
Abstract
Abstract
Moraga, Fernando A., Jorge Osorio, Rodrigo Calderón-Jofré, and Andrés Pedreros. Hemoconcentration during maximum exercise in miners with chronic intermittent exposure to hypobaric hypoxia (3800 m). High Alt Med Biol. 19:15–20, 2018.
Objective:
To evaluate the effect of maximum exercise on hemoconcentration in miners with chronic intermittent hypobaric hypoxia (CIHH) at 3800 m.
Materials and Methods:
Sixteen miners with CIHH at high altitude (3800 m) were subjected to maximum exercise levels on a cycle ergometer, increasing exercise load by 50 W every 3 minutes at sea level and high altitude (3800 m). During exercise, arterial oxygen saturation and heart rate were measured. Blood samples were taken at each step to measure hemoglobin concentration and hematocrit. Arterial blood oxygen content was also calculated.
Results:
At sea level, a decrease in arterial oxygen saturation to 92.1% ± 2.5% was observed at 150 W and the hematocrit, hemoglobin concentration and oxygen content were not altered. At high altitude, arterial oxygen saturation decreased, reaching 88.2% ± 4.9% at 50 W and remained constant during the entire exercise protocol. Hemoglobin concentration and hematocrit increased reaching 16.4 ± 0.9 g/dL and 48.8% ± 1.6%, respectively, at 100 W and were maintained until recovery. Arterial oxygen content was constant during exercise and increased in the recovery period.
Conclusion:
An increase in hemoglobin concentration during exercise compensates for the decline in arterial oxygen saturation, meanwhile arterial oxygen content remains constant.
Introduction
A
During acute hypoxic exposure (either normobaric or hypobaric), hemoglobin concentration rapidly increases due to a reduction in plasma volume (Gore et al., 1996). However, when exposure to hypoxia becomes chronic, the increased hemoglobin concentration is a result of elevated erythrocyte production (Richalet et al., 1994) stimulated by high expression levels of hypoxia-inducible factor (HIF), leading to increased erythropoietin synthesis (Semenza et al., 2006), elevated hemoglobin concentration, and improvement of oxygen transport in the blood (Mairbaurl, 1994). This increase in erythropoiesis is detectable by the 5th day at altitude and returns to normal levels at the 2nd or 3rd week of exposure (Winslow, 1984).
At sea level, arterial oxygen saturation decreases during maximum exercise caused by reduced diffusion capacity, an altered ventilation perfusion relationship, and hemoglobin's decreased oxygen transport capacity due to acidosis as a result of increased lactate concentration (Dempsey and Wagner, 1999; Nielsen, 2003). Under both normobaric and hypobaric hypoxic conditions, the decline in saturation is greater than normoxia due to lower oxygen availability (Dempsey and Wagner, 1999; Schoene, 2001).
Over the past 30 years, mining activity in Chile has relocated miners who normally live at sea level to work at an altitude >3000 m above sea level. This Chilean model of chronic intermittent hypobaric hypoxia (CIHH) exposure is characterized by alternating periods of work at high altitude and rest periods at sea level (Richalet et al., 2002; Moraga et al., 2014). Previously, several studies have shown long-term effects on the health of these miners during altitude acclimatization (Farias et al., 2006; Brito et al., 2007). Results reported by Richalet et al. (2002) indicated that the hematological response is gradual and characterized by increased hemoglobin and hematocrit concentrations during the first 12–18 months at high altitude (3800–4600 m), with a posterior decrease to values similar to those observed at sea level after 31 months of CIHH exposure. In the same study, as exposure time increased, a progressive decrease in physical performance was observed: explained by a decreased maximum HR due to the downregulation of β-adrenergic and upregulation of muscarinic receptors (Richalet et al., 1992). Also, a study that evaluated maximal aerobic capacity (VO2 max) performed in soldiers with CIHH exposure (6 months) at high altitude (3550 m) showed that these subjects presented decreased VO2 max at high altitude with increased oxygen transport capacity (Prommer et al., 2007).
Therefore, our aim was to evaluate the hemoconcentration response during a maximum exercise test performed at sea level and high altitude in a population of acclimatized miners exposed to CIHH at 3800 m.
Materials and Methods
Sixteen miners after a shift work altitude schedule were recruited. They were informed about the study and voluntarily consented to participate. All the miners were healthy, had moderate physical activity, and had been intermittently exposed to high altitude with alternating work shifts of 7 days at 3800 m and 7 days rest at sea level for between 7 and 36 months. Age (years), weight (kg), and height (m) were recorded, and the body mass index (BMI) was estimated using the relation: BMI = weight/height2 (expressed in kg/m2). All protocols used in this study were in accordance with international ethical guidelines (according to the Helsinki declaration) and approved by the Ethics Committee of the Medical Faculty of the Universidad Católica del Norte, Chile.
Exercise test and blood analysis
The miners performed two maximum exercise tests, one at sea level (in our laboratory) and the other at high altitude (in a clinic located in the mining complex); exercise was performed on a cycle ergometer (Model Corival, Lode) placed at sea level and at high altitude. The protocol consisted of three phases: warm up (5 min) at an intensity of 0 W; exercise test, increasing the load by 50 W every 3 minutes, while maintaining a cadence of 70 rpm, until exhaustion; and a recovery period of 30 minutes. All tests were performed on the 4th day at sea level or at high altitude. During the protocol, transcutaneous arterial saturation (SpO2, %) and HR (bpm) were assessed by a pulse oximeter (8500 M Nonin Medical, Inc.). The sensor was placed on an ear lobe (8000Q2 Nonin Medical, Inc.).
For hemoglobin and hematocrit measurements, blood samples were taken through a catheter inserted into the brachial vein, allowing for blood sample collection during the rest period, at the end of each load (50, 100 150, and 200 W), during the exercise test, and during recovery at 5, 15, and 30 minutes after the end of the test. For each sample, 3 mL of venous blood was collected in a tube with EDTA and analyzed on a hematological counter (Cell-Dyn 1400; Abbott), and hemoglobin concentration (g/dL) and hematocrit (%) were measured. Oxygen content (CaO2) was calculated using the following equation: CaO2 (mLO2/dL) = 1.34 (mLO2/gHb) × Hb (g/dL) × SpO2 (%) (Calbet et al., 2015), where the maximum volume of oxygen that combines with 1 g of hemoglobin is 1.34 and Hb is hemoglobin.
Statistical analysis
Data analyses were performed using GraphPad Prism version 5.03 (GraphPad Software, Inc.) and presented in tables as mean ± standard deviation and figures expressed as box-and-whisker plot. Significant differences were analyzed using ANOVA followed by a Newman–Keuls test. A p-value <0.05 was considered statistically significant.
Results
As shown in Table 1, BMI values indicate that volunteers were overweight. Also, basal hemoglobin concentration was significantly higher at 3800 m than at sea level.
Mean ± standard deviation, *p < 0.05, sea level versus high altitude.
Oxygen saturation (SpO2) during the maximum exercise test is shown in Figure 1. At sea level, SpO2 decreased compared with resting conditions, starting at 150 W and remained low until 200 W, where it then rapidly recovered after 5 minutes. At high altitude, saturation decreased earlier, reaching 88.2% at a load of 50 W and remaining low during the entire test at 200 W; however, it returned to resting values after 5 minutes of recovery.

Effect of increase in load exercise (Watts) on the arterial oxygen saturation (%). R5, R15, and R30 indicate 5, 15, and 30 minutes of recovery after maximum load. Data are shown as box-and-whisker plot at sea level (gray box) or at high altitude (white box). Asterisk represents significant difference with resting period (*p < 0.05).
Hematocrit levels measured at sea level were slightly increased (but not significantly) in proportion with increased exercise intensity (Fig. 2A). However, at high altitude, a significant increase in hematocrit was observed starting at 100 W and remained elevated until 5 minutes of recovery (p < 0.05), decreasing to resting levels after 15 and 30 minutes of recovery.

Hemoglobin concentration during the test (Fig. 2B) at sea level is slightly increased in proportion with increased exercise intensity, although this was not significant. However, a significant increase was observed at high altitudes from 100 W and remained until 5 minutes of recovery, reaching values similar to resting levels at 15 and 30 minutes after maximum exercise.
Finally, no significant alteration in oxygen content was observed with exercise intensity at either sea level or at high altitudes, and values remained similar to resting values during the entire test (Fig. 3). However, increased oxygen content was observed at high altitudes after 5 minutes of recovery, and remained slightly (but not significantly) high after 15 minutes of recovery and reached values similar to resting values after 30 minutes of recovery.
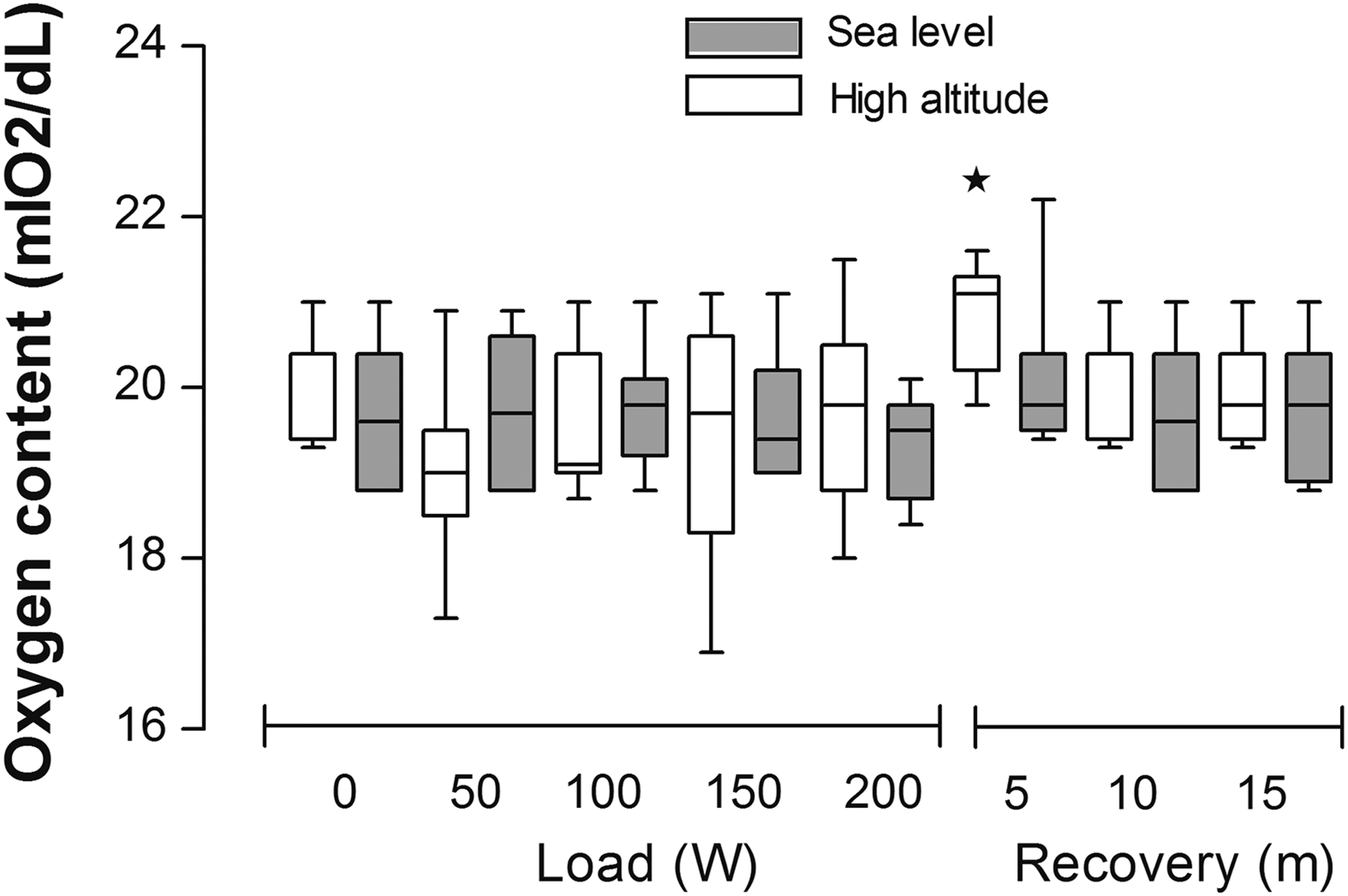
Effect of increase in load exercise (Watts) on the oxygen content (mLO2/dL). R5, R15, and R30 indicate 5, 15, and 30 minutes of recovery after maximum load. Data are shown as box-and-whisker plot at sea level (gray box) or at high altitude (white box). Asterisk represents significant difference with resting period (*p < 0.05).
Discussion
Our results show a rapid and transient increase in hemoglobin concentration at an altitude of 3800 m that directly correlates with exercise intensity. This elevated hemoglobin concentration compensates the fall in arterial oxygen saturation, keeping oxygen content almost stable during the maximum exercise test performed in miners with chronic intermittent exposure to hypoxia at high altitude.
Our study shows a slight elevation in resting hemoglobin concentration values (close to 5%) during normoxic conditions compared with those previously found in subjects evaluated at sea level (Winslow, 1984; Richalet et al., 2002; Heinicke et al., 2003). This difference could be explained by a cumulative erythropoietic response as a result of previous CIHH (7–36 months of exposure). This is similar to results described in a prospective study performed in 29 miners without previous experience at high altitudes, where the hemoglobin concentration at sea level was increased by 10% after 12 months of exposure to CIHH (Richalet et al., 2002). In addition, at high altitudes, the resting hematocrit values observed in our study were 45.9%, which is considered an adequate hematological response to CIHH (Richalet et al., 2002), and meet the requirements of the health ministry (in Chile) that provides a technical guide for high-altitude workers. Therefore, in our country, a hematocrit >55% is not recommended for working at high altitude. (www.minsal.cl/sites/default/files/guia_hipobaria_altitud.pdf).
However, this slight increase in hemoglobin concentration (hemoconcentration), previously described at sea level, becomes significantly elevated (by almost 5%) on the 4th day of arrival at 3800 m. This hemoconcentration is close (4.5% and 4%) to the concentration described in two studies performed with soldiers exposed to 6 months of CIHH, on the 1st and 4th day after arrival at 3550 m (Heinicke et al., 2003; Prommer et al., 2007). This effect was also described in miners without previous experience to high altitudes, where evaluations were performed on the 6th day, during the first exposure at 3800 m. These subjects showed an increased hemoglobin concentration of 13%, which then decreased by 6% after 12 months of exposure to CIHH, and remained at these levels until the 30th month (Richalet et al., 2002). These results, suggest that exposure time to CIHH is key to promoting a minor increase in hemoglobin concentration. This hemoconcentration pattern could be explained by reduced plasma volume and/or cell volume expansion (for more detail, see Siebenmann et al., 2017).
During acute hypoxic exposure (either normobaric or hypobaric), hemoconcentration rapidly increases due to a reduction in plasma volume (Gore et al., 1996) and severity of reduction is given to the altitude reach. In general, reduced plasma volume occurs when there is a negative water balance between intake and loss. At high altitude, reduced voluntary water intake was observed in climbers due to their blunted sensation of thirst and decreased food intake (Westerterp et al., 2000): evidence supported by studies in animal models (Jones et al., 1981). Also, higher respiratory fluid loss resulting from the dry air and the enhanced pulmonary ventilation stimulated by hypoxia and increased diuresis was mediated, in part, by respiratory alkalosis or diuresis controlled by hormones such as aldosterone, atrial natriuretic peptide, and sympathetic activity; along with other mechanisms such as an increased transvascular fluid leakage promoted by systemic inflammation (Siebenmann et al., 2017). The physiological outcome of plasma volume reduction, in acute exposure to high altitude, is a rapid hemoconcentration (Siebenmann et al., 2015).
A second mechanism is the expansion of red cell volume that could be explained by (1) hemoconcentration induced by vascular constriction, by increased sympathetic activity promoted by hypoxia. In agreement, a study carried out in llamas and sheep gradually exposed to high altitude showed that both species displayed a gradual increase in hemoconcentration, suggesting that sympathetic activity is involved in this process (Banchero and Grover, 1972); (2) another possible contributing factor that could explain the hemoconcentration is the release of stored red blood cells from the spleen. Studies have shown that simulated hypoxia or maximal exercise produce contraction of the spleen promoting the release of red blood cells, producing a hemoconcentration (Laub et al., 1993; Richardson et al., 2008, 2009). In both conditions, vascular and spleen contractions are due to increased norepinephrine concentration, studied and corroborated by a pharmaceutical approach using α1 adrenergic blockers, preventing the elevation in hemoglobin and hematocrit which supports an important sympathetic role (Steward and McKenzie 2002; Shepard, 2015); and (3) by increased erythropoiesis mediated by an increase in renal erythropoietin production that is finely regulated by tissue PaO2, stimulating the expression of HIF; leading to increased erythropoietin synthesis (Semenza et al., 2006).
Our study showed that exercise at high altitude induces a hemoconcentration of 9% at maximum exercise with increased exercise intensity and decreased arterial oxygen saturation. Despite the drop in arterial oxygen saturation (close to 10%), the transient increase in hemoglobin concentration is sufficient to maintain arterial oxygen content stability during the exercise test. In addition, when the study was performed at sea level, no hemoconcentration was observed. However, in other studies, a maximum exercise test was performed at sea level and at high altitude in soldiers exposed to CIHH, showing a hemoconcentration response of 10%, respectively (Prommer et al., 2007). This fact strongly suggests that in our subjects, CIHH exposure promotes peripheral vasoconstriction mediated by exercise intensity at high altitude, leading to redistribution of blood flow that favors oxygen delivery to the muscle and hemoconcentration in the vascular compartment. This allows oxygen content to remain almost stable during exercise, reducing tissue hypoxia (Calbet and Lundby, 2009; McArdle et al., 2009). In addition, previous studies carried out in subjects exposed to hypobaric hypoxia support the finding that peripheral vasoconstriction is necessary to maintain oxygen content (Lundby et al., 2008) and studies performed with spectroscopy near the infrared range showed that venous oxygen saturation does not decrease, indicating that increased hemoglobin concentration maintains tissue oxygen output (Costes et al., 1996; Boushel and Piantadosi 2000; Boushel et al., 2001; Calbet and Lundby, 2009).
Exercise-induced arterial hypoxemia was observed at sea level with a fall in the SpO2 of 94% and 92% observed at 150 W and 200 W, respectively. This fall in arterial oxygenation was previously considered “abnormal” (Mitchell et al., 1958). Nowadays, exercise-induced arterial hypoxemia is defined arbitrarily as a reduction in oxygen arterial pressure of 7.5 mmHg and/or oxygen saturation <95% during exercise at sea level (for more detail, see Nielsen, 2003). Arterial hypoxemia could be explained by V/Q mismatch or increased shunt, reduced oxygen availability (high-altitude exposure) (Schoene, 2001), lactic acidosis, etc. In our study, all volunteers often play football (soccer) at high altitudes and at sea level. They are healthy, without respiratory and cardiovascular disease, nobody smoked, and the company healthcare indicated that no one was exposed to pollutants such as carbon monoxide that could have explained the fall in oxygen saturation observed. In this sense, lactic acidosis decreased blood oxygen affinity, facilitating oxygen delivery to peripheral territories. This could be a better explanation for the decrease in arterial oxygenation observed in our study (Nielsen, 2003). Future evaluation of oxygen consumption and lactic concentration in this population exposed to CIHH is necessary.
Limitations
Our study did not perform a noninvasive evaluation of vascular function, such as near infrared spectroscopy or echo Doppler to demonstrate hemodynamic changes in subjects. Future studies considering these types of measurements in a larger population of workers are needed to obtain a better understanding of the mechanisms underlying the high-altitude physiological adjustments in the population of shift workers.
In conclusion, the elevated hemoglobin concentration compensates the fall in arterial oxygen saturation, maintaining oxygen content almost stable during the maximum exercise test performed in miners with chronic intermittent exposure to hypoxia at 3800 m; this increase in hemoglobin concentration during exercise or work at high altitudes could be a protective response against hypoxia.
Footnotes
Acknowledgments
The authors thank our assistant researcher Mr. Hervis Galleguillos and nutrition student Mr. Andres Robles who collaborated in this study. Furthermore, we would like to thank all the miners who voluntarily participated in the study. We are also grateful for the financial support of the project 07 CN13ISM-152 INNOVA CORFO-CHILE.
Author Disclosure Statement
No competing financial interest exist.