conducted in a highland environment and may be more realistic than the simulated hypobaric oxygen chamber. The obtained data can favorably reflect the true response of animals in the process of altitude acclimation. However, some issues need to be clarified, for example, what is the role of hyperglycemia regulation on feed behavior changes, the metabolic rate in high-altitude condition compared with the low-altitude environment after acclimatization for a long time.
In conclusion, our study found that the fluctuation of plasma leptin and ghrelin is consistent with the changes of feeding behavior and body weight when rats were reared in high-altitude conditions. Combining the response of neurons in arcuate nucleus during this process, we infer that plasma leptin or ghrelin acts on its receptors located in the arcuate nucleus and regulates NPY- or POMC-neuron to modify feeding behavior, which facilitate energy expenditure to adapt to the hypoxia condition in the process of altitude acclimatization. In addition, future studies should attempt to explore the interaction of leptin and ghrelin in the hypothalamus during altitude acclimatization.
Footnotes
Acknowledgment
This work is partly supported by the National Natural Science Foundation of China (81370448 and 81570725) to Y.H.J.
Author Disclosure Statement
No conflicting financial interests exist.