
Abstract
Abstract
Background/Aims:
High-altitude polycythemia is defined by the increase of hematocrit and hemoglobin at high altitudes caused by production of excessive erythrocytes. Eryptosis is a process by which mature erythrocytes undergo self-destruction sharing several features with apoptosis. However, the eryptosis in high-altitude hypobaric hypoxia is unknown. Thus, the main purpose of this study was to investigate whether chronic hypoxia affected eryptosis and, if so, by what mechanisms.
Methods:
Biotin labeling technology was utilized to study the survival of red blood cells in chronic hypoxia. Flow cytometry was used to determine the volume of mature erythrocytes from forward scatter, phosphatidylserine scrambling from annexin-V-binding, intracellular Ca2+ from Fluo-3-AM, reactive oxygen species (ROS) abundance from ROS-probe, and ceramide and CD47 abundance utilizing specific antibodies.
Results:
The volume of mature erythrocytes was significantly changed, and the percentage of annexin-V-binding cells was significantly decreased under chronic hypobaric hypoxia. Erythrocyte survival was improved under chronic hypoxia, and chronic hypoxia resulted in a decrease in intracellular Ca2+ in vivo and influenced eryptosis which was induced by the Ca2+-ionophore ionomycin (1 μM, 60 minutes) in vitro. Chronic hypoxia also resulted in an increase in CD47 and ceramide abundance, but it had no effect on ROS formation.
Conclusions:
Chronic hypobaric hypoxia can inhibit eryptosis by decreasing intracellular Ca2+ and increasing integrin-associated protein CD47.
Introduction
H
Thus, the purpose of this study was to investigate whether eryptosis was regulated by chronic hypoxia and, if so, by what mechanisms. To this end, flow cytometry was used to evaluate the volume of mature erythrocytes, phospholipid scrambling, intracellular Ca2+, ceramide abundance, integrin-associated protein CD47, and reactive oxygen species (ROS) in a chronic hypoxia rat model.
Materials and Methods
Chronic hypobaric hypoxia rat model
This experiment was approved by the Animal Research Committee of Qinghai University Medical College.
Twenty male Sprague-Dawley rats (Vital River, China) were randomly divided into two groups. Rats in the chronic hypoxic group were housed in a hypobaric chamber with a simulated altitude of 5000 m for 30 days (hypoxia group 5000 m), and those in the control group were maintained at an altitude of 2200 m in Xining, China. Blood samples were collected using an EDTA anticoagulant tube (BD) for RBC analysis.
RBC cultures and treatment with Ca2+ ionophore ionomycin
Fresh anticoagulated blood was centrifuged at 120 g for 20 minutes at 21°C, and the platelets and leukocyte-containing supernatant were disposed. RBCs were incubated in vitro at a hematocrit of 0.4% in Ringer solution which contained 125 mM NaCl, 5 mM KCl, 1 mM MgSO4, 32 mM HEPES, 5 mM glucose, 1 mM CaCl2; PH = 7.4. Eryptosis was triggered by Ca2+ ionophore ionomycin (1 μM,) (Merck Millipore, Germany) for 1 hour.
Biotin labeling and FACS analysis
In vivo RBC biotin labeling was performed as described previously (Gifford et al., 2006). Briefly, after housing 24 hours in 5000 m hypobaric chamber to adapt acute hypoxia, rats were anesthetized and an indwelling catheter was then placed proximal to the right atrium through a jugular vein. Blood was then labeled in vivo with NHS-biotin (30 mg/kg body weight) dissolved in dimethyl sulfoxide (DMSO) and injected. The catheter was then removed, and the vein was ligated. Biotin labeled cells were analyzed using a APC-Streptavidin antibody; 1 × 106 RBCs were stained with APC-Streptavidin antibody (BioLegend) for 1 hour at 4°C under protection from light and washed twice with phosphate-buffered saline (PBS)-bovine serum albumin (BSA).
FACS analysis of phospholipid scrambling and forward scatter
Phospholipid scrambling was analyzed using a FITC-Annexin V Apoptosis Detection Kit (BD) according to the manufacturer's instructions, and 1 × 106 RBCs were stained with Annexin-V-FITC at 37°C for 20 minutes under protection from light. The forward scatter (FSC) and Annexin-V fluorescence intensity were measured at an excitation wavelength of 488 nm and an emission wavelength of 530 nm on a FACSCalibur (Beckman, Germany).
Measurement of intracellular Ca2+
Intracellular Ca2+ was measured as described previously (Lupescu et al., 2014). In short, 1 × 106 RBCs were washed in Ringer solution and then loaded with 2 μM Fluo-3/AM (Biotium, Hayward, CA). After that, they were incubated at 37°C for 30 minutes under protection from light and washed twice. Ca2+-dependent fluorescence intensity was measured at an excitation wavelength of 488 nm and an emission wavelength of 530 nm.
Determination of ceramide formation and CD47
For determination of ceramide formation, 1 × 106 RBCs were stained for 1 hour at 4°C with ceramide monoclonal antibody (MID 15B4; Enzo Life Sciences) in PBS containing 0.1% BSA at a dilution of 1:5. After incubation with the first antibody, cells were washed twice with PBS-BSA and then stained for 30 minutes with the secondary antibody FITC-conjugated goat anti-mouse IgM (Southern Biotech) at a dilution of 1:100. Unbound secondary antibody was removed by repeated washing with PBS-BSA, and then cells were washed twice with PBS-BSA and analyzed. For CD47 analysis, 1 × 106 RBCs were stained with FITC-CD47 antibody (BioLegend) for 1 hour at 4°C under protection from light and washed twice with PBS-BSA.
Measurement of ROS in RBCs
The ROS in RBCs was determined using a total ROS/Superoxide Detection Kit (ENZ-51010; Enzo Life Sciences) according to the manufacturer's instructions. In brief, 1 × 106 RBCs were stained with Oxidative Stress Detection Reagent (Green) and Superoxide Detection Reagent (Orange) from light, and fluorescence intensity was measured with FL1 for Green (490/525 nm) and FL2 for Orange (550/620 nm), respectively.
Statistical analyses
All statistical analyses were performed with the GraphPad Prism 6 software. Data are expressed as mean ± standard error of mean (SEM). Statistical significance was tested using t-test. A value p < 0.05 was considered statistically significant.
Results
As shown in Table 1, the RBC, haemoglobin, and hematocrit levels were significantly increased, and the number of platelet was significantly decreased in chronic hypoxia group. Erythrocyte FSC was used to test whether chronic hypoxia had an effect on the volume of RBCs. The exposure to the simulated altitude of 5500 m for 30 days resulted in significant changes in the volume of RBCs. Specifically, the number of normal sized RBCs (10 < FSC <40) was reduced, but that of large RBCs (FSC ≥40) was increased (Fig. 1A). As shown in Figure 1B, the percentage of large RBCs was increased significantly in hypoxia group (1.637 ± 0.1649, n = 10 vs. 1.128 ± 0.0644, n = 10; p = 0.0101), while that of small RBCs was not changed (0.0890 ± 0.0083, n = 10 vs. 0.0710 ± 0.0088, n = 10; p = 0.1542).
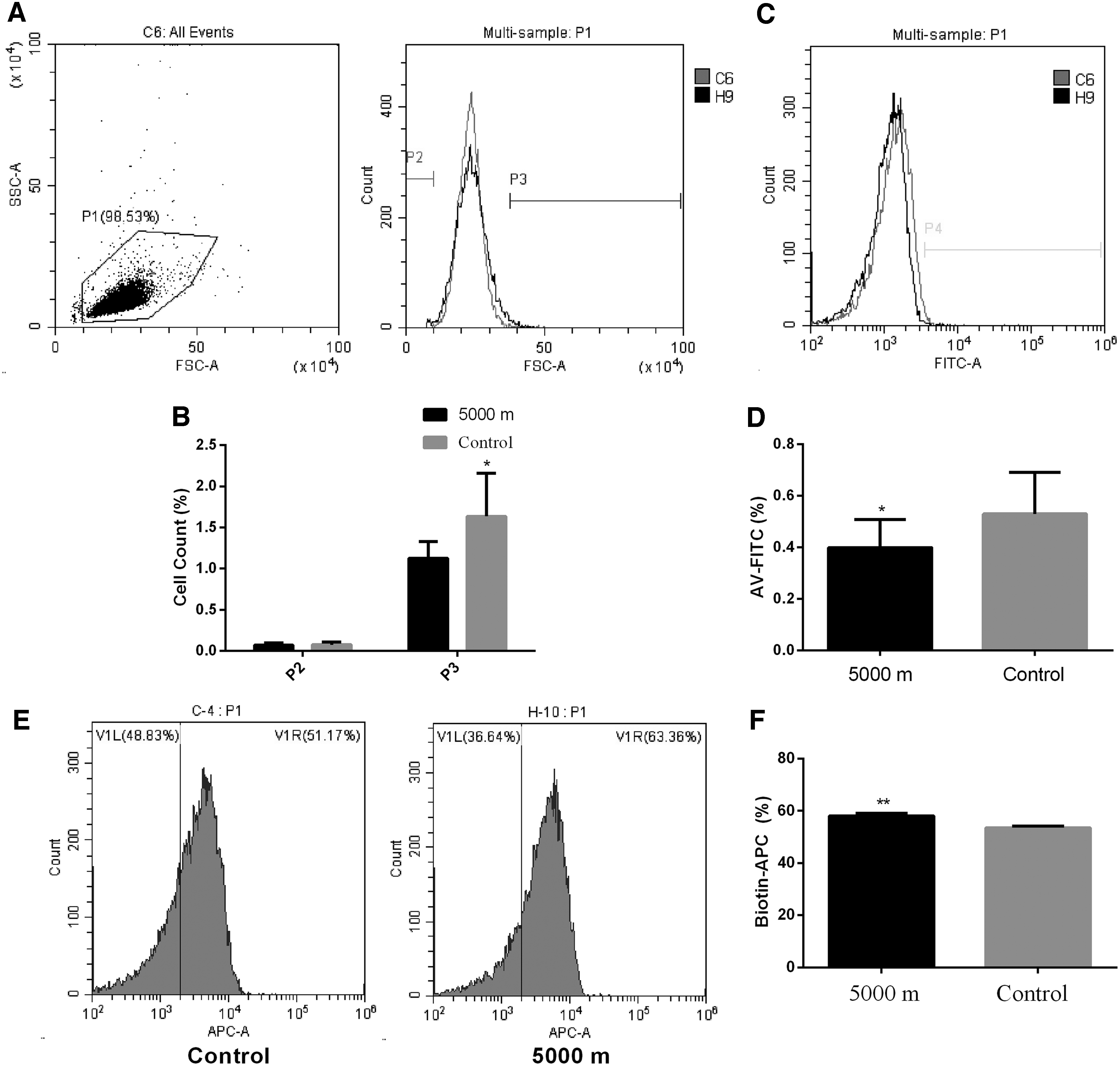
Effect of chronic hypobaric hypoxia on FSC and phosphatidylserine.
Blood Cell Count Analysis
The results were expressed as mean ± SEM.
Hb, hemoglobin; HCT, hematocrit; PLT, platelet; RBC, red blood cell; WBC, white blood cell.
The most important marker for eryptosis is phospholipid scrambling. The effect of chronic hypoxia on phospholipid scrambling was assessed by the percentage of rat erythrocytes binding annexin-V after exposure to hypoxia for 30 days. As shown in Figure 1C, the curved line moved to the left, indicating that the overall fluorescence intensity was reduced in hypoxia group. The percentage of erythrocytes binding annexin-V was significantly decreased in hypoxia group (0.3990 ± 0.0346, n = 10 for hypoxia group vs. 0.5300 ± 0.0511, n = 10 for control group, p = 0.0478). Biotin labeling was used to study RBC survival in chronic hypoxia. We found that biotin-positive RBCs were significantly increased in chronic hypoxia group (58.18 ± 0.8716 vs. 53.56 ± 0.5938, n = 10, p = 0.0004) (Fig. 1E, 1F). These data indicated that chronic hypoxia inhibited eryptosis.
The increase of cytosolic Ca2+ could induce cell membrane scrambling. To identify chronic hypoxia regulated cytosolic Ca2+ in RBCs, Fluo3 AM was used to quantify cytosolic Ca2+. As shown in Figure 2A and B, Fluo3 fluorescence was significantly decreased in hypoxia group compared with that in control group (3721 ± 105.0, n = 10 vs. 4143 ± 53.74, n = 10, p = 0.0038), indicating that cytosolic Ca2+ of RBCs was decreased in chronic hypoxia group. The percentage of erythrocyte Annexin V (AV)-FITC positive rate was significantly decreased in hypoxia group when exposed to Ca2+-ionophore ionomycin for 60 minutes (59.83 ± 3.54 vs. 81.67 ± 6.76, p < 0.0001) (Fig. 2C, D).
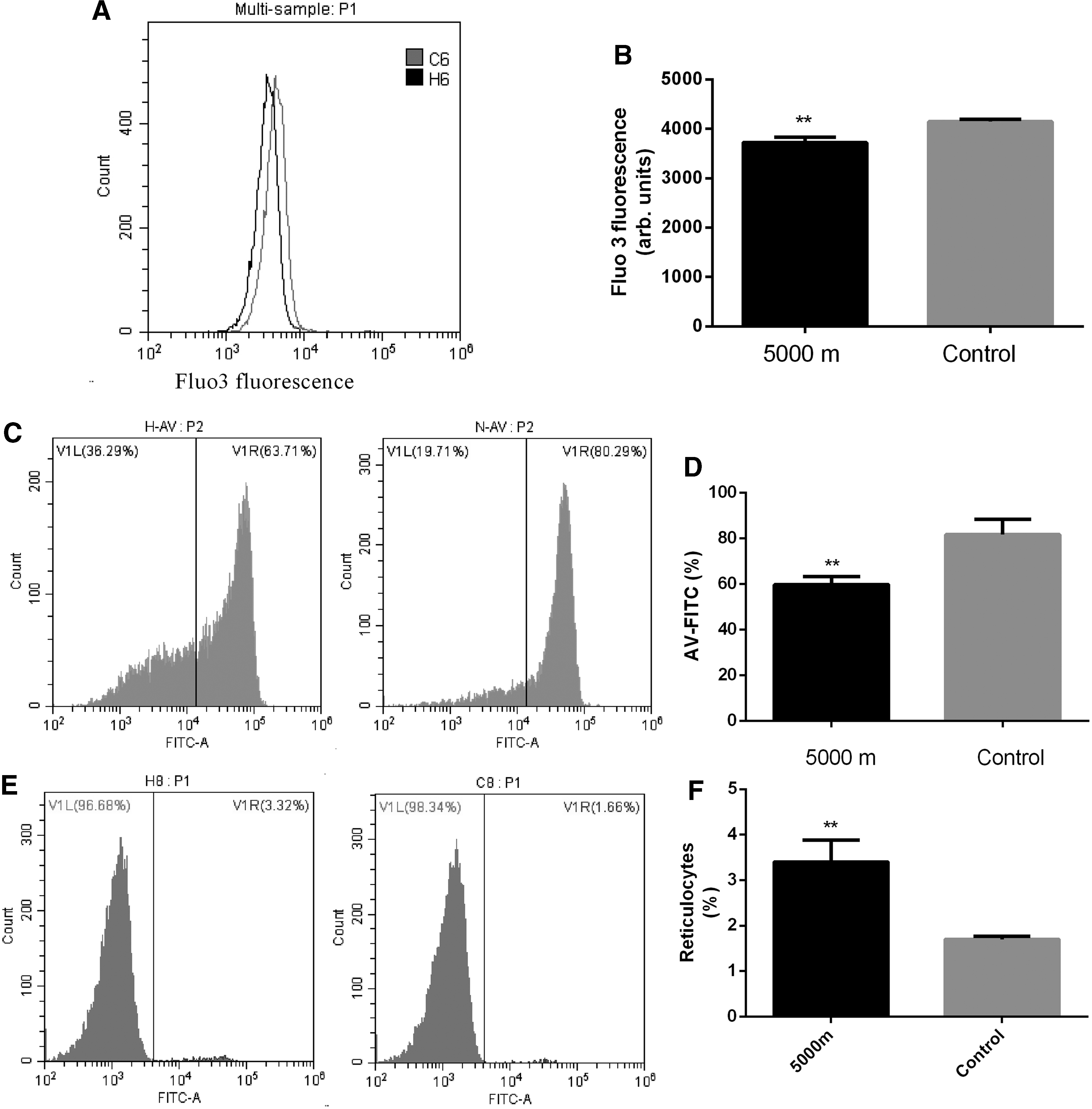
Effect of chronic hypobaric hypoxia on intracellular Ca2+.
The effect of chronic hypoxia on reticulocytes was determined by Flow cytometry. As shown in Figure 2E and F, the number of reticulocytes was significantly increased in hypoxia group (3.404 ± 0.4819, n = 10 vs. 1.701 ± 0.0659, n = 10, p = 0.0025).
Ceramide could enhance Ca2+ sensitivity of cell membrane scrambling (Mischitelli et al., 2016). Ceramide abundance at rat RBC surface was quantified using specific antibodies. As shown in Figure 3, the ceramide abundance was significantly increased in hypoxia group (2288 ± 187.1, n = 10 vs. 1684 ± 22.17, n = 10, p = 0.0049).

Effect of chronic hypobaric hypoxia on ceramide.
We assessed the role of oxidative stress by measuring the percentage of erythrocytes with ROS. However, there was no significant difference in the percentage of erythrocytes between hypoxia and control groups (1565 ± 123.0 vs. 1225 ± 148.0, respectively, p = 0.0945), as shown in Figure 4.

Effect of chronic hypobaric hypoxia on erythrocyte ROS.
CD47 in conjunction with signal regulatory protein alpha could prevent the clearance of RBCs by macrophages (Banerjee et al., 2015). To examine whether the effect of chronic hypoxia involved RBC phagocytized by macrophages, the effect of chronic hypoxia on CD47 was tested. As shown in Figure 5, CD47+ RBC was significantly increased in hypoxia group (4.997 ± 0.8167, n = 10 vs. 2.746 ± 0.3264, n = 10, p = 0.0197).
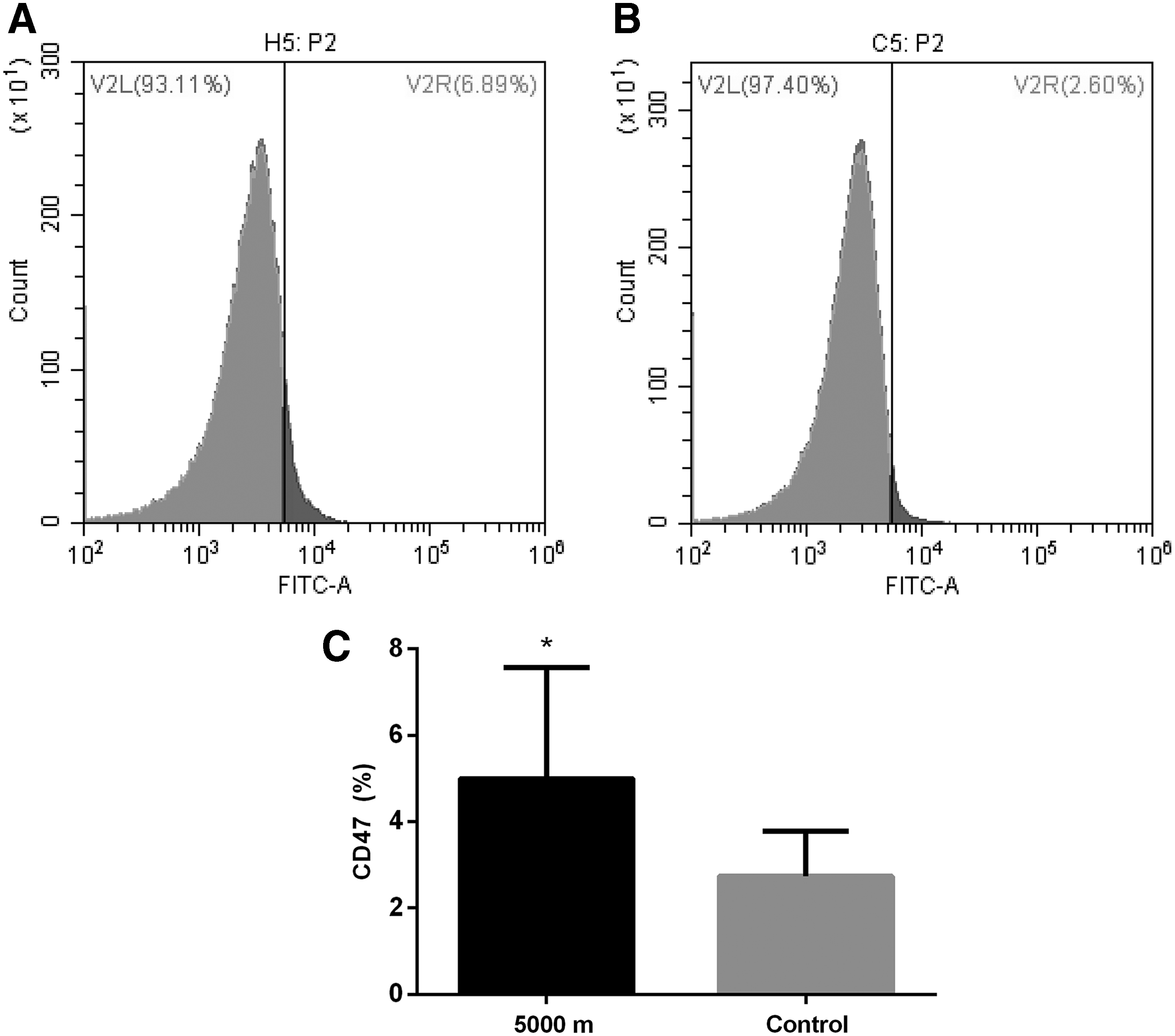
Effect of chronic hypobaric hypoxia on CD47.
Discussion
Previous studies on the mechanisms of HAPC have focused on the regulation of hematopoietic stem progenitor cells. However, the effect of chronic hypoxia on mature RBCs remains largely unknown. The present study revealed that hypoxia could not only cause an increase in erythrocyte formation but also it could affect the volume (Fig. 1A) and eryptosis (Fig. 1C) of mature RBCs. We also found that erythrocyte survival was improved under chronic hypoxia by biotin labeling technology (Fig. 1E). Interestingly, eryptosis was decreased by decreasing intracellular Ca2+ both in vivo and in vitro (Fig. 2) and cell membrane CD47 (Fig. 5) rather than by regulating ceramide (Fig. 3) or ROS (Fig. 4) in the chronic hypoxic rat model.
Unlike nucleated cells, there is no apoptosis in RBCs. Aged RBCs are normally phagocytized by macrophages of splenic and hepatic sinusoids in a process known as erythrophagocytosis (Wynn et al., 2013). Cell membrane scrambling and erythrocyte shrinkage are the hallmark of eryptosis, the suicidal erythrocyte death. Our results showed that FSC moved to the right (Fig. 1A), and AV-binding cells were significantly decreased (Fig. 1C) in rats exposed to a simulated altitude of 5000 m. CD47, the integrin-associated protein, acts as an autologous marker on erythrocytes (Basu et al., 2010). We found that CD47 was increased in 5000 m group (Fig. 5). RBC destruction is ultimately controlled by the antagonist effects of phosphatidylserine and CD47 (Arias, 2017). Erythrocyte cell membrane scrambling with phosphatidylserine makes erythrocytes easily to be phagocytosed by macrophages, and the presence of CD47 on the surface of erythrocyte diminishes phagocytosis (Burger et al., 2012). In the present study, we found that the levels of phosphatidylserine scrambling were decreased, and the CD47 expression was increased in chronic hypoxia group. The decrease level of cell membrane scrambling and increase level of CD47 could improve the resistance of erythrocytes to damage.
Suzuku and Dale (1987) first reported that the biotin labeling technology in studying RBC aging in 1987 labeled in vitro with NHS-biotin and retransfused into experiment animal exhibited normal aging dynamics. Dale and Norenberg (1990) found that this technique could be extended to in vivo labeling, and 90% of all circulating erythrocytes were labeled by intravenously infused biotin. Gifford et al. (2006) injected biotin into right atrium and found that it could raise the labeling rate to 99.8% in rats. We used this technology to study the survival of RBCs under chronic hypoxia, and we found that the percentage of biotin positive RBCs was higher in chronic hypoxia group (Fig. 1). It indicated that erythrocyte survival was improved in chronic hypoxia.
Exposure to high-altitude hypoxia can result in an increase in RBCs to compensate for hypoxia (Leon-Velarde et al., 2005). In this study, we also observed that hypoxia stimulated RBC production (Table 1 and Fig. 2E, F). However, an increased number of reticulocytes might lead to a decrease in AV-binding cytosolic Ca2+ and an increase in CD47. As expected, there was an overall decline of Annexin-V and cytosolic Ca2+ fluorescence intensity, especially in Ca2+ fluorescence (Fig. 2A, B). The same results were observed after in vitro culture in presence of ionomycin (Fig. 2C, D). These results indicated that hypoxia improved the resistance to cytosolic Ca2+ induced eryptosis. The effect of chronic hypoxia on reducing cell membrane scrambling was paralleled by a decrease of cytosolic Ca2+. An increase of cytosolic Ca2+ could stimulate phosphatidylserine scrambling and induce eryptosis (Lang et al., 2015). The concentration of cytosolic Ca2+ is regulated by many factors such as Gardos K(+) channels (Foller et al., 2010), platelet activating factor (PAF) (Xu et al., 2009), prostaglandin (Muravyov and Tikhomirova, 2014), and protein kinase C (Lang and Lang, 2015; Willems et al., 2015). Our results showed that the decreased intracellular Ca2+ prevented phosphatidylserine scrambling in hypoxia.
In this study, excess erythropoiesis was observed in chronic hypoxia rats (Table 1 and Fig. 2) and hypoxia inhibited eryptosis (Fig. 1), indicating that the imbalance between erythropoiesis and erythrocyte destruction may have contributed significantly to polycythemia. Hypoxia induces secretion of erythropoietin (Epo) (Arias, 2017) and proliferation and differentiation of erythroid precursor cells to form erythrocytes, which can be regulated by hypoxia inducible factors (Rankin et al., 2007). EPO can directly inhibit eryptosis through downregulated cation channel activity and decreased intracellular Ca2+ (Lang et al., 2016).
Eryptosis can also be triggered by enhancing ceramide abundance (Lang et al., 2015). Ceramide is generated by the breakdown of cell membrane sphingomyelin and sensitizes erythrocytes to the enhanced intracellular calcium concentration. PAF is overexpressed and involved in the development of multiple hypoxic injuries (Caplan et al., 1991; Palmer, 1995; Logani et al., 2000). Ceramide is regulated by PAF which stimulates sphingomyelinases (Lang and Lang et al., 2010; Jilani et al., 2013). The increase of ceramide abundance is always accompanied by the increase in membrane phosphatidylserine scrambling. Interestingly, the results of this study showed an increase in ceramide abundance in chronic hypoxia group (Fig. 3). ROS showed no significant difference (Fig. 4) in chronic rat model. These results indicated that chronic hypoxia regulated eryptosis not through PAF/ceramide pathway and oxidative stress, and some unknown mechanisms could be involved in chronic hypoxic induced eryptosis inhibition.
Conclusions
Chronic hypoxia inhibited eryptosis due to decreased intracellular Ca2+ and increased integrin-associated protein CD47, which supports the notion of its significant contribution to polycythemia at high altitude.
Footnotes
Acknowledgments
This work was funded by National Natural Science Foundation of China (No. 81360300, FT; No. 32571231, RLG), The Provence Natural Science Foundation of Qinghai (No. 2013-Z-931Q), and 123 Talent Project of Qinghai University. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the article.
Author Disclosure Statement
No competing financial interests exist.