
Abstract
Abstract
Sheth, Chandni, Hendrik Ombach, Paul Olson, Perry F. Renshaw, and Shami Kanekar. Increased anxiety and anhedonia in female rats following exposure to altitude. High Alt Med Biol. 19:81–90, 2018.—Anxiety disorders are chronic, highly prevalent conditions, often comorbid with depression. Both anxiety and depression form major risk factors for suicide. Living at altitude is associated with higher rates of depression and suicide, leading us to address whether anxiety disorders may also be amplified at altitude. Using a novel translational animal model, we previously showed that depression-like behavior increases with altitude of housing in female, but not male rats. We now use this model to examine the effects of altitude on both anxiety-like behavior and anhedonia, a core symptom of depression. After housing for a week at sea level, 4500 or 10,000 ft, rats were evaluated for anxiety in the open-field test or the elevated plus maze, and anhedonia in the sucrose preference test. Another group was tested at baseline. Anxiety-like behavior increased in females housed at altitude. In females, lower sucrose preference was seen in those housed at 10,000 ft versus those at sea level. Males showed no change in anxiety or anhedonia across groups. These data suggest that living at moderate-high altitude may pose a risk factor for those vulnerable to anxiety disorders, with the potential to be particularly detrimental to females at altitude.
Introduction
A
Anxiety disorders exhibit significant comorbidity with depression, suggesting that the two disorders may share a similar underlying pathophysiology (Hirschfeld, 2001). Disruption of serotonin function has been largely implicated in both depression and anxiety disorders (Albert et al., 2014). Interestingly, there is substantial evidence supporting sex differences in prevalence, with females showing a higher preponderance of depression and anxiety disorders than males (Kessler, 2003; Seedat et al., 2009). Of particular importance, anxiety and depression are established risk factors for suicide (Kanwar et al., 2013; Bentley et al., 2016).
Suicide is the 10th leading cause of death in the United States and exerts a significant economic and social burden (World Health Organization, 2014). In recent years, living at altitude has been established as an independent risk factor for suicidal behavior: rates of suicide increase significantly with altitude of residence, independent of varying cultural and socioeconomic factors (Brenner et al., 2011; Helbich et al., 2013; Asiri, 2014; Huber et al., 2014; Kim et al., 2014; Alameda-Palacios et al., 2015; Zaeh et al., 2016). Major depressive disorder (MDD), a major risk factor for suicide, is also found to increase with altitude of residence (DelMastro et al., 2011). Residing at altitude exposes individuals to chronic hypobaric hypoxia (low partial pressure of oxygen), which may alter brain physiology to contribute to the higher rates of depression and suicidal behavior at altitude.
In agreement with human studies, preclinical investigations also demonstrate a link between altitude and depression. For example, we have previously shown an altitude-dependent increase in depression-like behavior (DLB) in female, but not male rats (Kanekar et al., 2015) using the forced-swim test (FST). In other studies, rats exposed to chronic hypobaric hypoxia (simulating moderate to extremely high altitudes) exhibit significantly reduced brain levels of serotonin (Ray et al., 2011), which plays a crucial role in mood and emotion regulation (Zangen et al., 1997). Affective disorders have been increasingly considered to be disorders of brain energy metabolism (Moretti et al., 2003), and hypobaric hypoxia exposure is also found to disrupt brain energetics. A drop in prefrontal cortex brain creatine was observed in female rats after a week of housing at 10,000 ft simulated altitude (Bogdanova et al., 2014). Thus, reduced serotonin levels coupled with aberrant bioenergetics in the brain may provide a neurobiological basis for the link between altitude and higher incidence of depression and suicide. Since both depression and anxiety are linked to dysfunction of both serotonin and brain bioenergetics (Konradi and Ongur, 2017; Shimamoto and Rappeneau, 2017), we investigated whether anxiety behaviors may also be impacted by altitude of residence.
Research investigating the effects of altitude on anxiety is relatively scarce. Anecdotal evidence suggests a link between altitude exposure and anxiety in humans (Nicolas et al., 1999; Sracic et al., 2014; Risal et al., 2016). In one study, climbers participating in a 31-day period of gradual decompression in a hypobaric chamber simulating to a climb from sea level to Mount Everest (29,000 ft) exhibited an increase in state anxiety as evaluated by the Spielberger's State-Trait Anxiety Inventory (STAI) (Nicolas et al., 1999). Anxiety levels may also be exacerbated by living at moderate to moderate high altitude (4000–12,000 ft). A study by Risal et al. (2016) evaluated depression and anxiety levels in 2100 participants in Nepal using the Hospital Anxiety and Depression Scale (HADS) and found that living above 6560 ft of altitude (2000 m) increased odds ratios (ORs) for depression (OR 1.9), anxiety (OR 1.2), and comorbid depression/anxiety (OR 2). Prevalence of mood disorders rose with altitude of residence: anxiety levels rose from 15.5% at below 6560 ft to 18.5% above 6560 ft, depression from 4.4% to 8.1%, and comorbid depression/anxiety from 5.5% to 10.2% (Risal et al., 2016). Studies imply that both chronic and acute effects may be seen with altitude exposure in humans. A cohort of seven Marines traveling from sea level to altitudes between 6761 and 11,099 ft reported spontaneous acute onset anxiety within the first 6 days of arrival at altitude, suggesting that acute hypoxic exposure may also increase anxiety (Sracic et al., 2014). However, in rodent models, a link between anxiety and altitude has only been demonstrated in the extreme altitude range. After exposure to an altitude simulation of 25,000 ft for 2 weeks, rats showed increased anxiety-like behavior (Baitharu et al., 2013). Since human studies suggest that anxiety effects may be seen at much more moderate altitudes than the 25,000 ft tested in rats, we chose to investigate the effect of housing at moderate altitudes on anxiety-like behavior in our animal model for hypobaric hypoxia.
Animal models have been used for decades to investigate the etiology of mood disorders and evaluate the impact of stressors. Of particular interest, animal models have recently been used to evaluate anxiety, depression, and anhedonia, specifically with reference to impact on suicidal behavior (Gould et al., 2017). In our study, anxiety-like behavior was evaluated using two tests: the elevated plus maze test (EPM) and the open-field test (OFT). Both the OFT and the EPM are based on a rodent's natural aversion to open spaces, and are highly validated models for testing anxiety-like behavior in rodents (Prut and Belzung, 2003). Although short and deceptively simple, both have demonstrated good face, construct, and predictive validity in decades of use for assaying anxiety-like behavior in rats (Prut and Belzung, 2003; Walf and Frye, 2007). In the OFT, rats when placed in an open arena prefer to stay in the periphery (along walls or in corners) and avoid the more well-lit, open center. Less time spent in the center of the OFT chamber is indicative of increased anxiety (Hiroi and Neumaier, 2006). The EPM is a cross maze with two open arms and two closed arms. Behavior in the EPM is reflective of the conflict between a rat's preference for protected areas (closed arms) and the innate motivation to explore novel environments (open arms) (Pellow et al., 1985). Increased preference for the closed arms of the EPM represents higher anxiety levels. The simplicity of the EPM allows it to be used in combination with other behavioral tests for mood disorders, in this case, the sucrose preference test (SPT) for anhedonia. Anhedonia is a core symptom of depression, defined as the inability to feel pleasure in response to pleasurable stimuli. The SPT is a highly validated two-bottle preference test, widely used for decades to consistently measure anhedonia in rodents (Papp et al., 1991). The SPT measures a rat's innate preference for a sweet palatable drink (sucrose solution) over water (Overstreet, 2012). A bias toward a sweet rewarding drink is typical, and failure to develop this bias is indicative of anhedonia (Papp et al., 1991).
The purpose of this study was to investigate whether exposure to moderate altitude conditions (4500 and 10,000 ft) could increase levels of anxiety and anhedonia in an animal model. We have established a novel translational animal model of hypobaric hypoxia and used it to show that female rats exhibit incrementally greater DLB with increase in altitude of housing (Kanekar et al., 2015). Given that depression and anxiety have overlapping neurobiological and clinical signatures, and that anhedonia is a core symptom of depression, we hypothesized that, like depression, anxiety and anhedonia might increase with altitude of housing in our animal model.
Methods
Animals
For all experiments, male and female Sprague Dawley (SD) rats, approximately 125–150 g in body weight (6–7 weeks in age), were received from Charles River Laboratories (Raleigh, NC). Animals were housed individually in 20 × 40 × 20 cm polycarbonate cages and given food and water ad libitum. All animal procedures were approved by the University of Utah Institutional Animal Care and Use Committee and the VA ACORP, and were performed in accordance with the NIH Guide for Care and Use of Laboratory Animals. To ensure that females did not face any additional stresses to that experienced by males, vaginal testing for estrous cycle stage was not conducted (Allen et al., 2010; Kanekar et al., 2015).
Experimental groups
Animals were randomly assigned to four study groups: three altitude groups—sea level (Experimental group: SL), 4500 ft (Experimental group: 4.5K) or 10,000 ft (Experimental group: 10K), or a baseline group (Experimental group: day 0 or D0). Animals spent a week in acclimatization immediately after delivery, and were then put into altitude chambers (SL, 10K groups), or at local elevation (4.5K group) for 1 week. The D0 group was studied at the end of the week of acclimatization, to provide baseline values for status before the week of altitude exposure. All rats in the baseline group were thus tested for behavior at 7–8 weeks of age, while those in the altitude groups were behaviorally tested at 8–9 weeks of age.
Altitude simulations
Hypobaric hypoxia or hyperbaric chambers were used to reduce or increase pressure from ambient barometric pressure at the University of Utah Research Park (4500 ft) to simulate high-altitude conditions or sea level, respectively. Each altitude chamber consists of a compression cylinder (5.2 ft long, 1.8 ft in diameter) that easily fits six standard rat cages at a time. Rats in the SL group were placed in the hyperbaric chamber set to simulate sea level conditions (21% ppO2, 760 mmHg), while those in the 10K group were housed in hypobaric hypoxia chamber set at 523 mmHg (15% ppO2). The 4.5K group was housed at local elevation (Salt Lake City, UT: 18% ppO2, 644 mmHg) adjacent to the altitude chambers.
Elevated plus maze
After a week in different altitude groups, a group of rats were tested for anxiety-like behavior in the EPM at local conditions, as per the protocol from previously published literature (Walf and Frye, 2007). One hour before EPM testing, rats were moved to the testing room to habituate. The EPM is a cross maze with two closed arms and two open arms, elevated 92 cm off the ground. Each arm is 50 cm long and 10 cm wide, with 41 cm high walls on closed arms and no walls on open arms. For the EPM, a rat was placed in the center of the maze facing the same open arm, and allowed to roam freely for 5 minutes. The EPM was performed in red light (∼10 lux) at the same time of the day/light cycle (2–4 pm), with animals visually blocked off from experimenters. Behavior in the EPM is videotaped for later scoring.
Videos were manually scored by two independent researchers, with high reliability between scorers. Behavior on the EPM was scored as time spent in open arms, closed arms, or in the central square (Walf and Frye, 2007). Animals were considered to be in an arm when all four limbs were on the arm. The number of entries into open arms or closed arms was also scored. Two measures of anxiety-like behavior were used for EPM analysis: time spent in closed arms and number of entries into closed arms (Walf and Frye, 2007). The percentage of time spent in the closed arms was calculated relative to the total time of the test. The percentage of closed arm entries was calculated relative to total number of entries into both arms. More time spent in closed arms and a higher number of closed arm entries in the EPM are considered to be indicative of greater anxiety (Pellow et al., 1985).
Open-field test
A separate cohort of rats was tested in the open field for anxiety-like behavior at baseline (D0) or after a week at different altitudes, as previously described (Allen et al., 2010; Kanekar et al., 2015). The rat was placed in the center of the square OFT apparatus (50 × 50 × 50 cm) divided into 5 × 5 cm2 areas in a grid. The center of the field was illuminated by a 60-W bulb. Rats were allowed to explore the open field for 5 minutes. Behavior was recorded and later manually scored. Rodent behavior in the OFT was scored for time spent in the center and the periphery. The periphery was defined as 64 blocks of 5 × 5 cm grids from the walls of the OFT apparatus, while the inner 36 blocks of 5 × 5 cm blocks were considered to be the center of the apparatus (Seibenhener and Wooten, 2015).
Sucrose preference test
Immediately following the EPM, rats were tested for anhedonia in the SPT. In the SPT, a rat is presented with a bottle of a sucrose solution (1% in tap water) and one with water, and preference to drinking sucrose solution is measured (Overstreet, 2012). The SPT is conducted at local conditions (4500 ft), and sucrose and water intake were measured by weighing bottles before and after a 24-hour period. Sucrose preference was calculated as a percentage of sucrose intake over total fluid intake over 24 hours (Papp et al., 1991).
Statistical analysis
Statistical analysis was performed using GraphPad Prism software (La Jolla, CA). Data were analyzed using one-way analysis of variance (ANOVA) to examine effects of altitude on the dependent variables. Holm-Sidak multiple comparison tests were conducted to determine statistical significance with corrected p-values. Data are presented as mean ± standard error of the mean. Outliers were determined in the EPM as those that spent greater than 150 seconds in the center square as an indication of ambivalence. Furthermore, for EPM, OFT, and SPT, values that were 1.5 interquartile ranges above the third quartile or below the first quartile were considered statistical outliers (Tukey, 1977). Male and female rat data were analyzed separately to avoid any potential confounding effects due to sex, since rodent behavior (Simpson et al., 2012) and the impact of hypobaric hypoxia on rodent behavior (Kanekar et al., 2015) are shown to differ significantly by sex.
Results
Effects of altitude on anxiety in the EPM
In female rats, one-way ANOVA showed a significant main effect of altitude on percentage of time spent in closed arms [F(3,62) = 5.2, p = 0.003]. Holm-Sidak tests revealed that female rats housed at 4500 ft (p = 0.01) or 10,000 ft (p = 0.02) spent significantly more time in closed arms compared to the D0 group. Females in the 4.5K or 10K groups also showed a trend toward significance versus the SL group (p = 0.06 for both comparisons). However, there were no significant differences between females housed at 4500 ft versus those at 10,000 ft (Fig. 1A, n = 16–17 per group).
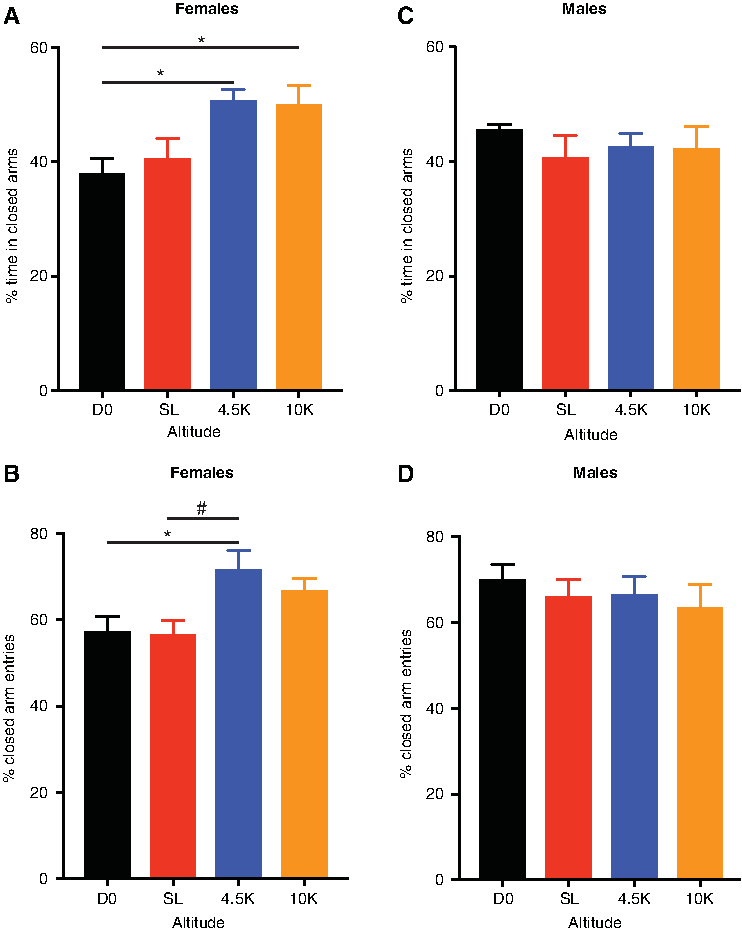
The effects of altitude on anxiety-like behavior in the EPM.
In females, housing at altitude also had a significant effect on percentage of entries into closed arms versus all entries [F(3,61) = 4.15, p = 0.01]. Holm-Sidak tests showed that female rats in the 4.5K group had a higher percentage of closed arm entries compared to the D0 (p = 0.03) and SL groups (p = 0.03). Differences between the 10K group versus the SL or D0 groups did not survive correction for multiple comparisons (Fig. 1B, n = 14–18 per group). However, Student's t-test showed statistically significant differences between the 10K and SL groups (t = 2.4, p = 0.02), as well as the 10K and D0 groups (t = 2.1, p = 0.04), with the 10K group exhibiting significantly more closed arm entries.
In contrast, in male rats, housing at altitude did not have a significant effect on anxiety-like behavior in the EPM, as seen in either time spent in closed arms [F(3,47) = 0.43, p = 0.73] (Fig. 1C, n = 11–16 per group) or in closed arm entries, [F(3,59) = 0.41, p = 0.75] (Fig. 1D, n = 13–17 per group).
Effects of altitude on anxiety in the OFT
In female rats, one-way ANOVA showed a significant main effect of altitude on percentage of time spent in center [F(3,26) = 4.6, p = 0.01]. Holm-Sidak tests showed that female rats housed at 4500 or 10,000 ft had significantly lower percentage of time in the center compared to the SL group (p < 0.05 for both comparisons). Although the comparison between D0 and 4.5K or 10K groups did not survive corrections for multiple comparisons, Student's t-test revealed a significantly lower percentage of time spent in center in the 4.5K and 10K groups compared to the D0 group (p < 0.05 for both comparisons) (Fig. 2A, n = 6–10 per group).
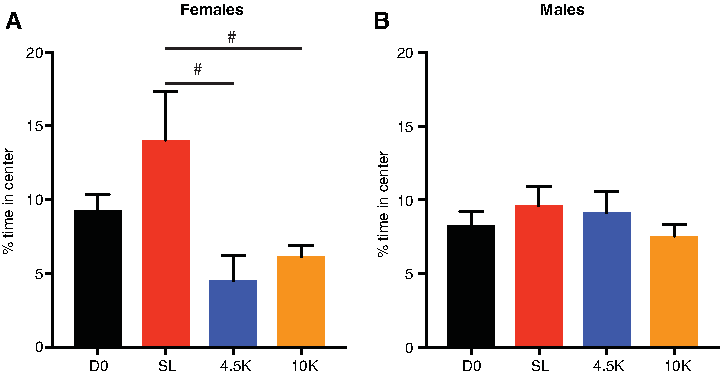
The effects of altitude on anxiety-like behavior in the OFT.
In male rats, one-way ANOVA revealed no significant effect of altitude on percentage of time spent in center [F(3,29) = 0.7, p = 0.54] (Fig. 2B, n = 6–10 per group).
Effects of altitude on anhedonia in the SPT
A significant main effect was seen of altitude on sucrose preference in female rats [F(3,61) = 3.35, p = 0.02]. Post hoc tests showed that the 10K group had significantly lower sucrose preference compared to the SL group (Fig. 3A, n = 16–17 per group). The difference between 4.5K and 10K did not survive correction for multiple comparisons, but a t-test showed a significant difference between the two groups, with the 10K group demonstrating lower sucrose preference than the 4.5K group (p < 0.05).

The effects of altitude on anhedonia in the SPT.
In male rats, housing at altitude for a week did not have a significant effect on sucrose preference [F(3,67) = 1.7, p = 0.17] (Fig. 3B, n = 17–18 per group).
Discussion
In this study, we present evidence that a week of housing of female rats at moderate altitude (4500 and 10,000 ft) increases anxiety-like behavior, as measured in both the EPM (Fig. 1) and the OFT (Fig. 2). In the EPM, higher anxiety-like behavior was seen in female rats housed at 4500 or 10,000 ft compared to the D0 group, as well as a trend toward increased anxiety versus the SL group (Fig. 1A, B). In the OFT, female rats housed at 4500 and 10,000 ft show increased anxiety compared to the SL group, as well as a trend toward increased anxiety versus the D0 group (Fig. 2A). In addition, significantly lower sucrose preference was seen in female rats in the 10K group versus the SL group (Fig. 3A), suggesting that anhedonia, a core symptom of depression, may also increase with altitude of housing in females. However, in striking contrast, male rats did not vary in their anxiety-like behavior in the OFT (Fig. 2B) or EPM (Fig. 1C, D), or in anhedonia in the SPT (Fig. 3B) from baseline values, regardless of altitude of housing. Housing at moderate altitude thus appears to be detrimental to the status of both anxiety and anhedonia in female SD rats, but not in males.
In a previous study, we used the FST to assess DLB in both female and male rats after a week of housing rats at sea level, 4500 or 10,000 ft. Similar to this study, females showed an incremental increase in DLB in the FST with altitude of housing, but males did not (Kanekar et al., 2015). Data from these studies together imply that females may be more vulnerable to hypobaric hypoxia-induced brain deficits, leading to significant changes in anxiety in the EPM and OFT, DLB in the FST, and anhedonia in the SPT.
In addition to studying the impact of housing at different levels of altitude, this study also examines a time course of altitude effects: rats in the D0 group are housed at local conditions (4500 ft) for a week of acclimatization, while those in the 4.5K group are housed for an additional week at local conditions. Female rats housed for 2 weeks at 4500 ft (4.5K group) exhibit significantly increased anxiety in the EPM, as well as a trend toward increased anxiety in the OFT, compared to those housed for 1 week at 4500 ft (D0 group). However, no temporal impact was seen in anhedonia in the SPT: sucrose preference did not vary between the D0 and 4.5K groups in female rats. While effects of altitude on anxiety-like behavior in females manifests at an altitude of 4500 ft in both the OFT and EPM in this study, the effects on anhedonia are only evident in those housed at 10,000 ft. Despite sharing several underlying neurobiological mechanisms, there is evidence that anxiety disorders and anhedonia also have certain distinct nonoverlapping neurobiological underpinnings (Ritov et al., 2016). Thus, it is plausible that exposure to 4500 ft alters neurobiological correlates that underlie anxiety, but not anhedonia.
Currently, there is very little in the literature regarding altitude effects on anxiety-like behavior in animal models. A previous study examined the effects of extreme high-altitude exposure (25,000 ft) for 3–21 days on behavior in the EPM solely in male SD rats (Baitharu et al., 2013). In this study, male rats exposed to a simulated altitude of 25,000 ft for 3 or 7 days showed significantly more anxiety in the EPM compared to the normoxic control group exposure. However, rats exposed to altitude for 21 days were not significantly different than normoxic controls with regard to behavior on the EPM (Baitharu et al., 2013), suggesting that recovery may occur over time in anxiety-like behavior (and potentially brain neurochemistry) in male rats at extremes of high altitude. In this study, male rats also exhibit an increase in anhedonia in the SPT, which persisted after 3, 7, and 21 days at 25,000 ft (Baitharu et al., 2013). These results suggest that, while 7–14 days of moderate altitude exposure may not affect anxiety or anhedonia in male SD rats in our model, exposure to higher altitudes may impact males to worsen these mood disorders. What point in this altitude range triggers behavioral deficits in males remains to be established, but this threshold appears to be higher for males than for female SD rats, suggesting a sex-specific bias in vulnerability to hypobaric hypoxia.
A sex-specific bias in prevalence of anxiety and depressive disorders is well documented clinically: both anxiety disorders and MDD are twice as common in women as in men (Kessler et al., 1993; Kessler, 2003; Altemus et al., 2014). Sexual dimorphism in the serotonin system may contribute to this difference in vulnerability (Donner and Lowry, 2013). In healthy adults, whole brain serotonin synthesis is 52% lower in women versus men (Nishizawa et al., 1997). Women also exhibit higher levels of the major serotonin metabolite 5-hydroxyindole acetic acid (5-HIAA) in the cerebrospinal fluid versus men (Rubinow et al., 1998). In addition, lower serotonin receptor binding has also been identified in the brain of women versus men (Rubinow et al., 1998), together suggesting that differences in the synthesis and metabolism of serotonin may contribute to higher prevalence of anxiety and depressive disorders in females.
In rats, sexual dimorphism is also seen in brain serotonin synthesis, tissue levels, and receptor binding in a brain region-dependent manner (Rubinow et al., 1998). Sex differences in the serotonin system may thus partly account for the higher prevalence of anxiety and depression in females. This study and our previous study (Kanekar et al., 2015) demonstrate that female rats are more vulnerable to a hypobaric hypoxia-induced increase in anxiety and DLB, suggesting that this may also be true for women residing at altitude.
Hypobaric hypoxia-induced changes in brain physiology, which may cause exacerbated anxiety and anhedonia, are currently unknown; however, an altitude-related reduction in brain serotonin levels may be involved. In previous studies conducted on male rats, exposure to extremes of high altitude (2 weeks at 25,000 ft) was found to reduce serotonin levels in the frontal cortex and brainstem (Ray et al., 2011), while 24 hours at 23,000 ft reduced serotonin levels in the striatum and hypothalamus by 34% (Prioux-Guyonneau et al., 1982). Exposure to chronic hypoxia (10% O2 for 14 days) severely reduces rat brain serotonin in a brain region-specific manner (Poncet et al., 1997). Hypoxia-induced alterations in serotonin levels may be a complex interplay between effects of hypoxia on synthesis and metabolism of serotonin. For example, hypoxia was found to decrease serotonin synthesis by reducing activity of the oxygen-dependent serotonin-synthesizing enzyme, tryptophan hydroxylase 2 (TPH2) (Kumar, 2011). Furthermore, even acute exposure to hypobaric hypoxia (23,000 ft for 3 hours) or intermittent exposure can disturb activity of the primary serotonin-metabolizing enzyme, monoamine oxidase (MAO) (Trouvin et al., 1986; Lam et al., 2017). In our model, a significant decrease in serotonin was seen in the female striatum after a week at 4500 or 10,000 ft from baseline (D0), while no change was seen in males (Sheth et al., article in preparation). Given that the striatum is an important neuroanatomical correlate of anxiety and anhedonia (Der-Avakian and Markou, 2012; Marazziti et al., 2015), this finding may provide a mechanistic explanation for the sexually dimorphic effects of altitude/hypoxia on anxiety and anhedonia. These data suggest a sex-dependent difference in the impact of altitude on brain serotonin, with males potentially having a higher trigger point for hypobaric hypoxia-induced brain neurochemical changes than females.
Aberrant serotonergic neurotransmission at altitude may not be the sole mechanism for deficits in anxiety disorders, depression, and anhedonia observed in our model. Hypobaric hypoxia exposure can also alter other brain neurotransmitter systems, including dopamine, norepinephrine, and glutamate (Kumar, 2011; Ray et al., 2011), which may also increase vulnerability to mood disorders at altitude. In addition, hypoxia exposure can be detrimental to brain cellular stress and inflammation, and has been documented to increase reactive oxygen species (Lam et al., 2017), inflammatory markers (Lam et al., 2017), and stress hormones (Baitharu et al., 2013), which may also potentially contribute to the behavioral deficits seen at altitude.
In other animal models (Page et al., 1999; Durkin et al., 2008; Guzzetti et al., 2008; Kulikov et al., 2011), low brain serotonin has been linked to loss of antidepressant function of the selective serotonin reuptake inhibitors (SSRIs), citalopram, fluoxetine, and paroxetine. In our model, we similarly observe a loss of antidepressant efficacy of these SSRIs in the FST after a week of housing rats at moderate altitudes (Shami Kanekar et al., 2017, In Preparation). These data further support the hypothesis that housing of rats at altitude can negatively impact brain serotonin levels, to potentially increase DLB, anxiety, and anhedonia, and cause a loss of SSRI efficacy. An altitude-related loss in SSRI function may have important implications in the treatment of both anxiety disorders and depression in people at altitude, since SSRIs are highly prescribed both as antianxiety medications (Koen and Stein, 2011) and as antidepressants (Koenig and Thase, 2009). Future studies will examine the efficacy of SSRIs in treating anxiety associated with altitude.
This study has a few limitations. Rats were delivered from a sea level facility and were allowed to acclimatize at local conditions for a week at local conditions (4500 ft) before exposure to altitude conditions. A step-wise increment in altitude has been shown to mitigate the effects of altitude on physiology in humans (Beidleman et al., 2009), suggesting that the observed effects of altitude on anxiety and anhedonia may be stronger without a step-wise increment. Second, all experiments were conducted at local conditions of 4500 ft, which may also mitigate the effects of hypoxia. Third, this study is a snapshot of how exposure to altitude can alter rodent behavior. The impact of hypobaric hypoxia on brain chemistry and behavior has been shown to vary in impact depending on the intensity (e.g., 10,000 vs. 20,000 ft), duration (e.g., hours vs. weeks), and chronic versus intermittent nature of exposure (Perhonen et al., 1997; Sharma et al., 2002; Maiti et al., 2008). Fourth, we tested anxiety and anhedonic behaviors in SD rats during young adulthood (8–9 weeks of age), and previous studies indicate that hypoxia may exert differential effects on the brain based on age (Scremin et al., 1980; Gozal et al., 2001; Alchanatis et al., 2008). While the SD strain has been widely used in altitude studies (Ray et al., 2011; Baitharu et al., 2013), it is likely that other rat strains may show a different vulnerability or more resistance to the impact of altitude exposure. For instance, we hypothesize that the Wistar Kyoto rats, which are considered a model for refractory depression (Will et al., 2003) and anxiety vulnerability (McAuley et al., 2009), may be distinctly more vulnerable to exposure to hypobaric hypoxia than the SD rats used in this study. Studies are yet to define the impact of age and rat strain on brain and behavioral deficits induced by altitude exposure. Despite these limitations, these data do demonstrate that exposure to hypobaric hypoxia alone can have detrimental effects on rodent anxiety and anhedonia.
These data have important translational and clinical implications for a targeted approach to mental health in chronic hypoxic conditions. Our animal studies suggest that people living at altitude, and particularly women, may be at elevated risk for developing anxiety and mood disorders due to hypobaric hypoxia-induced alterations in the brain, and may be resistant to treatment with SSRIs, the most commonly prescribed antidepressants/antianxiety medications. These studies may also be significant for people exposed to chronic normobaric hypoxia by health conditions. People suffering from chronic hypoxic diseases such as asthma, chronic obstructive pulmonary disorder, chronic bronchitis, cardiovascular diseases, and sleep apnea are similar to those living at altitude in exhibiting higher rates of MDD, anxiety, and suicidal behavior (Goodwin, 2011, 2012; Pommer et al., 2012; Webb et al., 2012). Of utmost significance, a hypoxia-associated increase in depressive and anxiety disorders coupled with treatment resistance could increase suicidal ideation, to worsen suicidal behavior in people exposed to chronic hypoxia by living at altitude or with chronic hypoxic health conditions.
Conclusions
This study demonstrates that exposure to moderate altitude for a week can increase anxiety and anhedonia in female SD rats, but not in males. To the best of our knowledge, this is the first study to identify a detrimental effect of moderate-high altitude on anxiety and anhedonia in an animal model.
Footnotes
Acknowledgments
We extend special thanks to Mr. Barry Evans for his expertise with the altitude chambers and Dr. Kristen Keefe for use of the EPM apparatus. This study was supported by a Veterans Administration Merit Review grant to P.F.R. The views expressed in this report are those of the authors and do not necessarily represent those of the Veterans Affairs Health Systems.
Author Disclosure Statement
P.F.R. has received grants from the NIH, the VA, and USTAR, and compensation as a consultant to Kyowa Hakko Kirin and Ridge Diagnostics. The other authors have no disclosures to declare.