
Abstract
Abstract
Cabrera-Aguilera, Ignacio, David Rizo-Roca, Elisa A. Marques, Garoa Santocildes, Teresa Pagès, Gines Viscor, António A. Ascensão, José Magalhães, and Joan Ramon Torrella. Additive effects of intermittent hypobaric hypoxia and endurance training on bodyweight, food intake, and oxygen consumption in rats. High Alt Med Biol. 19:278–285, 2018.—We used an animal model to elucidate the effects of an intermittent hypobaric hypoxia (IHH) and endurance exercise training (EET) protocol on bodyweight (BW), food and water intake, and oxygen consumption. Twenty-eight young adult male rats were divided into four groups: normoxic sedentary (NS), normoxic exercised (NE), hypoxic sedentary (HS), and hypoxic exercised (HE). Normoxic groups were maintained at an atmospheric pressure equivalent to sea level, whereas the IHH protocol consisted of 5 hours per day for 33 days at a simulated altitude of 6000 m. Exercised groups ran in normobaric conditions on a treadmill for 1 hour/day for 5 weeks at a speed of 25 m/min. At the end of the protocol, both hypoxic groups showed significant decreases in BW from the ninth day of exposure, reaching final 10% (HS) to 14.5% (HE) differences when compared with NS. NE rats also showed a significant weight reduction after the 19th day, with a decrease of 7.4%. The BW of hypoxic animals was related to significant hypophagia elicited by IHH exposure (from 8% to 12%). In contrast, EET had no effect on food ingestion. Total water intake was not affected by hypoxia but was significantly increased by exercise. An analysis of oxygen consumption at rest (mL O2/[kg·min]) revealed two findings: a significant decrease in both hypoxic groups after the protocol (HS, 21.7 ± 0.70 vs. 19.1 ± 0.78 and HE, 22.8 ± 0.80 vs. 17.1 ± 0.90) and a significant difference at the end of the protocol between NE (21.3 ± 0.77) and HE (17.1 ± 0.90). These results demonstrate that IHH and EET had an additive effect on BW loss, providing evidence that rats underwent a metabolic adaptation through a reduction in oxygen consumption measured under normoxic conditions. These data suggest that the combination of IHH and EET could serve as an alternative treatment for the management of overweight and obesity.
Introduction
O
Nonpharmacological overweight treatments aim to reverse the energy balance from positive to negative by reducing energy intake and increasing energy expenditure, with diet restriction and an increase in physical activity being the first recommended steps (Vairavamurthy et al., 2017). Many guidelines indicate that daily sessions of moderate exercise contribute to treating obesity (Jeffery et al., 2003), and it has been proven that endurance exercise training (EET) is especially useful in reducing fat mass while maintaining or gaining lean mass (Hansen et al., 2009).
It has also been described that chronic hypobaric hypoxia and time spent at high altitude elicit appetite suppression and a decrease in food intake, resulting in an energy imbalance that leads to weight loss (Surks et al., 1966; Boyer and Blume, 1984; Rose et al., 1988; Westerterp et al., 1994), although intermittent exposure to hypobaric hypoxia does not exert a detrimental effect on body mass in laboratory rats, at least under certain experimental conditions (Panisello et al., 2007). Traditionally, it has been accepted that chronic exposure to hypoxia has deleterious effects on health that range from pulmonary hypertension to loss of muscle mass, the latter contributing to weight loss through body deterioration (Pierson, 2000).
These adverse effects can be partially prevented by intermittent hypoxic exposure, and it has been suggested that adequate “doses” of hypoxia (i.e., intensity, frequency, and duration) may trigger compensatory adaptive mechanisms beneficial for health (such as BW loss), while minimizing its negative side effects (Chen et al., 2005; Dematteis et al., 2008; Zoccal et al., 2008). Although the cellular mechanisms and pathways remain unclear and the data are contradictory, there is evidence that intermittent hypoxia reduces blood glucose levels (Brooks et al., 1991), improves insulin sensitivity (Azevedo et al., 1995), and increases muscle glucose transporter-4 (GLUT-4) expression (Chiu et al., 2004). Hypoxia also stimulates the adrenergic system (Antezana et al., 1994) and promotes leptin gene expression (Ambrosini et al., 2002), thus increasing leptin release (Shukla et al., 2005). These events could directly or indirectly modulate food intake, energy expenditure, appetite control, and fat loss.
Given the abovementioned effects of endurance exercise and hypoxia exposure on BW, our main question was whether both strategies could be combined to induce an additive effect on BW. We also aimed to ascertain whether intermittent hypobaric hypoxia (IHH) and EET promote additive effects on food and water intake and oxygen consumption.
Materials and Methods
Animals
Twenty-eight Wistar male rats weighing 272 ± 5.9 g (mean ± SEM) were randomly divided into four groups (n = 7 each): normoxic sedentary (NS), normoxic exercised (NE), hypoxic sedentary (HS), and hypoxic exercised (HE). All animals were maintained in a room at T = 22°C and 50%–60% relative humidity in 12-h light/12-h dark cycles. Rats had unrestricted access to food (Scientific Animal Food and Engineering, A04) and untreated tap water. The study was approved by the local Institutional Review Board and followed the guidelines for care and use of laboratory animals in research recommended by the Federation of European Laboratory Animal Science Associations (FELASA).
IHH and EET protocols
Hypoxic groups (HS and HE) were subjected to an IHH protocol in a hypobaric chamber made of polymethylmethacrylate plastic walls, where vacuum was created by a pump (TRIVAC D5E; Leybold, Köln, Germany) controlled by two differential sensors (ID 2000; Leybold) driving a diaphragm pressure regulator (MR16; Leybold). The entire IHH protocol lasted for 33 days, with an initial 7 days of progressive acclimatization. On the first day, rats were exposed for 5 hours to a simulated altitude of 2500 m. Thereafter, altitude was incremented by 500 m/day until reaching a simulated altitude of 6000 m on the eighth day. From the eighth day until the 33rd day, animals were exposed for 5 hours/day and 5 days/week to a simulated atmospheric pressure of 49.3 kPa (369.8 mm Hg), equivalent to an altitude of 6000 m above sea level. The depressurization periods to reach the altitude and return to sea level conditions lasted for 12 minutes each.
After the hypoxic period (from 7.00 to 12.00 hours), HE animals were allowed to rest for 4 hours before initiating the endurance treadmill exercise. Normoxic groups (NS and NE) were maintained at a barometric pressure of 101.3 kPa (760 mm Hg), equivalent to sea level. Exercised groups (NE and HE) were adapted to the treadmill (LE 8710; Panlab, Barcelona, Spain) for 7 days before beginning the training program. For the first two days of exercise, animals were trained at 15 m/min and then the speed was increased by 5 m/min until reaching 25 m/min. The endurance exercise consisted of 1 hour/day of treadmill running for 5 weeks at a speed of 25 m/min (0% gradient). The nonexercised groups (NS and HS) were placed on a nonmoving treadmill for 1 hour to expose the animals to potential handling and environment stresses similar to those induced by the treadmill.
Body, heart, and gastrocnemius weight measurements and food and water intake
Every 3–4 days, animals were weighed and water and food intake was recorded. Total water and food intake was calculated at the end of the experiment and expressed as g/(kg·day) for each group (mean ± SEM). After 5 weeks of hypoxic and exercise training, animals were anesthetized with 50 mg/kg of sodium pentobarbital, their hearts and right gastrocnemius were excised and weighed, and heart-to-bodyweight (HW/BW) and right gastrocnemius-to-bodyweight (GW/BW) ratios were calculated.
Measurement of oxygen consumption
All oxygen consumption (
The
where
Statistics
Data for all parameters are expressed as sample mean ± standard errors. The Kolmogorov–Smirnov test (with Lilliefors correction) was used to test the data for normality. Homoscedasticity was determined by Levene's test. Comparisons between the four groups of BW evolution, food and water intake, and heart and gastrocnemius weights (GWs) were analyzed using a two-way analysis of variance (ANOVA). IHH and EET were considered as factors with two levels for each factor: normoxia or hypoxia (for IHH) and sedentary or exercised (for EET). The pairwise differences between the resulting groups (NS, NE, HS, and HE) were tested by means of a post hoc multiple comparison test (Holm–Sidak method). All ANOVA statistical tests were run using SigmaPlot 11 software (Systat Software, Inc., San Jose, CA). The level of statistical significance (p < 0.05, p < 0.01, or p < 0.001) is indicated in the figures, tables, and text.
To test differences in oxygen consumption before and after the protocols, an analysis of covariance (two-way analysis of covariance [ANCOVA]) with repeated measures was used. The ANCOVA considered two times (initial and final) × four groups (exercise and control) and was adjusted for percent weight change to determine differences between and within groups for dependent variables. When the ANCOVA revealed a significant interaction (time × group), the Bonferroni post hoc test was performed to determine differences between initial and final values in each group. ANCOVA statistical analysis was performed using PASW Statistics 18 for Windows with a significance level of p < 0.05.
Results
BW, HW, and right GW
Figure 1 shows the evolution of the rats' BWs for the different experimental groups. The first significant difference (p < 0.05) appeared between NS and HE after 9 days of IHH and EET. This difference increased as the IHH and EET protocol progressed, until reaching a maximum significant difference (p < 0.001) between these two groups at the end of the study, on the 33rd day, with a final BW difference of 14.5% in HE rats when compared with control NS animals (Table 1). The first significant effect (p < 0.05) of IHH alone appeared in sedentary animals on the 12th day of hypoxic exposure, reaching a final difference of almost 10% (p < 0.001) between NS and HS groups (Table 1). The significant (p < 0.05) effect of EET on BW, independently of IHH exposure, was first observed after 19 days of exercise protocol (Fig. 1), with a 7.4% BW difference at the end of the protocol (Table 1). The effect of IHH on exercised rats (p < 0.05) was observed after the 23rd day of hypoxic exposure, with a final 7.7% significant difference (p < 0.05) in BW between HE and NE animals (Table 1). No statistically significant differences in BW were detected between NE and HS groups (Fig. 1, Table 1).
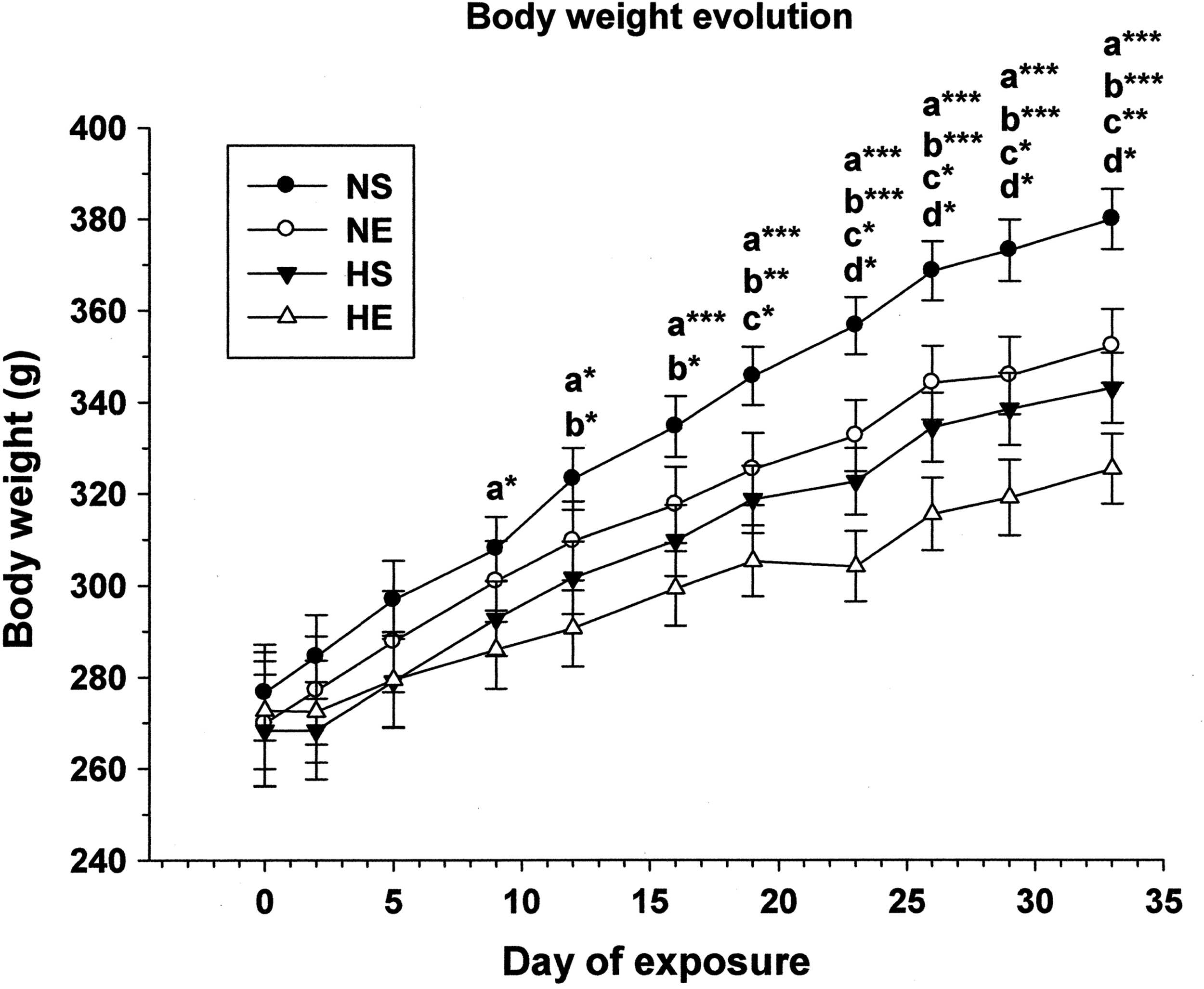
Bodyweight evolution in rats during combined exercise and intermittent hypobaric hypoxia exposure protocols. Asterisks represent significant differences at the levels *p < 0.05, **p < 0.01, and ***p < 0.001 between the following pairs: a, NS versus HE; b, NS versus HS; c, NS versus NE; d, NE versus HE. NS, normoxic sedentary; NE, normoxic exercised; HS, hypoxic sedentary; HE, hypoxic exercised.
Final BW, HW, and right GW in rats after applying a combined exercise and intermittent hypobaric hypoxia protocol. Ratios of heart weights (HW/BW) and right gastrocnemius weights (GW/BW) relative to bodyweights are expressed as milligrams of organ per gram of animal (mg/g).
Significant differences versus NS.
Significant differences versus NE.
Levels of significance: *p<0.05, **p<0.01, ***p<0.001.
BW, bodyweight; GW, gastrocnemius weights; HW, heart weights; NS, normoxic sedentary; NE, normoxic exercised; HS, hypoxic sedentary; HE, hypoxic exercised.
Heart weight (HW) was very similar in all groups (p > 0.05), with little and similar dispersion of all means. However, due to the abovementioned differences in final BW, the HW/BW ratio was significantly higher in both hypoxic groups (sedentary and exercised) compared with their normoxic counterparts (Table 1). Differences were observed in right GW, whereby exercised groups (NE and HE) displayed significantly higher GW than NS animals. HS rats also showed significantly higher GW than their normoxic counterparts, while presenting a wide dispersion in measures (Table 1). The combination of GW differences and significant differences in the final BW resulted in higher significance values for the GW/BW ratio (p < 0.001) in all groups compared with NS. Moreover, the final BW reduction observed in both hypoxic groups (HS and HE) yielded significantly higher values for this ratio when compared with NE animals.
Food and water intake
Figure 2 shows mean food and water intake in grams per kilogram of BW and day in all groups of animals. As can be seen in the food histograms, IHH exposure induced hypophagia. Both hypoxic groups (HS and HE) presented a significantly lower food intake than the normoxic animals (NS and NE) when comparing all the possible pairs. This reduction in food intake ranged from 8% to 12%. In contrast, the exercise protocol had no effect on food ingestion since similar values (p > 0.05) were found when comparing NS versus NE and HS versus HE groups.
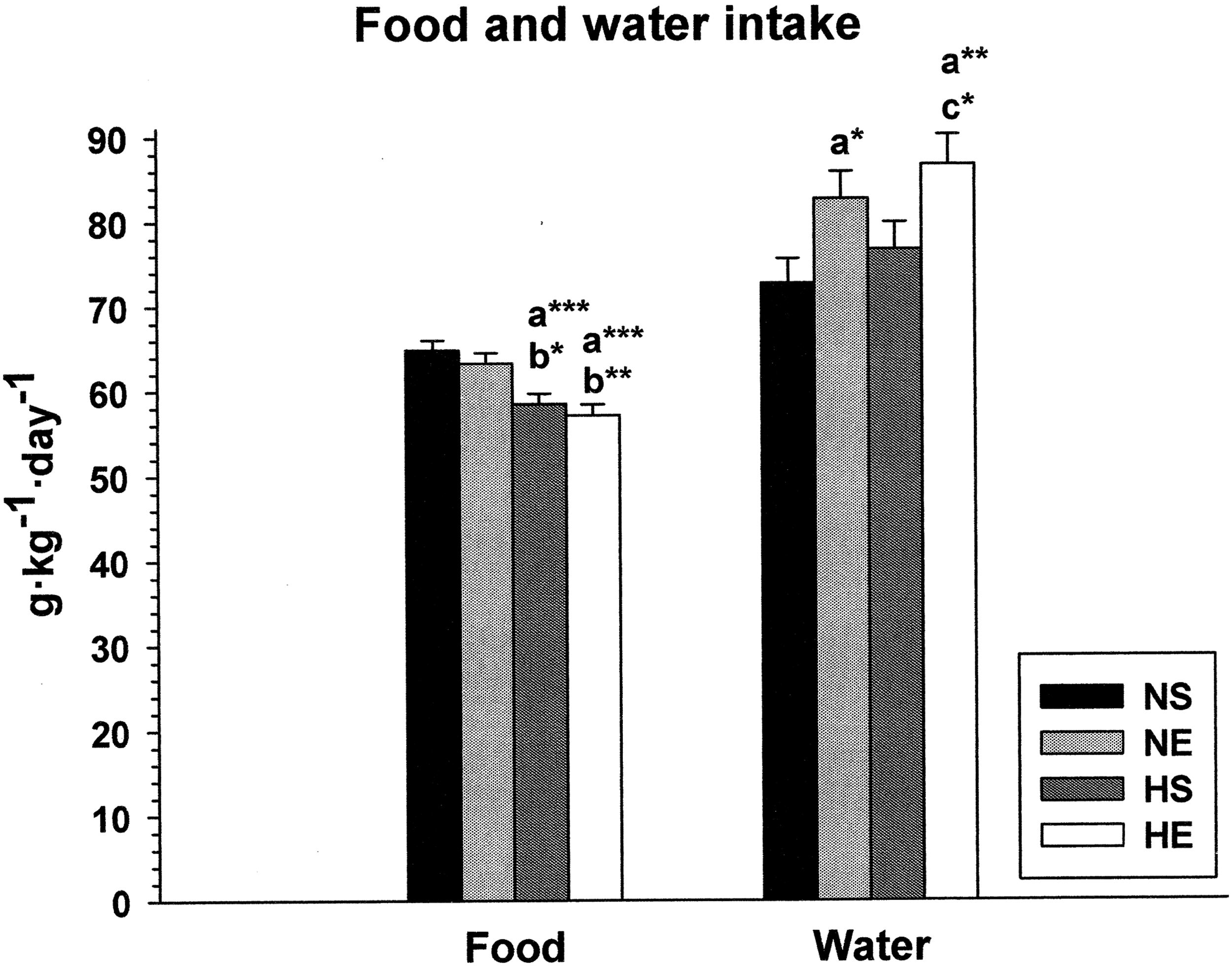
Food and water intake in rats during a combined exercise and intermittent hypobaric hypoxia protocol. Asterisks represent significant differences at the levels *p < 0.05, **p < 0.01, and ***p < 0.001 between the following pairs: a, versus NS; b, versus NE; c, versus HS.
Water intake was unaffected by the IHH exposure protocol since the differences between NS versus HS and NE versus HE were not statistically significant. However, clear statistically significant differences were observed between sedentary and exercised animals whereby the latter drank more water than the former. This increment in water intake ranged from 13% to 20%.
Oxygen consumption
Table 2 shows the mean oxygen consumption for each group before the beginning of the experimental procedures and at the end of the study. As explained in the Materials and Methods section, all oxygen consumption measurements were performed under normoxic conditions and the statistical test used to compare groups (ANCOVA) ruled out the effects of weight gain on oxygen consumption. Two interesting findings are of note. First, there was a significant reduction (p < 0.05) in oxygen consumption in both hypoxic groups (HS and HE) after IHH exposure, that is, we found a within-group difference between before and after IHH. Second, there was a statistically significant between-group difference (p < 0.05) after the IHH and EET protocol between NE and HE, suggesting an additive effect of hypoxia and exercise on the reduction in oxygen consumption, since no effect was observed when comparing NS with HS animals.
Oxygen consumption in rats before and after the experimental procedures. See Statistics for details.
p<0.05 NE versus HE.
p<0.05 within-group difference (Before versus After).
Discussion
BW and GW
We found a stepwise sequence of BW changes in the different experimental groups, with an additive effect of IHH and EET on reducing BW, which was significant from the ninth day. Moreover, IHH may have provided a stronger stimulus for BW loss than EET, as differences between NS and HS appeared 1 week before the differences between NS and NE (Fig. 1). These results shed interesting light on the possibility of losing weight under hypoxia, as has already been suggested elsewhere (Quintero et al., 2010; Urdampilleta et al., 2012; Kayser and Verges, 2013).
Several factors should be considered regarding hypoxic conditions, including the exposure pattern (intermittent, acute, or chronic) and type of hypoxia (normobaric or hypobaric), the intensity of hypoxia (the inspired PO2), and the dose of hypoxia (hours of exposure). For example, in a similar study, Chiu et al. (2004) also described an additive effect of IHH and EET and very similar BW evolution in the four groups (control, endurance training, hypoxic, and hypoxic followed by endurance training); however, they reported a greater effect of EET than IHH exposure on BW loss. The differences with our results could be attributed to the kind of hypoxia (normobaric vs. hypobaric), the level of hypoxia (14% vs. 9.5% inspired O2), and the dose of hypoxia (12 hours/day vs. 5 hours/day). Moreover, the exercise protocol was also different and would have had some influence on the differences, since their animals swam instead of ran and for a longer period of time (2–3 hours/day vs. 1 hours/day).
There is controversy about which body mass component is most affected by hypoxia and therefore responsible for the associated weight loss. It is well known that weight loss during prolonged exposure to hypoxia is mostly due to loss of muscle mass and dehydration caused by water imbalance (Rose et al., 1988; Frappell et al., 1992; Lippl et al., 2010). However, other studies have suggested that fat mass is the main component of body mass loss (Guillan and Klepping, 1985; Brooks et al., 1991). Nevertheless, our results clearly demonstrate that the EET protocol significantly increased absolute GW both in normoxic and hypoxic animals, which is explained by the well-known effects of endurance exercise on muscle hypertrophy (Konopka and Harber, 2014). Combined with the reduction in BW, this resulted in significantly higher GW/BW ratios. Surprisingly, the IHH protocol by itself also increased GW and GW/BW. Additional studies are needed to better explain this finding, especially in view of the high dispersion of GW values in this group.
Food and water intake
We observed a significant 8%–12% reduction in food intake in the hypoxic groups (Fig. 2). This is in agreement with several studies that have reported a hypophagic response under hypobaric hypoxia, both in humans and in rats (Guilland and Klepping, 1985; Rose et al., 1988). In some cases, this reduction has ranged from 10% to 50% (Sharma et al., 2002), and the intensity of hypophagia is probably related to the time and kind of hypoxia exposure. This hypoxia-induced hypophagia has been associated with the possible regulation of anorexigenic factors, such as leptin and ghrelin (Ambrosini et al., 2002; Shukla et al., 2005; Morishima and Goto, 2016), and may be a critical factor in weight loss associated with hypoxia.
In any case, since our results show that exercised animals had a similar food intake than sedentary animals, and considering the additional energy expenditure provoked by exercise, a more negative energy balance in exercised animals would explain their lower final BW when compared with their sedentary counterparts.
The results shown in Figure 2 also indicate a tendency, although not significant, toward additional water intake with the hypobaric hypoxia protocol when comparing NS versus HS and NE versus HE. Although pioneering studies of hypoxia in humans related weight loss during periods at high altitude to water loss (Gill and Pugh, 1964; Surks et al., 1966; Consolazio et al., 1968; Krzywicki et al., 1969), the fluid balance data obtained during Operation Everest II in a hypobaric chamber suggested that subjects were probably not dehydrated (Rose et al., 1988). The intermittent nature of our protocol may have elicited adequate acclimatization to altitude without excessive fluid loss, whereas the small, nonsignificant differences recorded might be due to the well-known hyperventilation described as a consequence of altitude exposure (Prabhakar, 2013), given that part of the water is lost during respiration through pulmonary ventilation.
The EET protocol alone did not seem to alter food intake, since no significant differences were found between sedentary and exercised animals, either under normoxic or hypoxic conditions. Indeed, the effects of exercise on food intake are still controversial, but a classic review (Blundell and King, 1999) showed that only 19% of all analyzed studies found an increase in energy intake after exercise, whereas 65% reported no changes regardless of the type and intensity of the exercise. Although exercise has been shown to increase the concentration of orexigenic neuropeptide Y in the metabolism centers in the hypothalamus (Lewis et al., 1993), more research is needed to explain the appetite control mechanisms triggered by exercise.
As regards water intake, our results are similar to those reported by Sharma et al. (2002) in rats, who observed an increase when they introduced the variable of physical activity to hypobaric hypoxia exposure. It is known that physical exercise produces a significant rise in body temperature, which elicits a heat loss response comprising increased blood flow to the skin and increased sweat secretion (Sawka et al., 2005). Combined with the increase in lung ventilation as a consequence of exercise, this explains the greater loss of water and consequently the compensatory higher water intake found in the exercised groups.
Oxygen consumption
Our results (Table 2) showed significant decreases in oxygen consumption in both hypoxic groups (HS and HE) after application of the chronic IHH protocol. Thus, whether applied alone or in combination with EET, this protocol decreased the basal metabolic rate when compared with baseline values recorded before IHH exposure. This was in clear contrast to results for the normoxic groups (NS and NE), where no significant differences in oxygen consumption were found before or after conducting the experiment. Taking all our data together, it could be argued that IHH induced a reduction in food intake (hypophagia) leading to a negative energy balance, which would explain the weight loss. The decrease in oxygen consumption could be the result of an energy conservation strategy similar to that described in studies on food-restricted rats as an adaptive mechanism to save energy (Luz et al., 2010).
There is some controversy in the literature about the effects of hypoxic exposure on basal metabolic rate. Several researchers have reported a reduction in oxygen consumption in some mammalian species during normobaric or hypobaric hypoxia (Hill, 1959; Gautier et al., 1989; Frappell et al., 1992; Nathaniel et al., 2012), which was accompanied by hypophagia (Alippi et al., 1983), whereas other authors have reported increases in the basal metabolic rate (Butterfield et al., 1992; Mawson et al., 2000; Lippl et al., 2010). In our opinion, two main factors could explain the reduction in oxygen consumption observed after the acclimatization protocol reported in this study.
On the one hand, several studies that have investigated the effect of hypoxia independently from other factors, such as cold or exercise, showed that hypoxia per se promoted a lipid to carbohydrate shift in the metabolic pathways increasing the activity of glycolytic enzymes, such as hexokinase and phosphofructokinase, and decreasing mitochondrial import and β-oxidation of fatty acids (see for review Favier et al., 2015). This could explain the observed reduction in oxygen consumption, since glycolytic pathways require less oxygen as compared with lipid oxidation. During acute exposure to hypoxia, it is well established that the defensive hyperventilation response leads to both increased water loss and blood alkalinization (Prabhakar, 2013). An increased use of carbohydrates can help to compensate these alterations. The higher respiratory quotient (RQ = 1) for carbohydrates, in contrast to that for lipids (RQ = 0.7), not only contributes to improve acid–base balance due to a higher CO2 production per mol of O2 consumed, but also increases alveolar PO2 during hypoxia (see West et al., 2013). Moreover, metabolic water production in relation to caloric value is also higher for carbohydrates (15 mL/100 kcal) than for lipids (11.8 mL/100 kcal) (West et al., 2013).
On the other hand, another possible explanation for the reduction in oxygen consumption would be a mechanism aimed at improving mitochondrial efficiency under hypoxic conditions. Previously published data on in vitro cardiac mitochondrial function in the same animals as those used in the present protocol (Magalhães et al., 2013, 2014) indicated that several key parameters associated with mitochondrial efficiency were significantly improved in the hypoxic groups. For example, the depolarization lag phase of the mitochondria was significantly lower in the HE group than in the other groups, indicating that less time was required to phosphorylate added ADP. Furthermore, after energizing in vitro mitochondria for different substrates, mitochondrial respiratory indices (RCR, respiratory control rate; and ADP/O, ratio) were significantly higher in hypoxic groups (and in the NE group for RCR) than in the control NS group. These indices, especially RCR, are considered the best general measures for in vitro mitochondrial function assays (Brand and Nicholls, 2011), and the higher values obtained indicate that hypoxic groups improved their coupling between respiration and ATP synthesis.
Finally, the combination of IHH and EET induced a significant increase in content of the mitochondrial protein ANT, which is a nucleotide translocator involved in ADP/ATP exchange across the mitochondrial inner membrane (Liu and Chen, 2013), thus playing an important role in mitochondrial bioenergetics. This better mitochondrial “fitness” in hypoxic groups could ensure good energy status under conditions of limited oxygen availability in tissues, resulting in a decrease in oxygen consumption. Hence, the decrease in total oxygen consumption could reflect changes in the cellular metabolism of different tissues, such as those found in the cardiac muscle.
Conclusion
Whether alone or combined with EET, IHH induced a reduction in food intake leading to a negative energy balance and weight loss concomitant with a decrease in oxygen consumption and basal metabolic rate probably to save energy. These results suggest that IHH+EET could be applied combined as a potential nonpharmacological tool for the management of overweight and obesity. Furthermore, IHH alone may present a promising strategy for overweight subjects with limited mobility.
Footnotes
Acknowledgments
The present study was supported by a research grant from the Research Center in Physical Activity, Health, and Leisure (CIAFEL) R&D Unit (PEst-OE/SAU/UI0617/2011) and partially by research grant ref. DEP2013-48334-C2-1-P from the Plan Nacional I+D+i (Spanish Ministry of Economy, Industry, and Competitiveness). The author Cabrera-Aguilera was supported by the CONICYT (Comisión Nacional de Investigación Científica y Tecnológica–Chilean National Commission for Scientific and Technological Research) “Becas Chile” Master Fellowship program; Grant No. 73170104.
Author Disclosure Statement
No competing financial interests exist.