
Abstract
Abstract
Heinonen, Ilkka, Olli Vuolteenaho, Juha Koskenvuo, Olli Arjamaa, and Mikko Nikinmaa. Systemic hypoxia increases circulating concentration of apelin in humans. High Alt Med Biol. 18:292–295, 2017.
Background:
Apelin is a hormone that regulates cardiovascular function, and its concentration is increased by hypoxia based on cell culture and animal studies. As it remains unknown as to whether hypoxia could affect apelin levels in humans, we investigated whether breathing normobaric hypoxic gas mixture increases the circulating apelin concentration in healthy male subjects.
Methods:
Ten healthy young men (age 29 ± 5 years, body mass index 24.7 ± 2.8 kg/m2) breathed normobaric hypoxic gas mixture (11% O2/89% N2) for 1 hour. Venous blood samples were obtained immediately before, and 2 and 24 hours after the start of the hypoxic exposure and analyzed for circulating apelin concentrations.
Results:
Arterial oxygen saturation decreased steadily from a baseline value of 99% ± 1% after the initiation hypoxia challenge and reached a steady-state level of 73% ± 6% within 20–30 minutes. Baseline apelin concentration was 3.3 ± 1.3 pmol/L and remained comparable (3.3 ± 1.4 pmol/L) to the baseline concentration at a 2-hour time point. However, apelin concentration at the 24-hour time point (5.5 ± 2.8 pmol/L) was significantly (by ∼67%) higher as compared with at both baseline and 2-hour time points (p < 0.05).
Conclusion:
In conclusion, in line with cell culture and animal studies, acute systemic hypoxia increases circulating apelin concentration also in humans.
Introduction
A
Methods
Subjects
Ten healthy Caucasian young men (age 29 ± 5 years, height 181 ± 4 cm, weight 81 ± 11 kg, body mass index 24.7 ± 2.8 kg/m2) volunteered to participate in this study. The purpose, nature, and potential risks of the study were explained to the subjects before they gave their written informed consent to participate. The subjects were normotensive nonsmokers with no history of hypercholesterolemia, not taking any medications, and had never experienced angina symptoms. The normal health status of the subjects was confirmed by a medical doctor by clinical examination, electrocardiography (ECG), and cardiac echocardiography before the experiments. The study was performed according to the Declaration of Helsinki and was approved by the Ethics Committee of the Hospital District of Southwest Finland.
Hypoxic exposure and blood sampling
The subjects were requested to avoid strenuous physical exercise for the 48 hours before the experiments. The hypoxic exposure was performed in a fasting state and always started at around 9AM with study preparations. In these preparations, a 12-lead ECG was positioned and an antecubital vein was cannulated for blood sampling. Venous blood samples were obtained before, and 2 and 24 hours after the start of the hypoxic exposure and analyzed for apelin as described later. After initial preparations, the subjects breathed normobaric hypoxic gas mixture (11% O2/89% N2) for 1 hour. Arterial oxygen saturation was continuously measured by pulse oximetry, and valid plethysmogram signal was confirmed throughout the studies. All of these measurements were performed when subjects were resting supine.
Analysis of circulating apelin concentration
The ethylenediaminetetraacetic acid (EDTA) plasma samples (1 mL) were extracted with SepPak C18 cartridges. The recovery of 50 fmol of synthetic apelin-13 added to 1 mL plasma and extracted with SepPak C18 was 69% ± 9% (mean ± standard deviation [SD], n = 13). Apelin concentrations in the plasma extracts were measured with a specific radioimmunoassay developed by us and described here. A specific goat antiserum was raised against human preproapelin66–77 (apelin-12). The tracer in the assay was Preproapelin62–77-Tyr77 (apelin-16 with tyrosine added to the COOH-terminus), radioiodinated with chloramine-T and purified by reversed-phase high-performance liquid chromatography (HPLC) as previously described (Ala-Kopsala et al., 2004). Synthetic human preproapelin65–77 (pyroGlu-apelin-13) was used as the calibrator. The incubation procedure was as follows: The calibrators and samples (100 μL) were incubated overnight at +4°C with a dilution of the antiserum (100 μL), resulting in 40% binding of the tracer in the absence of a competitor. Then, ∼10,000 cpm of the tracer (100 μL) was added and the incubation was continued for another 18 hours at +4°C. The bound and free fractions were separated by double antibody precipitation followed by centrifugation. The precipitates were counted for radioactivity in a Wallac CliniGamma gamma counter. The sensitivity of the assay was 0.4 fmol/tube, and the within-assay and between-assay coefficients of variation were <10% and <15%, respectively. The antiserum cross-reacts 100% with apelin-12, apelin-13, apelin-16, and apelin-36, but it does not recognize preproapelin68–77 (apelin-10), atrial natriuretic peptide (ANP), brain natriuretic peptide (BNP), C-type natriuretic peptide (CNP), endothelin-1, or adrenomedullin (cross-reaction <0.1%). Apelin-10, apelin-12, apelin-13, and apelin-16 were synthesized by us with an ABI 433A Peptide Synthesizer using Fmoc chemistry and purified by reversed-phase HPLC. Apelin-36 was purchased from Phoenix Europe GmbH (Karlsruhe, Germany).
Statistical analysis
Statistical analysis was performed by using GraphPad Prism 4.3. One-way analysis of variance (ANOVA) for repeated measurements was used for the analysis of statistical differences. ANOVA was followed by Bonferroni post hoc test if necessary to detect differences in different study time points. p Values <0.05 were considered statistically significant. All data are shown as mean ± SD.
Results
As previously reported (Heinonen et al., 2014b), all of the subjects had normal baseline arterial oxygen saturation of 99% ± 1% at baseline, which decreased steadily after the initiation of hypoxic breathing and reached a stable steady-state level (73% ± 6%) within 20–30 minutes after the initiation of hypoxic exposure. This level of oxygen saturation was maintained until the removal of the hypoxic mask after 1 hour, after which saturation quickly reached normal, pre-hypoxic values (99% ± 1%).
Baseline apelin concentration was 3.3 ± 1.3 pmol/L, and it remained unchanged at the 2-hour time point after the start of the hypoxic exposure (Fig. 1). Apelin concentration at the 24-hour time point was, however, significantly higher as compared with both baseline and 2-hour time points (p < 0.05; Fig. 1).
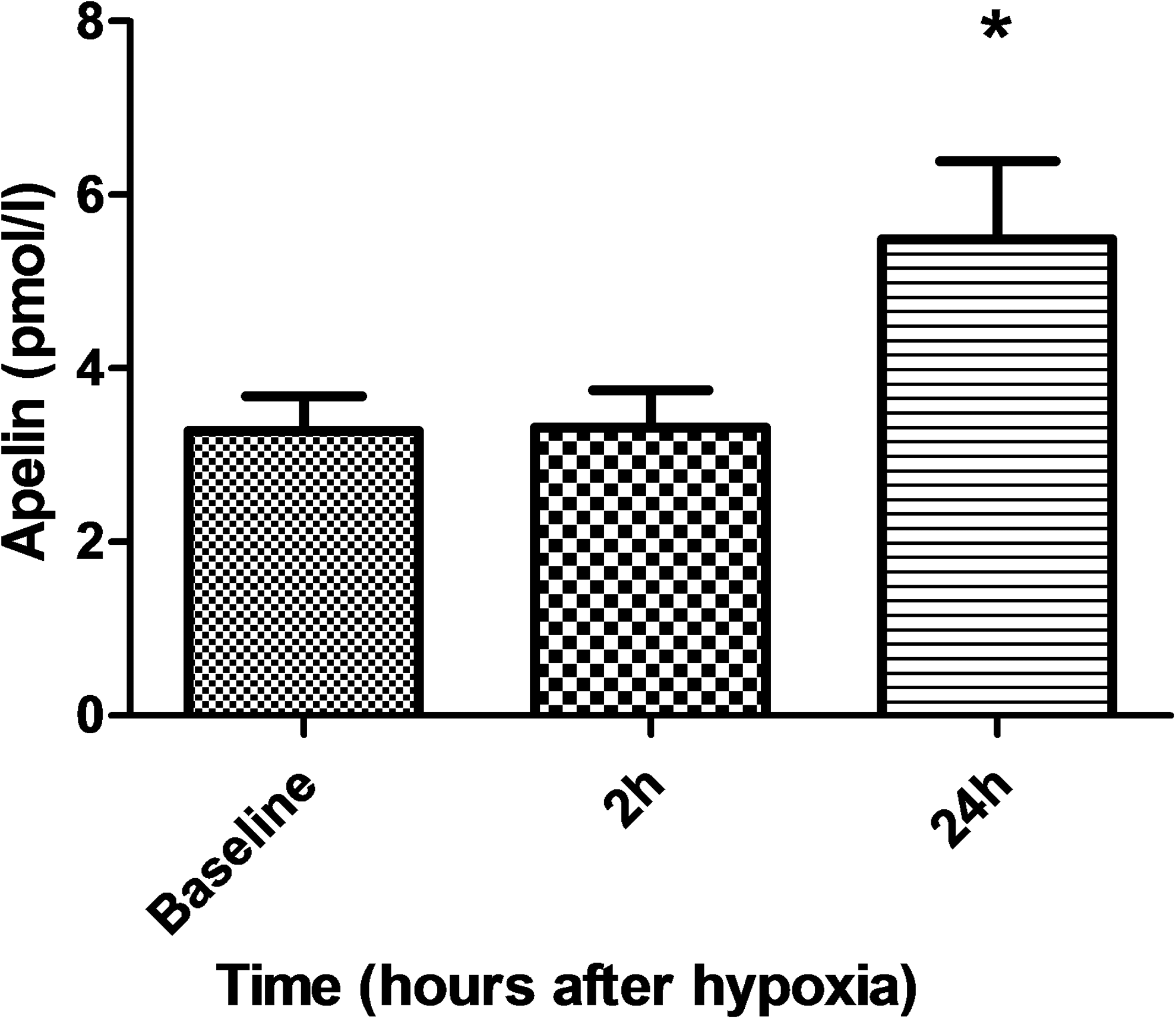
Circulating apelin concentration in human mixed venous blood before hypoxia (baseline) and 2 and 24 hours after the start of the exposure. *p < 0.05 compared with baseline and 2-hour time points.
Discussion
In this study, we repeatedly measured circulating apelin concentrations and could demonstrate that apelin was increased 24 hours after a 1-hour systemic hypoxic exposure in humans. This is in line with the previous experimental studies investigating apelin in response to hypoxia in cells and animals in a laboratory setting.
The animal and cell culture studies have elucidated that activation of apelin is triggered in response to hypoxia by hypoxia-inducible factor-related mechanisms (Ronkainen et al., 2007; Eyries et al., 2008; Sheikh et al., 2008; Kasai et al., 2010; Geiger et al., 2011; Zhang et al., 2016), after which it contributes to new blood vessel formation by triggering the proliferation of endothelial progenitor cells, among its other functions. Apelin concentration tends to be higher in high-altitude natives and lower in patients who suffer from high-altitude pulmonary edema compared with those who do not develop the disease at a high altitude (Mishra et al., 2015); however, to the best of our knowledge, its increase has not yet been shown experimentally in a well-controlled setting in hypoxic humans. In detail, in this study, in response to 1-hour fairly severe systemic hypoxia, apelin concentration remained similar to baseline levels 2 hours after the hypoxic period, but it increased markedly (by some 67%) 24 hours after the hypoxic exposure. This pattern of release suggests that apelin is not stored in the cells, but the expression of apelin is triggered by hypoxia and increased peptide concentration can be seen first several hours after the condition change inducing the apelin expression has occurred, because of the inherent time lag between the initiation of transcription and the resultant translation of mRNA to the protein product. We could not unfortunately take blood samples, for instance, at 12 hours after the hypoxia exposure because of logistic reasons, and, therefore, it is not known whether apelin concentration increased already earlier than after 24 hours after the exposure.
As our samples were collected from mixed venous blood, the cell source of apelin cannot be partitioned to one or even a few cell types in this study. Previous cell culture and animal studies, however, strongly suggest that apelin affects mainly cardiac myocytes and is released not only mostly from them but also from vascular smooth muscle and endothelial cells (Kawamata et al., 2001; Kleinz and Davenport, 2004; Kleinz et al., 2005). Interestingly, in addition to the cardiovascular system, there is an expression of apelin even in adipocytes, which is triggered by insulin, and apelin levels have been shown to be increased in obesity (Boucher et al., 2005).
In addition to its direct positive effects on cardiac function, apelin has been shown to increase the release of nitric oxide, which has many important functions in peripheral vasculature (Heinonen et al., 2011, 2013) and in the heart (Seddon et al., 2007; Simon et al., 2014). Consequently, apelin may be one of the key mediators regulating myocardial function and circulation, which are positively affected by regular physical activity (Heinonen et al., 2008, 2014a). Nevertheless, it remains to be shown as to whether circulating apelin concentration is higher in physically active subjects, or whether acute physical exercise can also increase apelin concentration. It has, however, been shown in numerous studies that hypoxic exposure has many positive effects on human cardiovascular function and metabolism (Heinonen et al., 2016). Therefore, as apelin is increased in hypoxia as demonstrated in this study, hypoxia is an interesting therapy option to improve human health, possibly through apelin-mediated mechanisms. In addition, apelin itself serves as a potential drug development target (Kleinz and Davenport, 2005; Gerbier et al., 2016).
In conclusion, in accordance with cell culture and animal studies, acute systemic hypoxia increases circulating apelin concentration also in humans as assessed in a well-controlled experimental hypoxic setting. As both apelin and hypoxia have many positive physiological effects on health, acute systemic hypoxia, or natural altitude exposure, they could serve as an alternative or adjunct therapy option to physical activity and other healthy activities to improve the function of the cardiovascular system.
Footnotes
Acknowledgments
The study was conducted within the Centre of Excellence in Molecular Imaging in Cardiovascular and Metabolic Research—supported by the Academy of Finland, University of Turku, Turku University Hospital, and Åbo Akademi University. The authors want to thank the personnel of the Turku PET Centre for their excellent assistance. The study was financially supported by the Academy of Finland (grant nos. 251572 and 258078, and Centre of Excellence funding), and the Hospital District of Southwest Finland.
Author Contributions
All authors have participated in the study planning, performing, and writing of this article. All authors have read and approved submission of the final version of this article.
Author Disclosure Statement
No competing financial interests exist.