
Abstract
Abstract
Introduction:
The adaptive response to systemic intermittent hypoxic therapy (SIHT) may be used for therapeutic advances due to the activation of multiple pathways involved in angiogenesis, immunomodulation, and tissue homeostasis. The aim of this study was to investigate the early age-dependent systemic response of different exposures of SIHT in mice.
Materials and Methods:
Sixty-four C57BL/6NRj female mice in three different age groups, young (4–5 weeks), adolescent (8–10 weeks), and adults (23–32 weeks), were exposed to SIHT. Different algorithms for equal hypoxic challenges (oxygen-decrease*time) were investigated to allow examination of the role of absolute hypoxia (oxygen-decrease) compared with relative hypoxia (total oxygen depletion over time). The systemic effects of angiogenetic regulation were investigated using blood samples analyzed by ELISA, proteome profiles, and proximity extension immunoassay. One-way analysis of variance with post hoc Bonferroni analyses was performed.
Results:
The early systemic response to SIHT was dependent on the absolute hypoxia rather than relative hypoxia over time. Serum erythropoietin (EPO) levels were increased significantly in young mice receiving low-oxygen SIHT treatments (10% and 15% oxygen). The expression of angiogenic proteins differed between the different age groups indicating an age-dependent response to SIHT. Focusing on hypoxia-inducible factor-1 (HIF-1) signaling, there was a trend toward upregulated angiogenetic response with younger age. Furthermore, clustering of protein expression in low-oxygen SIHT algorithms were found between young and adolescent mice. In adult mice, the majority of the proteins were downregulated as a response to SIHT. The systemic response of metabolites expressions was most pronounced in young mice. Systemic levels of cardiac troponin I (Tnni3) was unaffected by SIHT independent of age groups.
Conclusions:
The systemic response to SIHT is dependent on the absolute hypoxic exposure rather than the relative hypoxic depletion over time. Age-dependent effects of a short-term SIHT were associated with an increase in EPO, upregulation of angiogenetic pathways, and select metabolic and cell-surface proteins.
Introduction
Numerous systemic hypoxic treatment algorithms have been explored to improve athletic performance, but studies also suggest that the systemic biological adaptation to hypoxia-induced stress could be advantageous in therapeutic and regenerative medicine (Levine et al., 1992; Bakkman et al., 2007; Vogt and Hoppeler, 2010). Systemic intermittent hypoxic therapy (SIHT) is a ubiquitous stimulus that influences the vast majority of the physiological processes that are associated with altitude training. SIHT is defined as transient exposure to reduced oxygen availability in the air for a given period of time followed by a subsequent return to normoxia. The SIHT algorithms can be regulated by exposure time, number of intervals and rest periods, oxygen concentration, and duration of treatment. An immediate challenge for investigators is to identify the SIHT algorithm resulting in the most beneficial biological response and the lowest risk of potential pathological side effects, still within the limits of psychological treatment tolerance (Dale et al., 2014).
Many of the biological processes associated with a hypoxic challenge involves an adaptive response to ensure oxygen-homeostasis mediated by hypoxia-inducible factor (HIF) signaling in angiogenesis and erythropoiesis, shifting the metabolic pathways to compensate for the intrinsic response to hypoxic stress (Haase, 2010). Molecular response systems and processes are regulated at transcriptional and proteomic levels by oxygen sensing (Semenza, 1998). Erythropoietin (EPO), vascular endothelial growth factor (VEGF), and plasminogen activator inhibitor-1 (PAI-1) are among the adaptive genes responding to changes in oxygen availability.
Previous studies on age-dependent responses to hypoxia have demonstrated differences in catecholamine response and survival under hypoxia (Scremin et al., 1980; Mader et al., 1991). In aged compared with young tissues hypoxia led to reduced mRNA expression of target genes in the antioxidant defense system (Martin et al., 2002). Furthermore, hypoxic culture environments reduced not only transforming growth factor-β (TGF-β) receptors and expression indicating an age-dependent manner in cells but they also demonstrated a complete loss of responsiveness to hypoxia due to aging of the cells (Mogford et al., 2002). The etiology of aging is associated with oxygen deprivation in tissues. Collectively, these previous findings highlight that there may be an age-dependent response to SIHT. Inspired oxygen of 9%–16% at low-rate intermittence for limited time-periods has been associated with beneficial effects without pathological outcome, and the physiologically and biologically affected targets include the immune system, cardiovascular system, respiratory system, and nervous system (Dale et al., 2014; Navarrete-Opazo and Mitchell, 2014).
In the present study, we aimed at investigating age dependence of the systemic responses to hypoxia by exposure to SIHT using different algorithms with a constant hypoxic challenge (oxygen-decrease*time). We hypothesized that the early adaptive response to systemic intermittent hypoxia is dependent on the absolute hypoxia (oxygen tension) and with age.
Methods
Ethical approval
The experiments were conducted and approved under the guideline given by The Danish Animal Experiment Inspectorate (Case No. 2016-15-0201.00926). This study conformed to the Danish law for animal research (Act No. 1306 of 23/11/2007, Danish Ministry of Justice). All animals were housed at the animal facility at the Aarhus University Hospital, Department of Clinical Medicine according to the guidelines and procedures approved by the Danish Animal Experiments Inspectorate.
Study design
Sixty-four female mice were divided into three age groups: young (4–5 weeks old; n = 16), adolescent (8–10 weeks old; n = 24), and adults (23–32 weeks old; n = 24) and exposed to one of three different SIHT algorithms or a normoxic control environment and followed for 48 hours after the beginning of the first exposure. The different hypoxic challenges were calculated from an algorithm using oxygen decrease*exposure time*number of exposures. The selected treatments provided the similar total oxygen deprivation over time (area under the curve; Fig. 1). To apply the different hypoxic treatments, a normobaric hypoxic chamber was adjusted to replace atmospheric oxygen by reducing oxygen through a nitrogen membrane unit. The system was set at oxygen concentrations of 18%, 15%, and 10%. The SIHT algorithms were selected to provide a constant oxygen depletion: 18% for 640 minutes, one time/day, 1 day (+1 day rest); 15% for 160 minutes, one time/day, 2 days; and 10% for 30 minutes, three times/day, 2 days. Arterial blood was collected by cardiac puncture via diaphragm from the left ventricle of the heart. Immediately after collection of blood samples, a blood smear was made and air-dried. Serum was purified from whole blood, where the blood was allowed clotting by leaving it undisturbed at room temperature for 15–30 minutes, before centrifugation at 1000 g for 10 minutes. Serum samples were obtained and stored at −20°C.
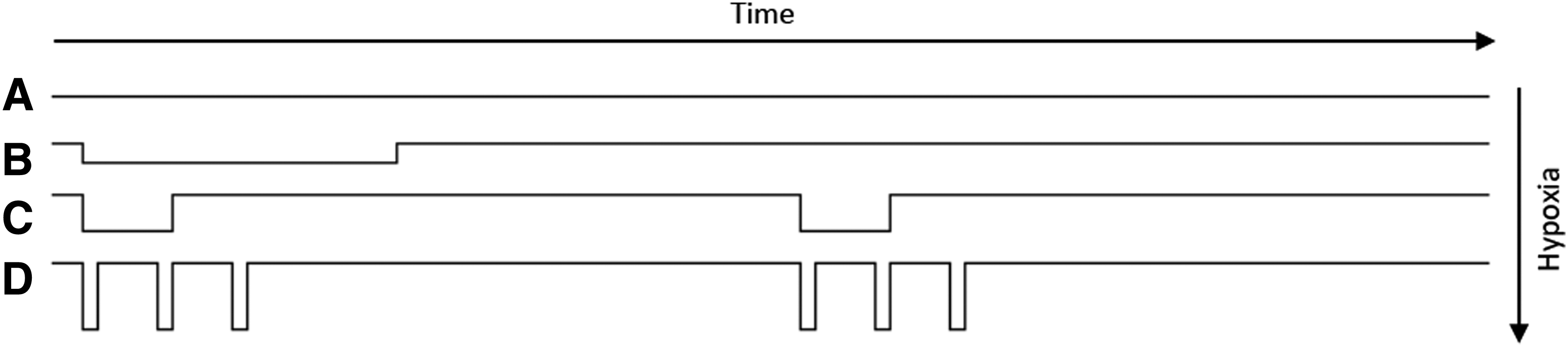
Hypoxia exposure over time. Visualizing area under the curve. Vertical axis represents different hypoxic strategies and thereby oxygen concentrations. Horizontal axis represents time as 2 days.
Animals
In this study, 4–32 week old female C57BL/6NRj mice were used (Janvier Labs, Le Genest-St-Isle, France). The mice were housed in certified animal facilities in a 12-hours dark:light cycle with unlimited water and standard chows supplies. At follow-up, the mice were euthanized by exsanguination through the left ventricle of the heart under sevoflurane inhalant anesthesia in a drop jar (Abbott Scandinavia, Solna, Sweden). Cervical dislocation was performed secondarily to assure euthanasia.
Serum EPO
EPO is synthesized by interstitial fibroblasts in the kidneys as a response to low oxygen and is thus used as a well-established marker of systemic sensation of hypoxia. Quantification of serum EPO levels were measured using the Quantikine ELISA Mouse Erythropoietin kit (MEPOOB); R&D Systems, Minneapolis, MN). The optical density was measured using microplate reader at 450 nm and the amount was calculated from a multiparameter logistic curve fit. Measurements were performed according to the manufacturer's instructions.
Leucocytes
Whole blood smears were air-dried and stained with May–Grünwald–Giemsa method. Leucocytes were manually counted using an × 100 objective in 20 fields of monolayer smear using Axioplan Zeiss Microscopy. All leucocytes counts were performed by the same experienced laboratory technician.
Angiogenesis array
Serum samples were used and the arrays were washed and incubated with biotinylated detection antibodies according to the manufacturers. IRDye 800CW Streptavidin (LI-COR Biosciences, NE) 1:2000 was applied and the signal intensity of each spot corresponding to phosphorylated protein was determined using Odyssey® SA far-red Imaging (LI-COR Biosciences) at 800 nm, resolution 50 μm, intensity 7, and Image Studio Software version 2.1.10 (LI-COR Biosciences) (Joergensen et al., 2016).
Proximity extension immunoassay
Olink MOUSE EXPLORATORY assay (Proseek Multiplex; Olink Proteomics, Uppsala, Sweden) was used for the assessment of proteins influenced by SIHT at different ages. BioXpedia (Aarhus, Denmark) analyzed 92 selected proteins from the blood samples. Each mouse exploratory protein's function description, analytical measuring range, and validation data are available from Olink. This provides an exploratory unbiased approach to ensure nonexclusion of potential novel target proteins potential molecular interactions involved in the selected mechanisms. In brief, 1 μL serum sample was used for proximity extension (PE) immunoassay based on antibodies linked to DNA oligos analysis (Lundberg et al., 2011; Assarsson et al., 2014; Greenwood et al., 2015). Olink NPX Manager software was used for processing of data. Data are presented as log2-transformed arbitrary units normalized protein expression (NPX). Public-access bioinformatics databases, including UniProt, Gene Ontology (GO), KEGG pathways, and STRING, were used for the understanding of the biological relevance of the differently expressed proteins.
Statistical analysis
All data are presented as the mean ± standard deviation (SD). The statistical analyses were performed using STATA Statistical Software (Vers. 13.1; StataCorp LP, Collage Station, TX). All data were checked for normality using histograms and QQ plots. One-way analysis of variance followed by Bonferroni post hoc test was performed on EPO, percentage of leucocytes and NPX data. Data are presented as mean values with SD. A significance level of p < 0.05 was used.
Results
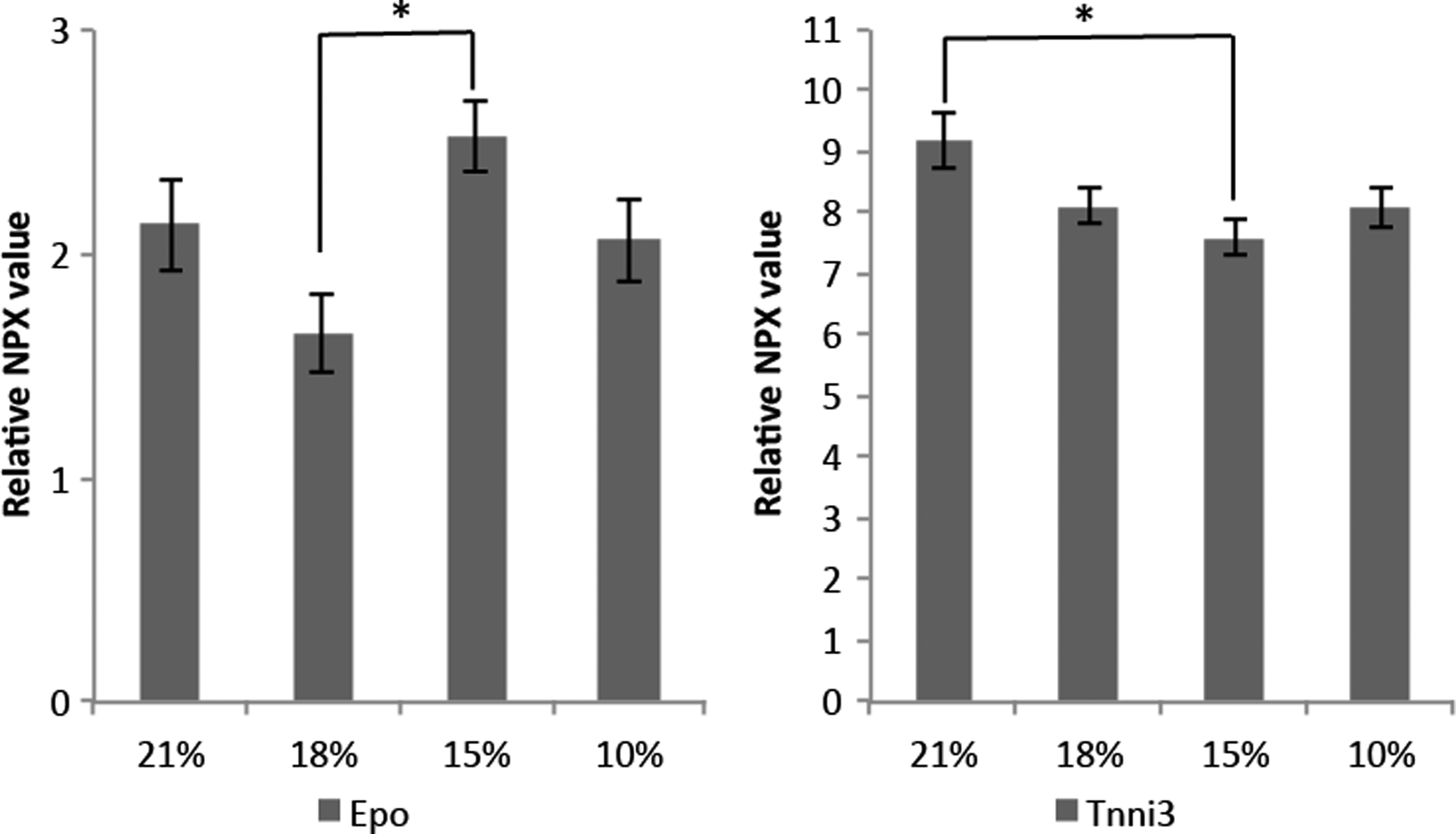
Serum EPO increased significantly for young mice with decrease in oxygen level and was significantly higher in 15% and 10% of oxygen comparing adolescent and adult mice. No significant effects were observed within the adolescent and adult mice groups (Fig. 2). There were no differences in the percentage of leucocytes among the SIHT treatments in young mice.
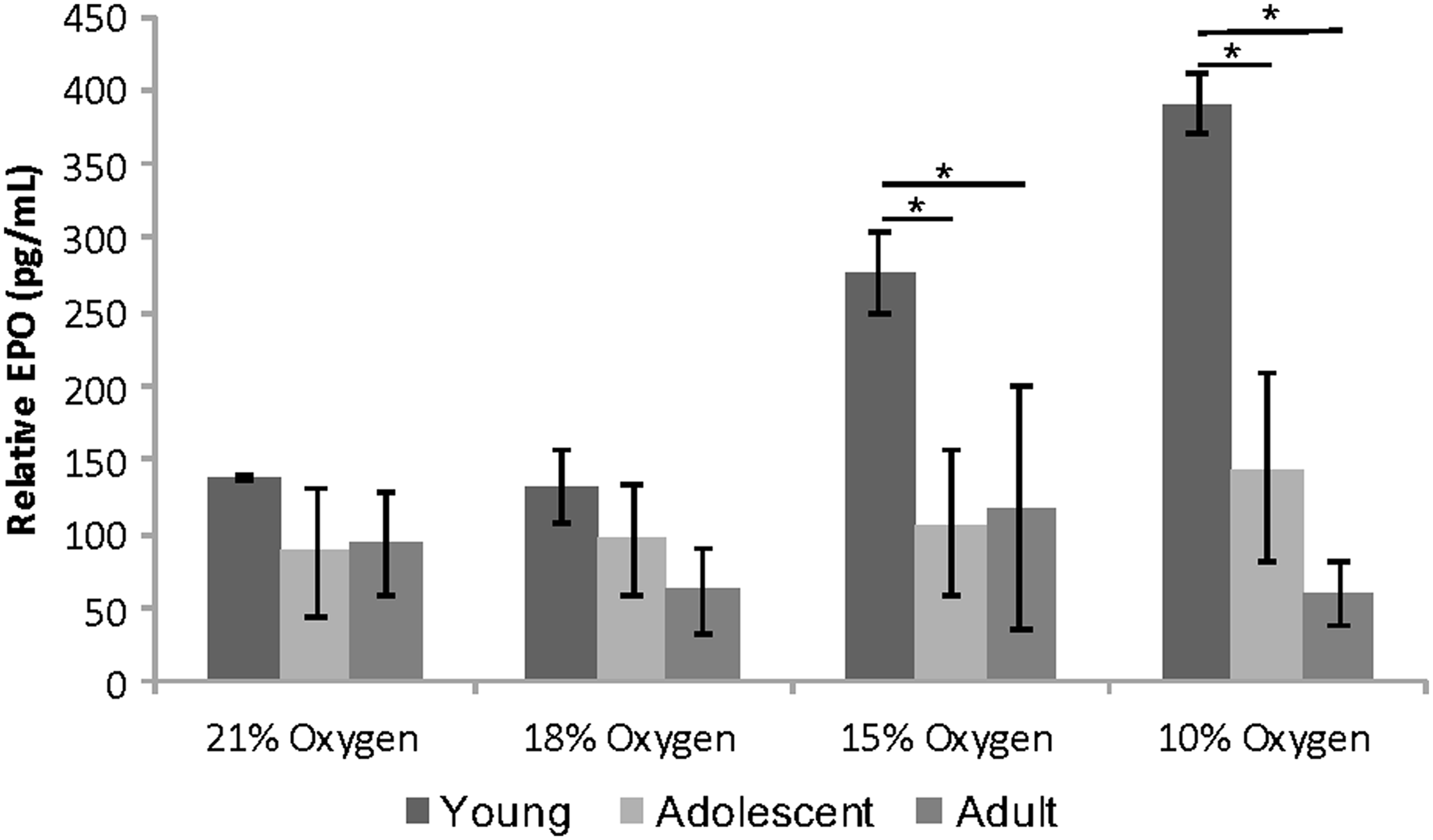
Relative EPO expression in serum. Vertical axis represents relative EPO concentration in serum. Horizontal axis represents different SIHT denominated by the oxygen concentration. Data are presented as mean ± SD. *Represents significant differences. EPO, erythropoietin; SD, standard deviation; SIHT, systemic intermittent hypoxic therapy.
To establish an understanding of angiogenic candidate proteins that may be influenced by SIHT, we investigated changes of angiogenesis-related proteins at different ages. Fifty-three proteins were screened, and young mice exposed to 10% and 15% hypoxia showed more similar heatmap expression compared with 18% hypoxia (Fig. 3A). The majority of the investigated proteins were downregulated in adult mice, especially in 15% and 18% SIHT. The expression pattern in the heatmap was not mirrored between the ages of mice, indicating different responses to hypoxia depending on age. Interestingly, the expressions grouped very similar within each age group with the lowest order of grouping found between ages (Fig. 3A). A comparison of the global volume of protein expression between the different hypoxia conditions showed a large variation in up- and downregulated expression depending on age (Fig. 3B). The expression patterns were most similar between young and adolescent mice compared with adult mice.
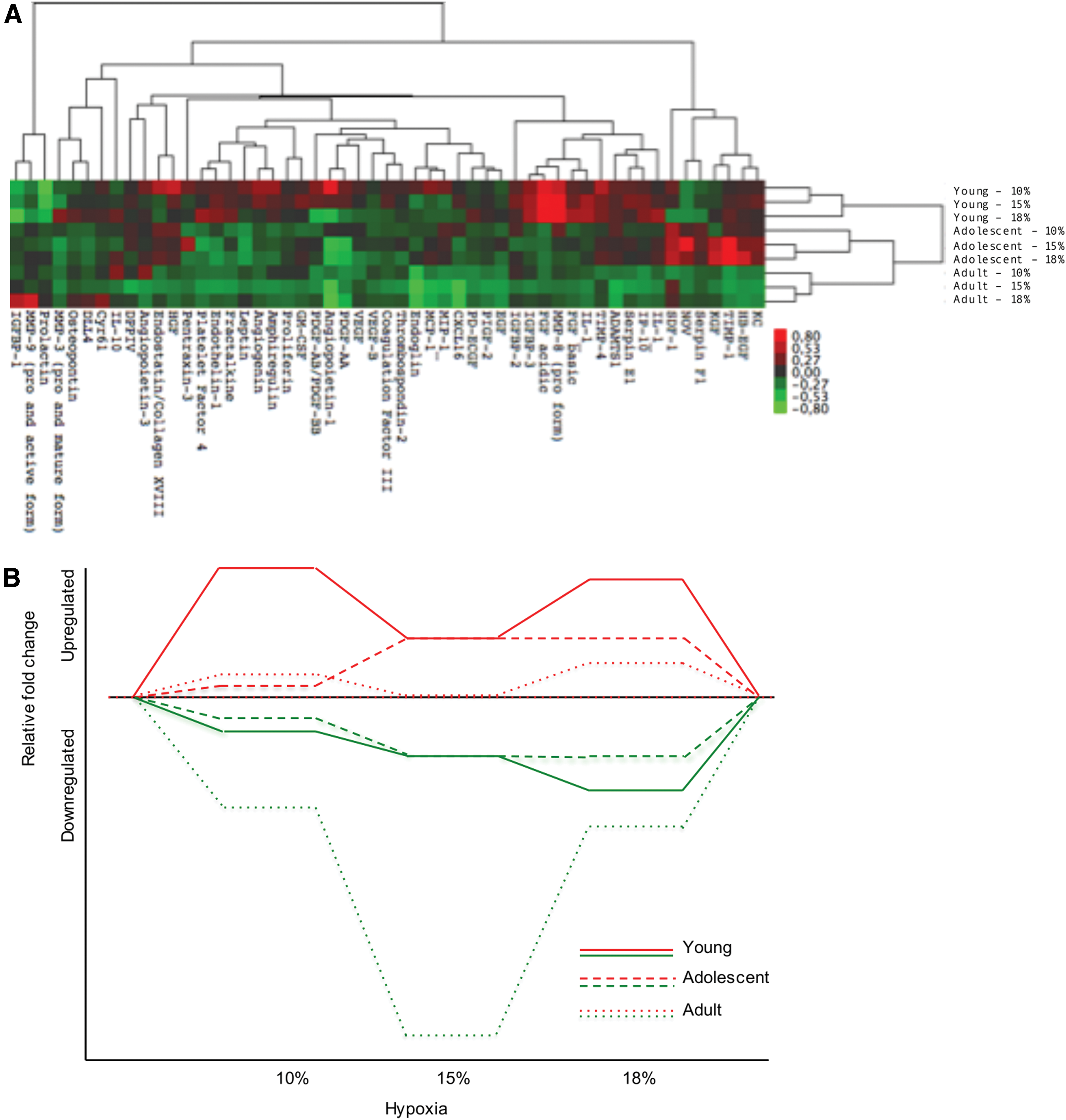
Proteomic assessment of angiogenesis-related proteins.
Subsets of the proteome array were analyzed individually. The HIF-1-dependent signaling in angiogenesis were investigated comparing VEGF, epidermal growth factor (EGF), tissue inhibitor of metalloprotease-1 (TIMP-1), SERPINE1 (PAI-1), and angiopoietin-1 (Ang-1) for different SIHT relative to controls at 21% oxygen (Fig. 4A). HIF-1 signaling in adult mice continued to cluster differently from the young and adolescent mice. Inflammatory cytokine expression subsets were also investigated individually for interleukins 1α, 1β, and 10 (IL-α, IL-1β, and IL-10) and delta-like 4 (DLL4) (Fig. 4B) in response to age and hypoxic conditions. There was a tendency for these inflammatory markers to be upregulated with increasing age and decrease in absolute oxygen, age being the most significant factor.
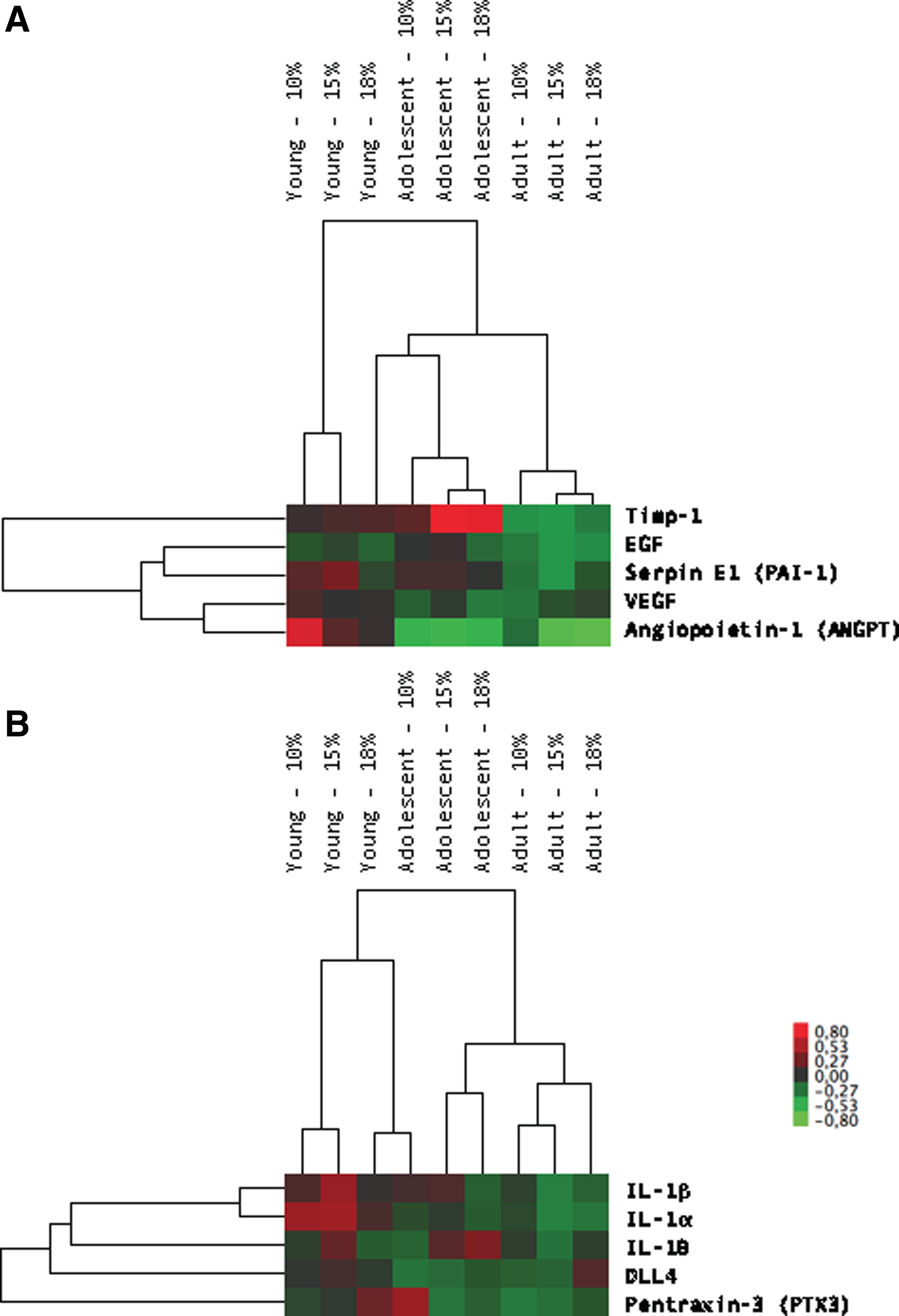
Hierarchal clustering of HIF-1 signaling pathway in angiogenesis
Further analysis identified 20 proteins that were significantly influenced in at least one of the SIHT algorithms in a particular age group. Increased age tended to result in a lower number of proteins significantly regulated by SIHT. The differentially expressed proteins were predominantly assigned to on the metabolic processes and processes related to the extracellular region. Many of the proteins were localized extracellularly (Table 1). Among the proteins that were more different in young mice were metabolites such as Parp1, Cpe, Erbb4, Qdpr, and Lpl. The primary metabolic processes involved were cellular amino acid, nucleobase-containing compounds and proteins (Fig. 5). For adolescent mice, a few metabolites were differently expressed and the nearest predicted functional partners in STRING version 10.5 were inflammatory mediators such as IL-1β, IL-6, and IL1r-1/2 (Supplementary Fig. S1). In adult mice, metabolites involved in lipid homeostasis was involved and visualized as the nearest predicted functional partners in STRING version 10.5 (Supplementary Fig. S2).
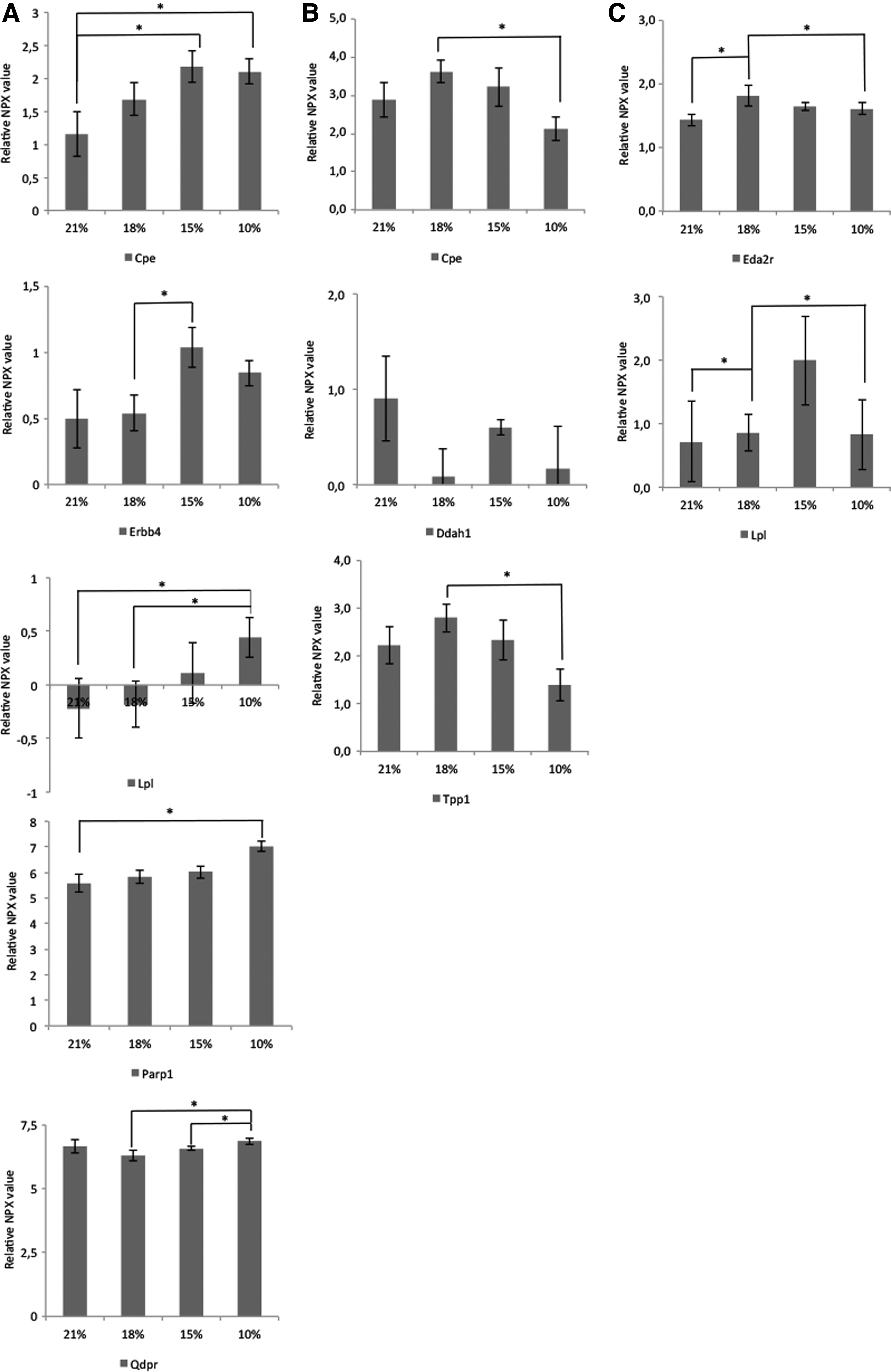
Relative NPX of metabolic process-related proteins for young
Proteins of Interest that Demonstrate an Influence of Age
Vertical columns include: Age, UniProt accession number, protein name, biological location, and p-values from one-way ANOVA test comparing different hypoxia conditions.
ANOVA, analysis of variance.
Cpe, Erbb4, Lpl, Parp1, Qdpr, Ddah1, Tpp1, and Eda2r were increased in all ages (Fig. 6). In young mice, many of the proteins related to the metabolic process increased with increasing degree of hypoxia. For instance, 10% and 15% hypoxia had significantly higher protein expression compared with 21% and 18%. However, a reverse tendency was observed for adolescent mice where metabolism-related proteins decreased the expression with increasing hypoxia. Selected proteins such as Cpe and Lpl were overlapping between the ages.

Relative NPX of cell surface receptor signaling pathway in young mice. Vertical axes represent NPX value on log-2 scale. Horizontal axes represent hypoxia conditions corresponding to different degrees of SIHT and specific proteins related to cell surface receptor signaling pathway. Significance (*) represents differences between SIHT in young mice.
Proteins associated with cell-surface receptor signaling were identified in young mice. As shown in Figure 7, 15% SIHT deviated from the other SIHT conditions on several of the eight-cell surface receptor signaling pathway proteins such as Cpe, Erbb4, Fst, Il23r, Notch3, Tgfa, Wfikkn2, and Wisp1. Interaction extracted from STRING version 10.5 indicated that Erbb4 and Tgfa had binding interactions and the predicted functional partners would be the Erbb signaling pathway (KEGG Pathways 4012), which are involved in angiogenesis (Strong Russell et al., 1999; Junttila et al., 2000; Szklarczyk et al., 2017).
Relative NPX of Tnni3. Vertical axis represents NPX value on log-2 scale. Horizontal axis represents hypoxia conditions corresponding to different degrees of SIHT. *Represents significant differences.
In addition, age-dependent proteomic analysis resulted in the significant changes in expression of cardiac troponin I (Tnni3) (ischemic marker) proteins comparing the different SIHT conditions (Fig. 7). Tnni3 as well as EPO are associated with hypoxia and according to STRING version 10.5 vasculogenesis (GO:0001570) and cardiac muscle tissue development (GO:0003229) (Supplementary Fig. S3).
Discussion
We applied a descriptive unbiased investigation of systemic proteins using a highly sensitive exploratory multiplex PE analysis to show that effect of SIHT on selected metabolic, inflammatory, and angiogenic proteins was dependent on the absolute decrease in oxygen and on the age of the recipient. Proteins were expressed to a higher degree with young age and a higher absolute number of proteins regulated were seen in young compared with the older animals. Different algorithms for oxygen deprivation were applied, and we found that although all treatments had similar total oxygen deprivation over time, the largest systemic response was observed with the largest absolute decrease in oxygen concentration. Angiogenesis-associated signaling was among the highly regulated, while the metabolic processes and cell-surface receptor systems (EGF family; Erbb4 and Tgfa) were affected by SIHT.
Mitochondria are organelles with high oxygen demands to maintain the energy consumption of the respiratory chain (Warburg, 1956). They are also considered major stakeholders of aging processes due to the formation of reactive oxygen species (Wu et al., 2018). We observed increased metabolic protein expression in SIHT algorithms with lowest absolute oxygen levels in young mice, whereas adult mice exhibited the reverse tendency by decreased protein expression in low oxygen. The increases in metabolic proteins are a double-edged sword since some of the metabolic proteins have been suggested as biomarkers in diseases. For instance, increased levels of follistatin have been associated with low-grade inflammation in patients with diabetes mellitus type 2 (Hansen et al., 2013); WNT1 inducible pathway protein 1 has been correlated with the severity of renal fibrosis (Zhong et al., 2017); and carboxypeptidase E as a biomarker for pulmonary neuroendocrine tumor (He et al., 2004). According to the GO annotation database, 10 genes have been associated with a negative regulation of cellular response to hypoxia, and among these are Ddah1 (Blake et al., 2017). In adolescent mice, Ddah1 was reduced by exposure to SIHT compared with 21% oxygen, and this has previously been associated with the expression of VEGF and tumor hypoxia (Buijs et al., 2017). The energy consumption in hypoxic conditions switches to glycolysis. Cpe was elevated in young and adolescent mice at 15% and 18% oxygen indicating metabolic plasticity due to hypoxia, and this has been shown to enhance the glucose flux into the citric acid cycle as seen in cancer cells (Ilina et al., 2017). Systematic pathway analyses are required to clarify the presence of causality in our design.
Some proteins displayed enhanced interaction indicating a biological connection. For instance, a link between the Erbb signaling has previously been associated with hypoxia in young mesenchymal stem cells (MSCs) compared with aged MSCs (Mohd Ali et al., 2016). We suggest a SIHT-dependent elevated level of systemic EPO and reduced level of Tnni3. Serum EPO was high after 10% and 15% SIHT in young mice, and it was demonstrated to be age-independent. Interestingly, the HIF-1 signaling pathway plays a central role in inducing gene products of TGFα, EPO, and EGF receptor signaling. Hypoxia-induced increase in EPO synthesis has been shown to exert protective effects such as memory improvement, antidepressive behavior, and ischemia-reperfusion (Cai et al., 2003; Miskowiak et al., 2012). The reduced response of HIF-1 in adult mice is in agreement with earlier results demonstrating impaired HIF-1 stabilization with aging (Ceradini and Gurtner, 2005). The elegance of modulating the HIF-1 pathway using SIHT could benefit the presence of systemic stem cells as HIF-1 is an upstream regulator of stem cell biology potentially augmenting endogenous physiological pathways in tissue repair (Ceradini and Gurtner, 2005).
To use SIHT therapeutically, it is important to acknowledge that we did not observe any increase in the cardiac myocyte ischemic marker, Tnni3, compared with 21% oxygen. Tnni3 has been associated with acute and chronic heart failure (Sato et al., 2009).
The present study is limited to the types of SIHT treatments investigated and does not provide any evidence of the age dependence of longer exposure periods and the systemic responses to adaptation. The oxygen saturation curve in mice is right-shifted compared with humans, and the mass-specific metabolic rate is seven times larger in mice than in humans (Demetrius, 2005). To reduce biological variance, we only used female mice and the magnitude of the early adaptive response to hypoxia in males may be different. Using different algorithms of SIHT, we applied hypoxia in single or multiple dosages. Hence, the addition of multiple hypoxic challenges could potentially bias the results. These are important factors that should be accounted for in attempts for clinical translation.
Conclusion
We have shown that a profound systemic response can be induced by exposure to SIHT and that the response is dependent on age and the absolute decrease in oxygen level. Angiogenesis-associated signaling was among the highly regulated, while the metabolic processes and cell-surface receptor systems were also affected by SIHT.
Footnotes
Acknowledgments
We gratefully thank laboratory technicians Anna Bay Nielsen and Anette Baatrup, Orthopaedic Research Laboratory, Aarhus University Hospital, Denmark for the excellent technical assistance provided and BioXpedia, Aarhus, Denmark, for the Olink proximity extension immunoassay. This study was funded by Aarhus University Research Foundation NOVA grant (AUFF-E-2015-FLS-9-7).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.