
Abstract
Abstract
Introduction:
Cognitive function has been compromised during high-altitude (HA) exposure due to slowing of mental processing.
Materials and Methods:
A total of 20 Indian and 20 Kyrgyz soldiers were studied at 4111 m to assess cognitive function in two different ethnic groups. Paired associate learning, pattern recognition memory, spatial span (SSP), spatial working memory (SWM), choice reaction time (CRT), and simple reaction time (SRT) were evaluated at sea level and on days 3, 7, 14, and 21 of HA stay and on day 3 of deinduction.
Results:
All the parameters were significantly affected at HA. Indian soldiers were acclimatized by 7 days but Kyrgyz soldiers required 21 days for acclimatization. A slow impairment in SWM, CRT, and SRT was observed in Kyrgyz soldiers than in Indian soldiers and it continues throughout 21 days of HA stay, but for Indian soldiers the deterioration was maximum on day 7 and improvement in SWM, CRT, and SRT was observed on day 14 and close to baseline value on day 21. After deinduction, although Indian soldiers attained the normal value, Kyrgyz soldiers had higher value than baseline in SSP, SWM, CRT, and SRT.
Conclusion:
Difference in the cognitive performances of Indian and Kyrgyz soldiers may be due to the ethnogenetic diversity of these two groups.
Introduction
Hypoxic hypoxia causes impairments in different components of cognitive functioning, that is, ability to learn, remember, express information verbally (Towners et al., 1984; Muthuraju and Pati, 2014; Phillips et al., 2017), concentration, cognitive flexibility (Regard et al., 1989), spatial memory (Cavaletti et al., 1990; Shukitt-Hale et al., 1994), and fear memory (Kumari et al., 2018). Even acute mild hypobaric hypoxia affects cognitive performance (Bouak et al., 2018). Animal studies have revealed that CA1, CA3, and entorhinal cortex of hippocampus and layer II of prefrontal cortex are the most vulnerable to hypoxic damage (Maiti et al., 2008).
Neuropsychological testing revealed a decline in visual long-term memory after ascent to altitudes between 5488 and 8848 m (Hornbein et al., 1989). Shukitt-Hale et al. (1996) reported that neuronal damage in human brain after chronic exposure to hypobaric hypoxia persists even for a year or longer (Hornbein et al., 1989; Cavaletti et al., 1990; Cavaletti and Tredici, 1993) after deinduction, and a marked reduction was observed in short-term memory after exposure to hypobaric hypoxia for 1 hour at 4400 m (Shukitt-Hale et al., 1996). Lemos et al. (2012) reported that after 24 hours exposure in a normobaric chamber simulating an altitude of 4500 m reduces the quality and quantity of sleep; increases depressive mood, anger, and fatigue; and also reduces attention, visual and working memory, concentration, executive functions, inhibitory control, and speed of mental processing. In contrast to these findings, no significant change was observed in cognitive function in simulated altitudes up to 5500 m (Kennedy et al., 1989) and other experiments on Mount Everest at 6600 m (Nelson et al., 1990).
The central mechanisms responsible for the effect of hypobaric hypoxia on cognitive performance are not clear. The increase in event-related potential P300 latency indicated that long duration of stay (6 months) at high altitude (HA, 4115 m) slows the stimulus evaluation and signal processing (Thakur et al., 2011). It is likely that the direct effects of mild transient hypoxia on brain cause variations in the level of specific neurotransmitters involved in cognitive processes (Freeman and Gibson, 1988) because the experimental studies on rats showed alterations in synthesis of monoamines under simulated hypoxia (Ray et al., 2011). These heterogeneous results might be due to ethnic variation of different experimental populations.
It is well established that ethnic disparities exist in the slope of decline in cognitive function (Masel and Peek, 2009). Boivin (1991) reported that Scottish children performed significantly better than the Zairian children on the visual–spatial task. Mexican American ethnicity is a significant variable associated with poor cognitive functioning on a battery of screening tests, including domains associated with frontal/executive functions than non-Hispanic white elderly residents of El Paso County (Heller et al., 2006). Gurland et al. (1999) reported that cognitive impairment was more common in a group of Caribbean Hispanics in New York City than among non-Hispanic whites. Irrespective of ethnic variation, cognitive and neuropsychological functions have been impaired at HA and the effects depend on altitude and duration of stay.
Thus, in this study, we assessed the cognitive functions of two different ethnic groups (Indian and Kyrgyz soldiers), residents of different geographical location, and evaluated under similar conditions of HA exposure.
Materials and Methods
Subjects and study protocol
The study was conducted on 20 Indian and 20 Kyrgyz soldiers in the age group 21 to 30 years (mean ± SD = 25.2 ± 3.05) and 21 to 32 years (mean ± SD = 24.5 ± 4.43), respectively, with an educational qualification of higher secondary level.
They were lowlanders with no previous exposure to HA. Indian soldiers were flown to Bishkek, Kyrgyzstan, and after 2 days of rest, baseline recording in both Indian and Kyrgyz soldiers was carried out at Bishkek (800 m above sea level), Kyrgyzstan on day 3 and thereafter, they were inducted to an altitude of 4111 m (Syok Pass, Kyrgyzstan) for HA studies. Soldiers were inducted to altitude by road and it took about 3 hours to arrive at 4111 m from Bishkek. After a stay of 21 days, soldiers were returned back to Bishkek for deinduction studies (Fig. 1). Atmospheric temperature was ranging from 28°C to 35°C at Bishkek and from 5°C to 20°C at Syok Pass, whereas inside the rooms the temperature was maintained at 22°C ± 2°C.
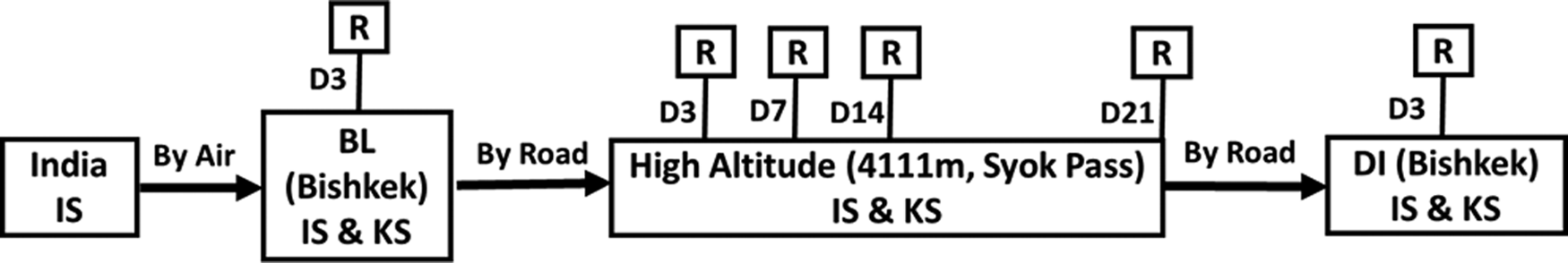
Schematic diagram of experimental protocol. Indian soldiers (IS) reach Bishkek by air, and baseline (BL) recordings (R) were performed in both IS and Kyrgyz soldiers (KS) on day 3 (D3). Soldiers were transported by road from Bishkek to 4111 m altitude (Syok Pass) where they stayed for 21 days. High-altitude recordings were carried out on day 3 (D3), day 7 (D7), day 14 (D14), and day 21 (D21). After that, soldiers were deinducted (DI) to Bishkek by road and all the parameters were recorded on day 3.
At HA, proper winter clothing was provided to all the soldiers. Soldiers followed proper acclimatization schedule and engaged themselves in indoor games, watching television, playing table tennis, billiard, etc., along with morning drill, walking, and other galley duties at HA. The purpose of the experiment was explained to all of them and all gave individual written informed consent to participate in the study. The protocol of the experiment was approved by the Ethics Committee of the Institute. All the guidelines of Helsinki protocol were followed. The participants were free from any neurological disorder and cardiac problems. They were not taking any sort of psychotropic, psychoactive, or other drugs during the course of the study. Studies were carried out in both Indian and Kyrgyz soldiers on day 3 after reaching Bishkek (baseline), on days 3, 7, 14, and 21 after induction to HA, and on day 3 of deinduction to baseline.
Arterial oxygen saturation measurement
Saturation of peripheral oxygen (SpO2) was measured using finger pulse oximeter (Nonin Medical, Inc.).
Cognitive function
For the evaluation of cognitive function, Cambridge Automated Neuropsychological Test Battery (CANTAB) (Torgersen et al., 2012) was used, which is a semiautomated neuropsychological test battery applied on a laptop PC, developed at the University of Cambridge. CANTAB contains different neuropsychological tests in five cognitive domains: visual memory, semantic/verbal memory, decision-making and response control, executive function, and attention.
Motor screening test
Motor screening test was conducted first to screen for ability to cooperate with the apparatus. Soldiers were instructed to point on a flashing cross as soon as it appears (Fig. 2).

Pictorial diagram of motor screening test. A series of crosses were presented at different locations on the screen. The subject must touch a cross properly that will disappear with an alarm sound leading to appearance of the next cross on the screen.
Paired associate learning
Paired associate learning (PAL) is a test of episodic and visual memory but also depends on the ability of spatial planning. The performance on PAL depends on input mainly from the temporal lobe, but also from the frontal lobe. Soldiers had to remember the location of different patterns appearing on the screen and then point out where on the screen the pattern initially was shown (Fig. 3). The difficulty level was increased ranging from two to eight patterns to be remembered (Sahakian et al., 1988).

Pictorial diagram of paired associate learning. Boxes were displayed on the screen and were opened in a randomized order. One or more of them will contain a pattern. The patterns shown in the boxes were then displayed in the middle of the screen, one at a time, and the subject must touch the box where the pattern was originally located.
Pattern recognition memory
Soldiers were required to learn a series of 12 simple, but abstract, colored visual patterns located in the center of the screen. Each pattern was presented for 3 seconds. In the recognition phase, 12 pairs of colored patterns were then presented on the screen one at a time and the soldier had to identify the target pattern from a “distracter” pattern. This procedure was repeated with 12 new patterns, and the soldier was given a total score (maximum = 24) expressed as a percentage (Mehta et al., 1999). This test is sensitive to dysfunction in medial temporal areas of the brain and is relatively insensitive to dysfunction in the frontal lobe (Fig. 4).

Pictorial diagram of pattern recognition memory. Subjects were presented with series of visual patterns, one at a time, in the center of the screen. These patterns were designed so that they could not easily be given verbal labels. In the recognition phase, the subject was required to choose between a pattern they have already seen and a novel pattern.
Spatial span
Spatial span (SSP) is a computerized version of the Corsi Blocks task, assessing working memory capacity. This gives a measure of frontal lobe functioning. A pattern of white boxes is shown on the screen. Some of the boxes change in color, one by one, in a variable sequence. At the end of the presentation of each sequence, a tone indicates that the soldier should touch each of the boxes colored by the computer—in the same order as they were originally presented. In the reverse mode, the tone indicates that the soldier should touch each of the boxes colored by the computer—in the reverse order to the order in which they were originally presented (Fig. 5). In both modes, the number of boxes in the sequence is increased from a level of two at the test to a final level of nine (Owen et al., 1990).
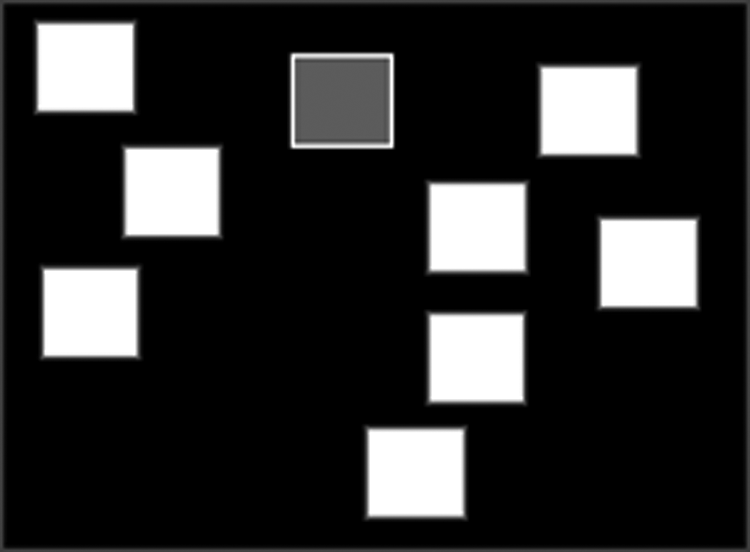
Pictorial diagram of spatial span. Few white boxes appear on the screen that change color one by one in random order. The subjects have to remember the sequence and give their response after a beep by touching the boxes in the correct order.
Spatial working memory
Spatial working memory (SWM) is a test of the subject's ability to retain spatial information and to manipulate remembered items in working memory. It is a self-ordered task, which also assesses heuristic strategy. This test is a sensitive measure of frontal lobe and “executive” dysfunction. The test begins with a number of colored squares (boxes) being shown on the screen. The aim of this test is that, by process of elimination, the soldier should find one blue “token” in each of a number of boxes and use them to fill up an empty column on the right hand side of the screen. The number of boxes is gradually increased from 3 to 8 boxes (or to 10 boxes in the second mode). The color and position of the boxes used are changed from trial to trial to discourage the use of stereotyped search strategies (Fig. 6).
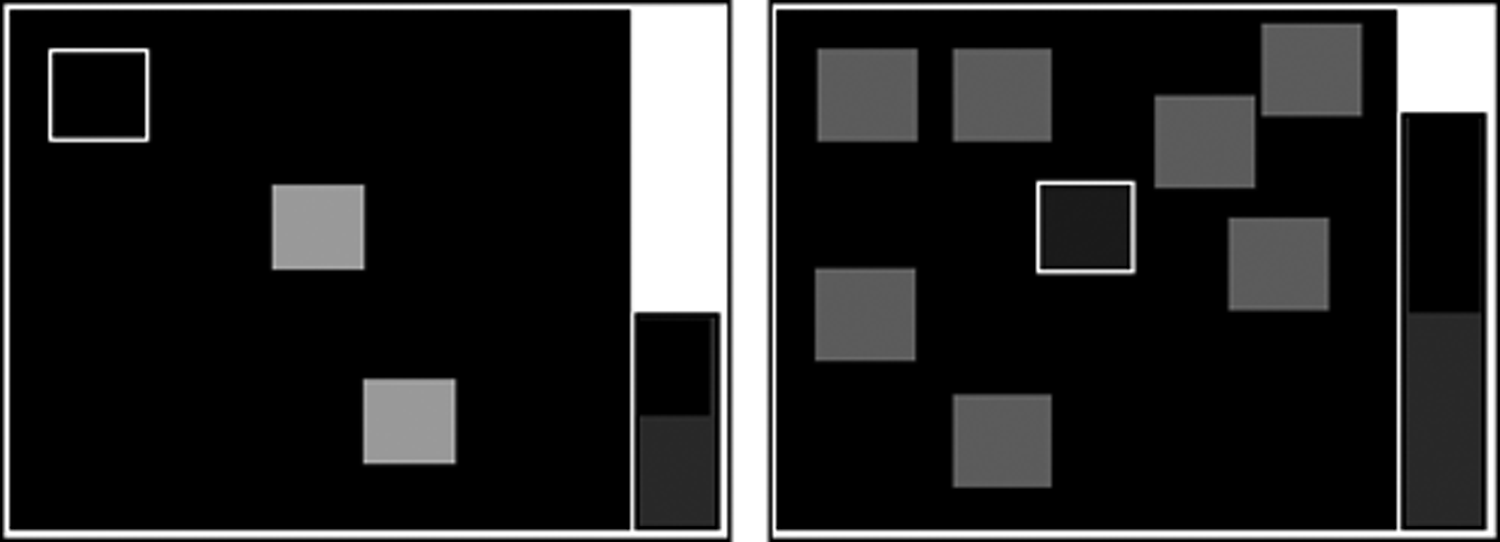
Pictorial diagram of spatial working memory. Some colored boxes appear on the screen. The subject has to look for a blue token that the computer has hidden inside one of the boxes. Only one token will be hidden at a time. If the subject did not find the blue token inside the box he touched, he should look for the token in another box. Subjects should collect enough blue tokens to fill the black hole (home) on the right on the screen.
The soldier must touch each box in turn until one opens with a blue token inside (a search). When a blue token was found, the soldier has to place it in the right column (“home”) by touching the right-hand side of the screen. The soldier must then begin a new search for the next blue token. It may be in any of the boxes that so far have been empty. This is repeated, until a blue token was found in every box on the current screen. Touching any box in which a blue token has already been found is an error. The soldier decides the order in which the boxes are searched. The computer determines the number of empty boxes that must be visited (discounting errors). Performance at the harder levels of this task is enhanced by the use of a heuristic search strategy (Owen et al., 1990).
Choice reaction time
Choice reaction time (CRT) is a 2-CRT test. An arrow-shaped stimulus is displayed on either the left or the right side of the screen. The soldier must press the left hand button on the press pad if the stimulus is displayed on the left hand side of the screen, and the right hand button on the press pad if the stimulus is displayed on the right hand side of the screen (Fig. 7).

Pictorial diagram of choice reaction time. An arrow appears on the screen. When the arrow is pointing to the left, a subject has to press the left button and when the arrow is pointing to the right, the subject has to press the right button.
Simple reaction time
Simple reaction time (SRT) is a test that measures SRT—through delivery of a known stimulus to a known location to elicit a known response. The only uncertainty is with regard to when the stimulus will occur, by having a variable interval between the trial response and the onset of the stimulus for the next trial (Fig. 8).

Pictorial diagram of simple reaction time. A square appears on the screen. As soon as it appears, a subject has to press the button on the press pad as quickly as possible.
Statistical analysis
Values are expressed as mean ± SEM. The data were analyzed using Prism 5 software (Graph Pad). To compare the changes in the same group in different phases, data were analyzed using one way analysis of variance with repeated measure and post hoc testing with Bonferroni multiple comparison test. p Value <0.05 was considered significant.
Results
Arterial oxygen saturation measurement
In case of Indian soldiers, SpO2 was significantly decreased after 3 days (87 ± 0.5; p < 0.001) of HA induction than baseline (BL, 98 ± 0.1). On day 7, SpO2 was found to be 90 ± 0.5 (p < 0.001 than BL), which further increased to 91 ± 0.4 (p < 0.001 than BL) and 92 ± 0.3 (p < 0.001 than BL) on days 14 and 21, respectively.
In case of Kyrgyz soldiers, SpO2 level was significantly decreased after 3 days (88 ± 0.7; p < 0.001) of HA induction than BL (98 ± 0.2). On day 7, SpO2 was found to be 88 ± 0.7 (p < 0.001 than BL), which further increased to 91 ± 0.4 (p < 0.001 than BL) and 92 ± 0.4 (p < 0.001 than BL) on days 14 and 21, respectively.
Paired associates learning
There was no significant change in total number of errors at HA than at sea level in Kyrgyz soldiers, but total number of errors were significantly higher (p < 0.01) in Kyrgyz soldiers than in Indian soldiers at sea level, after 3 days at HA exposure and after deinduction. In Indian soldiers, total number of errors were significantly higher (p < 0.01) after 7 days of exposure than at sea level, and after that it was decreasing and close to baseline value after 21 days (Fig. 9A). No significant changes were observed in total trial (Fig. 9B) and memory scores (Fig. 9C) in both the groups at HA.
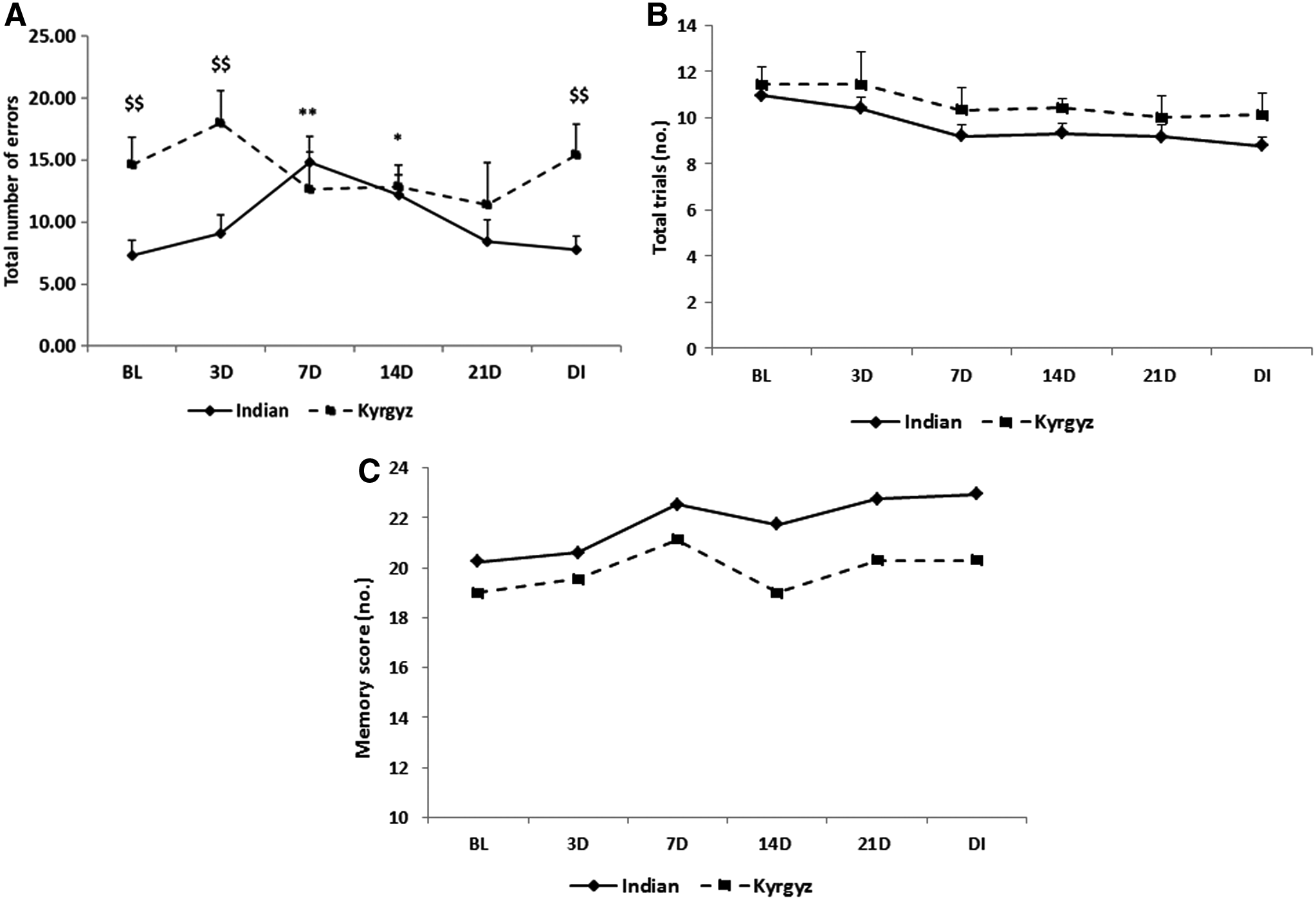
Effects of high altitude on paired associate learning. Line graph represents total number of errors
Pattern recognition memory
Percentage correct of pattern recognition memory was significantly decreased (p < 0.01) on day 3 of exposure in Kyrgyz soldiers and it was gradually increased from 7th day and close to sea level value after deinduction. In case of Indian soldiers, percentage correct was significantly decreased on 7th day and it was further decreased significantly (p < 0.01) on 14th day, but the value was close to sea level on the 21st day. When we compared between the two groups, the percentage correct was significantly decreased (p < 0.01) on 3rd day and after deinduction in Kyrgyz soldiers than Indian soldiers (Fig. 10).
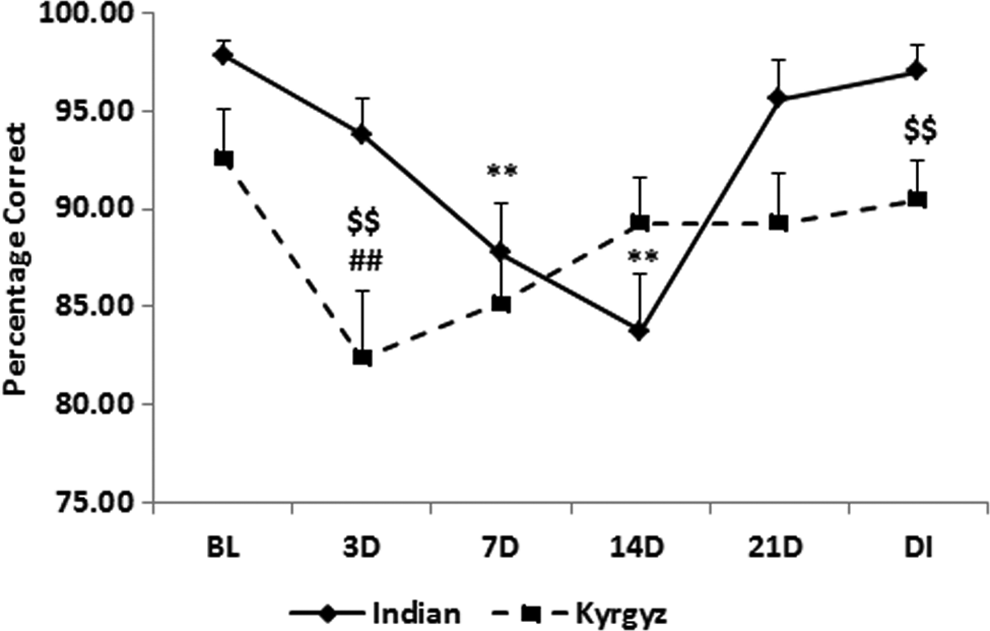
Effect of high altitude on pattern recognition memory. Line graph represents percentage correct of pattern recognition memory at sea level baseline (BL), on day 3 (3D), day 7 (7D), day 14 (14D), day 21 (21D) of high-altitude exposure, and on day 3 of deinduction (DI). Values are mean ± SEM. **p < 0.01 as compared with Indian baseline; $$p < 0.01 as compared with respective Indian groups; ##p < 0.01 as compared with Kyrgyz baseline.
Spatial span
The total errors of SSP increased from day 3 of exposure but were significantly higher (p < 0.01) on 14th day in both the groups. In Indian soldiers, it comes to near baseline value after 21st day and after deinduction, but in Kyrgyz soldiers, total errors were still significantly higher (p < 0.05) after deinduction than at baseline (Fig. 11).
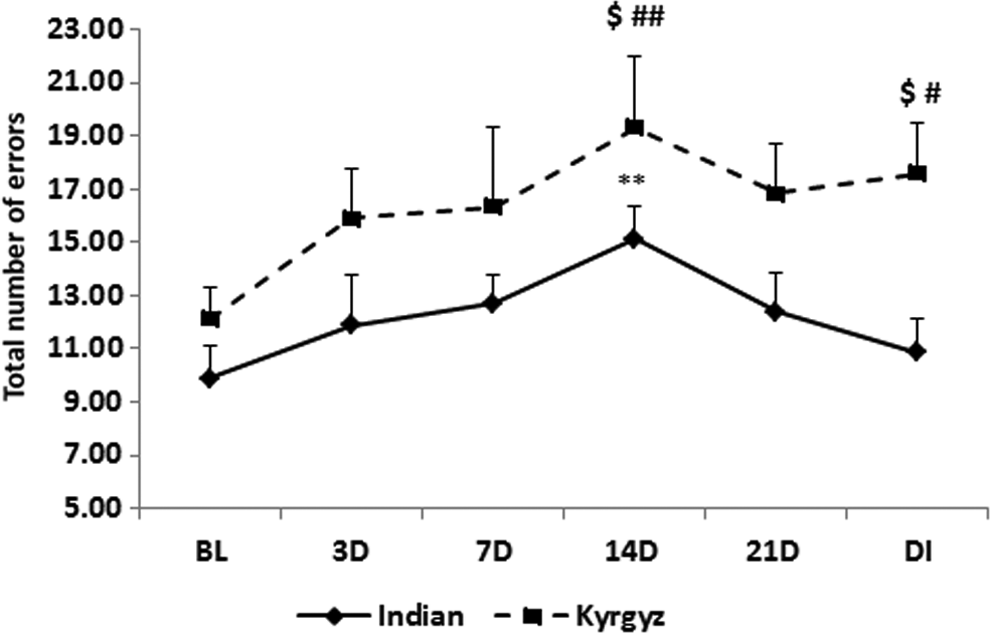
Effect of high altitude on spatial span. Line graph represents total number of errors of spatial span at sea level baseline (BL), on day 3 (3D), day 7 (7D), day 14 (14D), day 21 (21D) of high-altitude exposure, and on day 3 of deinduction (DI). Values are mean ± SEM. **p < 0.01 as compared with Indian baseline; $p < 0.05 as compared with respective Indian groups; #p < 0.05, ##p < 0.01 as compared with Kyrgyz baseline.
Spatial working memory
In Kyrgyz soldiers, between errors of SWM were significantly increased (p < 0.05) on 7th and 14th day and further increased (p < 0.01) on 21st day of stay than baseline. After deinduction, it decreased but was significantly higher (p < 0.05) than baseline. When compared with Indian soldiers, it was significantly higher (p < 0.05) on 21st day of exposure and after deinduction. In Indian soldiers, between errors were significantly increased from day 3 (p < 0.05) and further increased (p < 0.001) on 7th day than baseline, but decreased from day 14 and returned near to baseline after deinduction (Fig. 12).
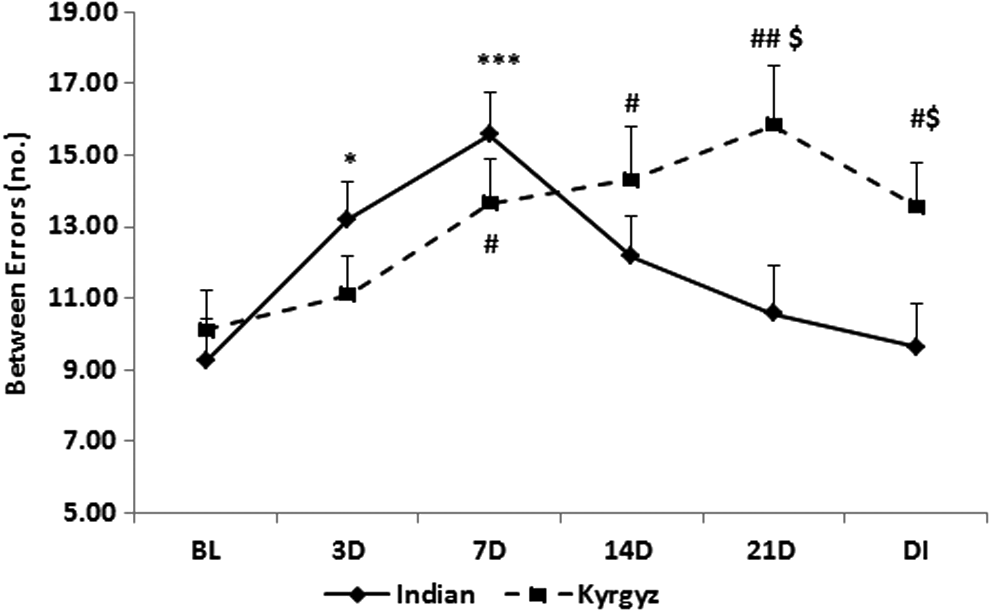
Effect of high altitude on spatial working memory. Line graph represents between errors of spatial working memory at sea level baseline (BL), on day 3 (3D), day 7 (7D), day 14 (14D), day 21 (21D) of high-altitude exposure, and on day 3 of deinduction (DI). Values are mean ± SEM. *p < 0.05, ***p < 0.001 as compared with Indian baseline; $p < 0.05 as compared with respective Indian groups; #p < 0.05, ##p < 0.01 as compared with Kyrgyz baseline.
CRT and SRT
Correct latency of CRT (Fig. 13) and SRT (Fig. 14) in Kyrgyz soldiers was significantly increased (p < 0.05) on 14th day. No further increment was observed on 21st day, but it was significantly higher in both SRT (p < 0.01) and CRT (p < 0.05) after deinduction than baseline. In Indian soldiers, latency of both reaction times was significantly higher (p < 0.01) on 7th and 14th day (p < 0.05), but decreased from 21st day and close to baseline value after deinduction. Both SRT (p < 0.01) and CRT (p < 0.05) were significantly higher in Kyrgyz soldiers than in Indian soldiers after deinduction.
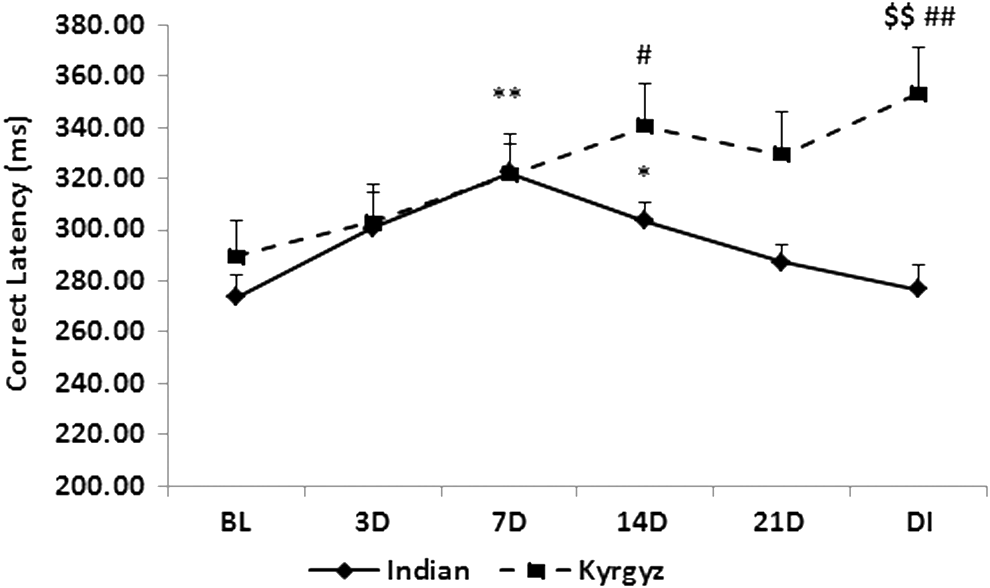
Effect of high altitude on choice reaction time. Line graph represents correct latency of choice reaction time at sea level baseline (BL), on day 3 (3D), day 7 (7D), day 14 (14D), day 21 (21D) of high-altitude exposure, and on day 3 of deinduction (DI). Values are mean ± SEM. *p < 0.05, **p < 0.01 as compared with Indian baseline; $$p < 0.01 as compared with respective Indian groups; #p < 0.05, ##p < 0.01 as compared with Kyrgyz baseline.

Effect of high altitude on simple reaction time. Line graph represents correct latency of simple reaction time at sea level baseline (BL), on day 3 (3D), day 7 (7D), day 14 (14D), day 21 (21D) of high-altitude exposure, and on day 3 of deinduction (DI). Values are mean ± SEM. *p < 0.05, **p < 0.01 as compared with Indian baseline; $p < 0.05 as compared with respective Indian groups; #p < 0.05 as compared with Kyrgyz baseline.
Discussion
The findings of this study support the conclusion that there is a marked difference in acclimatization pattern in cognitive performance between Kyrgyz and Indian soldiers. This difference is consistent throughout the HA stay. Indian soldiers are mostly acclimatized after 7 days of stay or 14 days, but Kyrgyz soldiers are acclimatized after 21 days. Deterioration of cognitive performance is maintained in Kyrgyz soldiers even after returning back to sea level. The poorer performance before acclimatization can be attributed neither to fatigue, since all individuals were tested on 3rd day after their arrival at Syok Pass, nor to mountain sickness, since none of the subjects suffered from such symptoms throughout the whole study. A retest effect could not be entertained to account within group because in some tests Kyrgyz soldiers had higher value after deinduction than baseline.
Results of Indian soldiers are in agreement with Crowley et al. (1992) who described an initial decline followed by a progressive improvement in cognitive performance during prolonged exposure to acute hypobaric hypoxia in a decompression chamber in American soldiers. However, results of Kyrgyz soldiers are similar with the findings of Cavaletti and Tredici (1993), wherein memory, reaction time, and concentration were less efficient when Italian climbers were evaluated 75 days after their return to sea level. Hence, our study suggests that disparity in cognitive ability between Kyrgyz and Indian soldiers at HA may be due to ethnicity and the effects of hypoxia on the brain and, in particular, on some of its structures, whereas the role played by the hippocampus, cortex, and striatum in learning and memory is well known (Titus et al., 2007).
Maiti et al. (2008) reported that DNA fragmentation and apoptotic cell death in hippocampus, cortex, and striatum may be involved in HA-induced memory impairment, and after 7 days of exposure, the effect was more pronounced, but after 21 days of exposure, recovery was observed. Exposure to hypobaric hypoxia equivalent to an altitude of 7600 m for 14 days leads to oxidative stress, chromatin condensation, and neuronal degeneration in the hippocampus (Hota et al., 2008). Increased ionotropic receptor stimulation by glutamate under hypobaric hypoxic conditions could lead to calcium-mediated excitotoxic cell death, resulting in impaired cognitive functions (Hota et al., 2008). HA hypoxia could cause increased cellular oxidative stress (Baily and Davies, 2001). This oxidative stress may be one of the causative factors in HA-induced memory deficit (Maiti et al., 2006), and nitric oxide system is involved in hypobaric hypoxia-induced oxidative stress in rat brain (Maiti et al., 2010). Cholinergic dysfunction is also one of the mechanisms involved in hypobaric hypoxia-induced memory impairment (Muthuraju et al., 2011).
The gene EGLN1, from the hypoxia-inducible factor (HIF) pathway, was identified as being involved in the hypoxic adaptation of Andeans and Tibetans (Ji et al., 2012), as both the groups have adapted to an extremely hypoxic habitat. Furthermore, the identification of the EGLN1 gene from the HIF pathway suggests a common adaptive mechanism for Eurasian populations residing at different altitudes with different oxygen pressures (Ji et al., 2012). Suzuki et al. (2003) observed that there is a genetic variation in the HIF-1alpha gene in Sherpas as compared with Japanese and found a novel dinucleotide repeat polymorphism in intron 13 of the HIF-1alpha gene. Interleukin-6 promoter polymorphism plays a role in morphometric changes of genetic variation in hippocampal shape in mild cognitive impairment (Shen et al., 2004). From these studies, it is clear that ethnic variation plays a pivotal role in HA adaptation. Analysis of mitochondrial DNA and Y-Chromosome short-tandem-repeat polymorphisms reveals a marked genetic diversity in Kyrgyz and Indian populations (Kivisild et al., 1999; Chaix et al., 2004; Quintana-Murci et al., 2004; Heyer et al., 2009).
Thus, it may be concluded that ethnogenetic variation between these two groups might be the cause for their dissimilar pattern of cognitive functioning at HA. Furthermore, to identify specific genes responsible for these ethnogenetic variations, it is important to conduct genetic screening studies.
Footnotes
Acknowledgments
This study was supported by Defence Research and Development Organization (DRDO), Ministry of Defence, Govt. of India. The authors thank the office of Director General Armed Force Medical Services for providing Indian army soldiers for the study. The authors also express their sincere thanks to Ministry of Defence of the Kyrgyz Republic for providing Kyrgyz soldiers for the study.
Author Disclosure Statement
No competing financial interests exist.