
Abstract
Abstract
Cao, Xue-Feng, Zhen-Zhong Bai, Lan Ma, Shuang Ma, and Ri-Li Ge. Metabolic alterations of Qinghai–Tibet plateau pikas in adaptation to high altitude. High Alt Med Biol. 18:219–225, 2017.—To determine specific metabolic alterations in the myocardium of plateau pikas (Ochotona curzoniae) and potential metabolic biomarkers involved in their adaptation to the high-altitude environment of the Qinghai–Tibet Plateau. Ten pikas were captured by traps in the Kekexili Reserve (4630 m a.s.l; n = 5) and at the foot of the Laji Mountain (2600 m a.s.l; n = 5) on the Qinghai–Tibet Plateau, Qinghai Province, China. Metabolite levels were determined by gas chromatography time-of-flight mass spectrometry (GC-TOF-MS) metabolomics, and multivariate statistical analysis was performed. Several metabolites involved in carbohydrate, fat, energy, and redox homeostasis pathways were significantly altered in pikas living at 4630 m. In addition, those pikas showed increased levels of lactic acid, sarcosine, 4-hydroxybutyrate, methionine, tartaric acid, ribose, tyrosine, pentadecanoic acid, 2-monoolein, 3,5-dihydroxyphenylglycine, trehalose-6-phosphate, succinic acid, myoinositol, fumaric acid, taurine, 2-hydroxybutanoic acid, gluconic acid, citrulline, and glutathione, but decreased levels of oleic acid and 2′-deoxyadenosine 5′-monophosphate. Metabolic activity is significantly altered in the myocardium of pikas in the high-altitude areas of the Qinghai–Tibet Plateau. This study provides important insights into metabolic biomarkers related to the adaptation of pikas to high-altitude hypoxia.
Introduction
H
The plateau pika (Ochotona curzoniae), a member of the Ochotonidae family, is a small rabbit-like mammal native to the Qinghai–Tibet Plateau (Yu et al., 2012). Some pika fossil samples found on the Qinghai–Tibet Plateau are about 20–30 million years old (Wang et al., 2008). Pikas prefer to live in the mountain areas in an altitude of 2500–5300 m a.s.l., and thus they are highly adapted to the extreme hypoxia, cold, and intense ultraviolet radiation. Previous studies have investigated the effects of hypoxia on the pulmonary circulation, liver, and neuroendocrine system (Ge et al., 1998), whereas the cardiovascular adaptations of pikas to hypoxia remain poorly understood. Sakai et al. (2003) showed a lower right ventricular weight in pikas than in rats living at sea level, which was related to the lower pulmonary arterial pressures and minimal right ventricular hypertrophy (Sakai et al.,2003). Qi et al. (2008) observed that pikas could improve their adaptation to hypoxia by increasing mitochondrial surface density, microvessel density, and myoglobin content in heart (Qi et al., 2008). Interestingly, pikas also showed left ventricular hypertrophy and higher vascular endothelial growth factor mRNA expression than rats, indicating that pikas had better cardiac efficiency in a severe hypoxic environment because of the long-term adaptation (Pichon et al., 2013).
Metabolites are the small molecule intermediates and products of metabolism that together compose the complete metabolome of a biological sample, providing insight into the metabolic processes in the body (Fuss and Cheng, 2016). Metabolites are the end products of cellular regulatory processes and their levels can be regarded as the ultimate response of biological systems to genetic or environmental changes (Fiehn, 2002). It is likely that functional cardiovascular changes in pikas living on the hypoxic Qinghai–Tibet Plateau are accompanied by changes in metabolite levels associated with fuel and energy balance, particularly as the purpose of oxygen delivery and utilization in tissues is to support oxidative metabolism.
However, little is known about the mechanisms of hypoxia adaptation in plateau pikas from the perspective of myocardial metabolism. To address this problem, we investigated the changes of cardiac metabolites in pikas in Qinghai, China, by gas chromatography time-of-light mass spectrometry (GC-TOF-MS) metabolomics.
Materials and Methods
Animals
Ten pikas were captured by traps in the Kekexili Reserve (4630 m a.s.l.; n = 5) and at the foot of the Laji Mountain (2600 m a.s.l.; n = 5) on the Qinghai–Tibet Plateau, Qinghai Province, China. The two groups were matched in terms of gender, weight, and living environment (e.g., vegetation, capture time, and site of sampling) (Table 1). Myocardial tissues were harvested and stored in liquid nitrogen until analysis. This study was approved by the Ethics Committee of the Medicine College of Qinghai University.
Main reagents and instruments
Bis-(trimethylsilyl)-trifluoroacetamide (BSTFA) with 1% trimethylchlorosilane (TMCS) solution was purchased from Regis Technologies Inc. (Morton Grove, IL). L-2-chlorophenylalanine (internal standard, CAS#:103616-89-3) was obtained from Shanghai Hengbai Biotech. Co., Ltd., (Shanghai, China). The instruments used in this study included an Agilent 7890A gas chromatography system (Agilent), a Leco-Chroma Pegasus HT TOF mass spectrometer (Leco), and a grinding mill (Shanghai Jingxin Technology Co., Ltd., Shanghai, China).
Myocardial samples and GC-TOF-MS analysis
About 50 mg of frozen myocardial tissues collected from each pika was added into a 2-mL eppendorf tube, and then extracted with 0.4 mL of extraction liquid (methanol–chloroform, 3:1) and 20 μL of L-2-chlorophenylalanine (1 mg/mL stock in double distilled H2O [ddH2O]) as an internal standard in a 2 mL microcentrifuge tube. Samples were mixed by vortex for 30 seconds, homogenized in a ball mill for 6 minutes at 45 Hz, and then centrifuged at 12000 rpm for 15 minutes at 4°C. The resulting supernatant (0.35 mL) was transferred into a 2 mL GC/MS glass vial. The extracts were dried in a vacuum concentrator without heating, and 80 μL of methoxyamination hydrochloride (20 mg/mL in pyridine) was added and incubated at 80°C for 30 minutes. Then, 100 μL of BSTFA reagent (1% TMCS, v/v) was added and incubated at 70°C for 2 hours. GC-TOF-MS was performed on an Agilent 7890 gas chromatography system coupled to a Pegasus HT TOF mass spectrometer. A DB-5MS capillary column coated with 5% diphenyl cross-linked with 95% dimethylpolysiloxane (inner diameter: 30 m × 250 μm, film thickness: 0.25 μm; J&W Scientific, Folsom, CA, USA) was used. A 1-μL aliquot of the analyte was injected in splitless mode. Helium was used as the carrier gas, wherein the front inlet purge flow was 3 mL/min and the gas flow rate through the column was 1 mL/min. The initial temperature was kept at 50°C for 1 minute, and raised to 290°C at a heating rate of 10°C/min, and then kept at that temperature for 12 minutes. The temperatures of the injection, transfer line, and ion source were 280, 270, and 250°C, respectively. The energy was −70 eV in electron impact mode. The MS data were acquired in full-scan mode with the m/z range of 85–600 at a rate of 20 spectra per second after a solvent delay of 366 seconds.
Multivariate statistical analysis
The raw GC-TOF-MS data were analyzed as described previously (Lisec et al., 2006). Chroma TOF 4.3X software (LECO) and LECO-Fiehn Rtx5 database were used for raw peak exaction, data baseline filtering and calibration, peak alignment, deconvolution analysis, peak identification, and integration of the peak area. The missing values were replaced with half of the minimum value. A total of 405 peaks that represented 272 metabolites were detected by the interquartile range denoising method. The retention time index (RI) method was used for peak identification, and the RI tolerance was set to 5000. In addition, the internal standard normalization method was used in the data analysis, and the normalized data about the peak number, sample name, and normalized peak area were fed into the SIMCA 14.0 software package (Umetrics, Umea, Sweden) for a series of multivariate statistical analyses. Principal component analysis (PCA) was performed to observe the data set and display the similarities and differences. The partial least squares-discriminant analysis (PLS-DA) was used for cluster analysis, and linear regression was used for categorical variables in supervised learning. The R2 values were used to describe how well the data were mathematically reproduced. They ranged from 0 to 1, with 1 indicating perfect fitness. The Q2 values were recorded as the percentage variation of the response predicted by the model, or how accurately the model could predict new data. The PLS-DA model was validated by permutation tests (n = 200) and then converted into corresponding orthogonal projection to latent structures-discriminant analysis (OPLS-DA) models.
Metabolite identification
The discriminating compounds were identified by comparing RI with data from the LECO/Fiehn Metabolomics Library, which gave a similarity value to indicate the accuracy of compound identification (Kind et al., 2009). Peaks with a similarity index higher than 700 were assigned compound names, those with a similarity value between 200 and 700 were processed as putative annotation, whereas those with a similarity value lower than 200 were processed as analytes. After the OPLS-DA analysis, differentially expressed metabolites between the two groups could be identified. Potential candidates were chosen based on the contribution of the variable importance for the projection (VIP) extracted from the first principal component of the OPLS-DA analysis. The greater the consistent difference in metabolite levels between groups, the more important a metabolite would become in creating the final model, as indicated by the VIP value. VIP values higher than 1.0 were first selected as significantly changed metabolites, and the rest variables were assessed by Student's t-test. p values <0.05 were considered to be statistically significant. Variables not significantly changed were discarded. Significantly different metabolites were cross-referenced to the pathways by searching KEGG databases (www.genome.jp/kegg/).
Results
Characterization of GC-TOF-MS data
The GC-TOF-MS data were normalized and exported into Chroma TOF 4.3X software and LECO-Fiehn Rtx5 database for analysis of metabolic variability in the myocardial samples. Thirty-eight of a total of 271 peaks were identified as metabolites by the interquartile range denoising method. The score plots of the PCA model and the OPLS-DA model obtained from the 4630-m group and the 2600-m group are shown in Figure 1. It showed that R2Y = 0.354 and Q2Y = 0.908, indicating that the models were stable and had good fitness and predictive ability. We used sevenfold cross-validation to estimate the robustness and the predictive ability of our models. The R2 and Q2 intercepts were 0.85 and −0.0967 after 200 permutations, respectively. The low Q2 intercept indicated high robustness and reliability of the model and a low risk of over fitting. A loading plot was constructed to indicate the contribution of variables to the difference between the two groups, and the important variables situated far from the origin. VIP values exceeding 1.0 were first selected as changed metabolites, and the remaining variables were assessed by Student's t test (p > 0.05). Variables that were not significantly changed were discarded (Table 2).

PCA and OPLS-DA analysis of GC-TOF-MS metabolite profiles. GC-TOF-MS, gas chromatography time-of-flight mass spectrometry; OPLS-DA, orthogonal projection to latent structures-discriminant analysis; PCA, principal component analysis.
Fold-change: the 4630-m group versus the 2600-m group.
GSH, glutathione; RT, retention time; VIP, variable importance for the projection.
Differentially expressed metabolites between the 4630-m and the 2600-m groups
Table 2 gives several differentially expressed metabolites between the 4630-m and the 2600-m groups (VIP >1 and fold change >1.5 or fold change <0.5). Pikas living at 4630 m showed significantly higher levels of lactic acid, sarcosine, 4-hydroxybutyrate, methionine, tartaric acid, ribose, tyrosine, pentadecanoic acid, 3,5-dihydroxyphenylglycine, 2-monoolein, trehalose-6-phosphate, succinic acid, myoinositol, fumaric acid, taurine, 2-hydroxybutanoic acid, gluconic acid, citrulline, and glutathione (GSH), but significantly lower levels of oleic acid and 2′-deoxyadenosine 5′-monophosphate in the myocardial tissues (p < 0.05).
Receiver operating characteristic (ROC) curves have been used in evaluating the diagnostic accuracy of biomarkers in previous studies. In this study, ROC curves were plotted to evaluate the prognostic performance of differentially expressed metabolites. The closer the apex of the ROC curve toward the upper left corner, the higher the area under the ROC curve (AUC), and the greater the discriminatory ability of the test. The AUC value was used to measure the discrimination between experimental and control subjects. An AUC value of 1 indicated perfect accuracy, an AUC value >0.9 indicated extremely high accuracy, whereas an AUC value <0.7 indicated an inaccurate test. Interestingly, we found a large number of differential metabolites with very high AUC values (Fig. 2). The highest AUC values (1 or 0.92) were observed in sarcosine, tartaric acid, tyrosine, 2-hydroxybutanoic acid, 3,5-dihydroxyphenyl glycine, pentadecanoic acid, and 4-hydroxybutyrate (Fig. 2). Thirteen metabolites (methionine, myoinositol, ribose, GSH, taurine, trehalose-6-phosphate, citrulline, lactic acid, fumaric acid, succinic acid, methyl palmitoleate, and sitosterol) had an AUC value >0.76, indicating an excellent discriminatory ability. Therefore, these differential metabolites shown in Figure 2, such as sarcosine, tartaric acid, tyrosine, 2-hydroxybutanoic acid, and 3,5-dihydroxyphenyl glycine, could serve as potential biomarkers of the auxiliary diagnosis and hypoxia adaptation of humans and animals living on the Qinghai–Tibet Plateau.
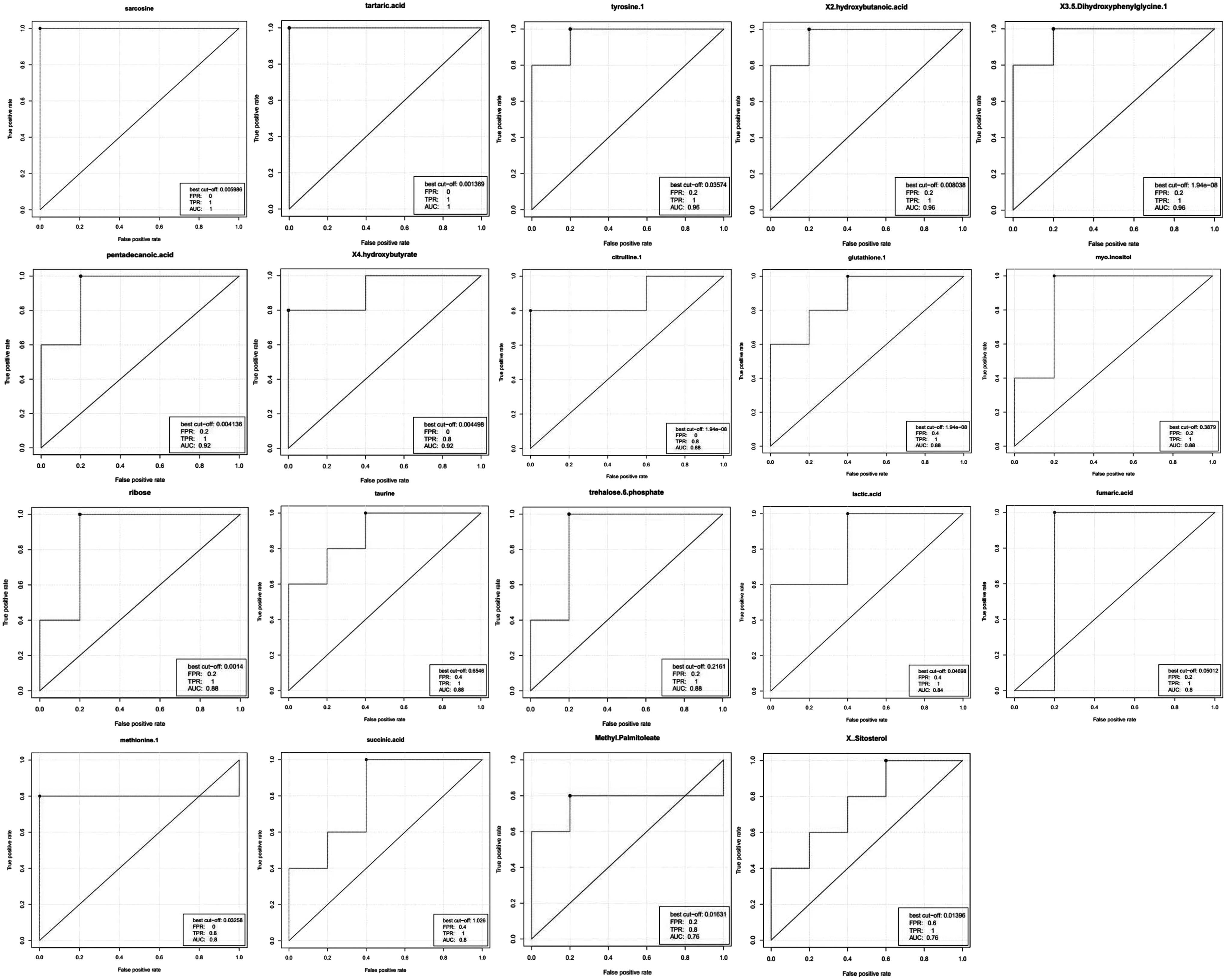
ROC plots of metabolites with the highest AUC values. ROC, receiver operating characteristic; AUC, area under the curve.
Discussion
In this study, alterations of several metabolites were observed in the myocardium of pikas at a high altitude of 4630 m on the Qinghai–Tibet Plateau. During hypoxia, the lack of oxygen delivery results in the decrease of adenosine triphosphate (ATP), the build-up of lactate, and alterations of many other metabolites. The tricarboxylic acid cycle (TCA) comprises a series of enzymatic reactions to generate energy from intermediates derived from carbohydrates, fats, and proteins. The levels of succinic acid, fumaric acid, and TCA cycle intermediates were elevated in the higher altitude (4630 m) pikas, and their high VIP levels suggested the importance of the TCA cycle in hypoxic adaptation. Hypoxia may cause a blockage in the pathway, with intermediates piling up behind the block. Succinate accumulation could be considered as a metabolic signature of ischemia/hypoxia. The production of succinate improved the redox state and thus helped to adapt to hypoxia (Wiesner et al., 1988). Fumarate has been suggested to be cardioprotective through activation of the Nrf2 antioxidant pathway (Ashrafian et al., 2012). However, further studies are needed to elucidate their potential mechanisms in pikas. Notably, no changes were observed in other intermediates involved in the TCA cycle. However, it remains unclear whether the increased levels of metabolites result from the increased flux rates or blocks in the TCA cycle with accumulation of upstream intermediates. The increase in metabolism in the TCA cycle results in an increase in energy supplies in situations of low energy production such as hypoxia, which is observed during the bouts of exercise. This may occur, however, at the expense of increased oxidative stress if the oxygen supply was unable to meet the needs of the electron transport chain. We indeed observed dramatic changes in pathways involved in the GSH synthesis. However, increased lactate production implied that glucose oxidation may not be increased in high altitude pikas.
The accumulation of some fatty acids, such as 4-hydroxybutyrate, pentadecanoic acid, 2-monoolein, 2-hydroxyvaleric acid, methyl palmitoleate, and 2-hydroxybutanoic acid in 4630-m pikas have fold changes higher than 1.4, indicating an increase in lipid metabolism in those pikas. Cardiomyocytes are dependent heavily on fatty acid oxidation for energy production, which consumes more oxygen per carbon oxidized. However, numerous studies have shown that pikas have evolved adaptive mechanisms to deal with challenges such as high resting metabolic rate and have lower rates of oxygen consumption (Wang et al., 2006). The AUC value of 2-hydroxybutanoic acid, pentadecanoic acid, and 4-hydroxybutyrate was 0.96, 0.92, and 0.92, respectively. Therefore, the increase in those fatty acids could help to alleviate the shortage of energy under chronic hypoxic conditions.
Oxidant stress may result from the failure to capture electrons generated. Thus, it is not surprising to observe changes in pathways involved in regulating the redox state of cells in high-altitude pikas. GSH, which played a critical role in the redox regulation and antioxidant protection in all cells (Emiliani et al., 2014), was increased in higher altitude pikas. The observed increase in 2-hydroxybutanoic acid may be associated with the changes in GSH, as it is a byproduct of the conversion of cystathionine to cysteine and GSH, and may reflect the increased flux of methionine into this pathway. Methionine and taurine levels were also increased in high-altitude pikas. Taurine, a metabolite of cysteine, is a β-amino acid with a high concentration in the excitable tissues. It has an antioxidant action, and thus taurine supplementation can reduce the increased oxidative stress in association with decreased superoxide radical production (Silva et al., 2011). Taurine has been demonstrated to be essential for cardiac function, and thus it could be used to increase cardiac contractility in heart failure (Schaffer et al., 2016). Several metabolites were significantly increased in the myocardium of 4630-m pikas with direct implications for cardiac function, including sarcosine, trehalose-6-phosphate, tyrosine, and 3,5-dihydroxyphenylglycine. The fold changes of these metabolites were consistent with that of the AUC values (Table 2, Fig. 2). Sarcosine is an N-methyl derivative of the amino acid glycine that can increase myocardial contractile force and cardiac output. Trehalose is a disaccharide used as an energy source in lower organisms, but it may also protect mammals from antioxidant damage. Trehalose reduced ultraviolet radiation-induced damage caused by reactive oxygen and nitrogen species and decreased changes in the corneal optics (Cejkova et al., 2010). Tyrosine supplementation may enhance individuals' cognitive function in a hypoxic and/or cold environment (Taylor et al., 2015). Exposure to a cold environment results in changes in the concentration of dopamine, epinephrine, and norepinephrin (Avakian et al., 1984). As a consequence, plateau pika has a relatively high response sensitivity in a low oxygen environment. However, further research is needed to determine whether this is the case in plateau native animals. Elevated 3,5-dihydroxyphenylglycine in heart is a known response to hypoxia, which may modulate blood flow at high altitudes (Liu et al., 2011). Myoinositol is a ubiquitous carbohydrate that participates in nucleic acid metabolism, cellular energy metabolism, protein synthesis, and nourishing brain cells. Inosine, a breakdown product of adenosine associated with inflammation, hypoxia, and tissue injury, was also increased in high-altitude pikas. However, the levels of 3′-deoxyadenosine 5′-monophosphate were decreased in the 4430-m group. However, it remains unknown whether this is associated with a global increase in nucleotide breakdown as indicated by the increase in Myoinositol, a decrease in nucleotide synthesis as indicated by the increase in ribose, or changes in overall energy balance. It can enhance the activity of many enzymes, particularly coenzyme A and pyruvate oxidase, thus facilitating the metabolism of myocardial cells under hypoxic conditions.
The potential roles of other differentially expressed metabolites (i.e., sitosterol, methyl palmitoleate, and oleic acid) in hypoxic adaptation and cardiac function are not well understood. However, it is important to note that all of these metabolites have high VIP values and may be clinically relevant for the hypoxia adaptation of pikas. Further studies are warranted to better understand the functions of those metabolites.
In summary, we performed metabolomics analysis in pikas living at middle- and high-altitude areas of the Qinghai–Tibet Plateau. The results clearly showed increased glycolysis and decreased fatty acid oxidation, which were consistent with the changes observed in some other species and organs exposed to hypoxia. These changes may reflect possible adaptation to oxidative stress initiated by sustained oxidative reactions proceeding in the presence of inadequate oxygen. However, some other metabolites with unknown functions to heart were also noted.
One limitation of this study is that the metabolite levels were determined at a steady state, whereas a metabolite may be elevated through either increased production, decreased utilization, or both. Thus, flux analysis, for example, of glucose metabolism or GSH synthesis needs to be performed. In addition, we did not determine the levels of key enzymes that regulated the metabolite levels, which could be the focus of our next research. Pikas were captured at middle (2600 m a.s.l.) and high (4630 m a.s.l.) altitudes, so the relevance of the results to organisms at lower altitudes needs to be further validated. Finally, a wide variety of factors are implicated in the adaptation to altitude, such as cold and ultraviolet irradiation, which need to be considered in future controlled studies.
Conclusions
This is the first report of the identification of metabolic pathways in myocardial tissues of pikas associated with hypoxia adaptation at high altitudes by GC-TOF-MS-based metabolomics. The GC-TOF-MS analysis demonstrated metabolic alterations in pikas of the 4630-m group. Differential metabolites in myocardial tissues with high VIP values, such as sarcosine, tartaric acid, tyrosine, 3-hydroxypropionic acid, 3,5-dihydroxyphenylglycine, pentadecanoic acid, 4-hydroxybutyrate, citrulline, GSH, myoinositol, lactic acid, ribose, succinic acid, methionine, taurine, and trehalose-6-phosphate, may be of great importance for hypoxia adaptation at high altitudes, in particular for protection from oxidative stress and maintenance of energy production. Metabolomics can provide more comprehensive and reliable information to better understand metabolic response under the hypoxic condition. The pathway and function analysis may provide new insights into hypoxia adaptation mechanisms and novel biomarkers for chronic exposure to high altitude.
Footnotes
Acknowledgments
This study was supported by the National Program on Key Basic Research Project of China (No. 2012CB518200), National Natural Science Foundation of China (No. 31571231), and Qinghai-Utah Joint Research Key Lab for High Altitude Medicine (No. 2014-ZJ-Y39).
Author Disclosure Statement
No competing financial interests exist.