
Abstract
Abstract
Background:
Exposure to hypobaric hypoxia (HH) has been reported to cause neurodegeneration and memory impairment. Hippophae rhamnoides, Prunus armeniaca, and Rhodiola imbricata, the indigenous plants of Indian Trans-Himalaya are widely used in traditional Tibetan and Amchi system of medicine. These are rich sources of diverse bioactive metabolites having prophylactic and therapeutic uses against a wide array of neurodegenerative diseases. The objective of this study was to elucidate the prophylactic and neuroprotective efficacy of formulated phytococktail (PC) against simulated HH-induced neurodegeneration in male Sprague Dawley (SD) rats.
Materials and Methods:
A PC containing H. rhamnoides fruit pulp, P. armeniaca fruit pulp, and R. imbricata dry root extract (100:50:1) was formulated. The neuroprotective efficacy of PC was evaluated in male SD rats following exposure to 7 day HH at simulated altitude (25,000 ft, 282 mm Hg). Rats were divided into four groups viz., normoxia group (NOR), normoxic group treated with PC (NORPC), 7 day hypoxic group treated with vehicle (7DH), and 7 day hypoxic group treated with PC (7DHPC). Memory impairment and neuromorphological alterations were measured. Targeted protein expression was analyzed by immunoblotting study.
Results:
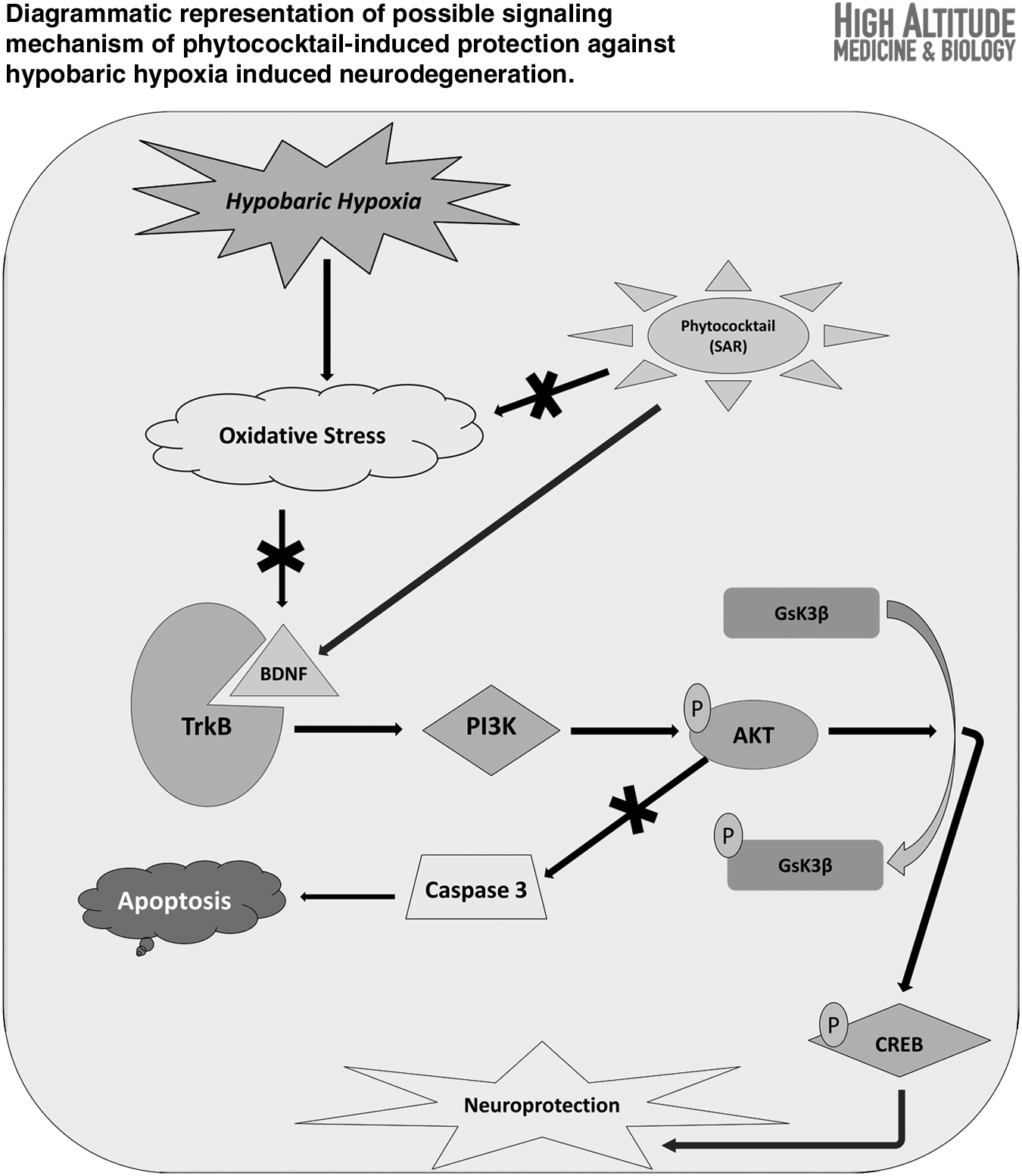
PC supplementation significantly reduced the oxidative stress markers during exposure to HH. Spatial memory impairment by HH was significantly ameliorated by PC. HH-induced augmented pyknosis, decreased dendritic arborization, and increased Hoechst-positive neurons in hippocampal CA3 region were significantly ameliorated by PC. Immunoblotting study showed upregulation of BDNF and TrkB expression by PC. PC also prevented the hippocampal neurodegeneration by activating the PI3K/AKT signaling pathway, which leads to GSK-3β inactivation by its phosphorylation and alleviation of hippocampal Caspase3 expression leading to inhibition of apoptotic neuronal cell death.
Conclusion:
The present study advocates the potential role of PC as an effective neuroprotective supplement in preventing HH-induced neurodegeneration. Activation of the PI3K/Akt pathway through BDNF/TrkB interaction following PC supplementation after exposure to HH inhibits hippocampal neuronal apoptosis and memory impairment.
Graphical Abstract
Introduction
Hypobaric hypoxia (HH) in high altitude or in simulated conditions has been reported to be associated with memory impairment and mood alteration (Baitharu et al., 2013; Barhwal et al., 2015). Furthermore, the altered brain functions in high altitude may partially be attributed to free-radical-mediated oxidative stress (Bakonyi and Radak, 2004; Maiti et al., 2006). Morphological alterations in the hippocampus in HH have also been reported (Shukitt-Hale et al., 1996). As HH is a persisting natural condition at high altitude, a study needs to be conducted to establish possible prophylactic and therapeutic strategies. Cold, arid, and high-altitude regions of the Indian Trans-Himalaya have been shown to induce maladaptation in both acclimatized and nonacclimatized lowlanders (Singh and Selvamurthy, 1993; Schoene, 1996; Dhar et al., 2014, 2018; Palmer and Clegg, 2014).
However, the diverse floral and faunal wealth of the Himalayas also has the underlying remedy to combat these problems. The Himalayan plants are widely used in the traditional system of medicine, both as prophylactics and therapeutics for high-altitude maladies (Singh and Chaurasia, 2000; Ballabh et al., 2008; Dhar et al., 2013a). The indigenous plants of the Trans-Himalaya, such as sea buckthorn (Hippophae rhamnoides L., Elaeagnaceae), apricot (Prunus armeniaca L., Rosaceae), and roseroot (Rhodiola imbricata Edgew., Crassulaceae), are widely used in traditional Tibetan medicine and as nutritional additives.
These are rich sources of diverse bioactive metabolites having prophylactic and therapeutic uses, widely used in the traditional system of medicine (Amchi system of medicine) for treatment of common ailments and have significant health-promoting effects against a wide array of neurodegenerative diseases (Chaurasia et al., 2003; Stobdan and Singh, 2009). Therefore, we aimed to formulate an adaptogenic and neuroprotective botanical supplement called phytococktail (PC) with these three plants to ameliorate HH-induced maladies in animal models.
The PC was found to possess important nutritional attributes, high antioxidant capacities, and diverse bioactive metabolites having promising biological properties (Dhar et al., 2012, 2013a, 2013b), depicted in Supplementary Figures S1 and S2. In the present study, we aimed at evaluating the neuroprotective potential of the PC in male Sprague Dawley (SD) rats following exposure to simulated HH. We also evaluated the toxicological safety profile of PC according to the guidelines of the organization for economic cooperation and development (OECD).
Materials and Methods
Collections of plant materials
Sea buckthorn (Hippophae rhamnoides L.), apricot (Prunus armeniaca L.), and roseroot (Rhodiola imbricata Edgew.) were collected from Choglamsar, Leh [3500 m above mean sea level (msl), latitude 34°6′38.9664″N, longitude 77°35′10.3992″E], in September, apricot field gene bank of Defense Institute of High Altitude Research, Leh (3500 m msl, latitude 34°8′16.119″N, longitude 77°34′19.2216″E) and Chang-La, Changthang (5330 m msl, latitude 34°2′49.812″N, longitude 77°55′49.7778″E), respectively, in the month of September to October 2010. The voucher specimens of H. rhamnoides (HR 6–8), P. armeniaca (PR 4–6), and R. imbricata (RI 5–7) have been deposited at the institutional herbarium for future reference.
Preparation of the PC
The PC was prepared through our previously described process (Dhar et al., 2012). Briefly, the powdered materials of H. rhamnoides fruit pulp, P. armeniaca fruit pulp, and R. imbricata dry root extract were taken in the ratio of 100:50:1 (mg/mL), mixed properly to obtain a homogenous mixture of the PC. We optimized the ratio in the PC (sea buckthorn:apricot:roseroot) based on previous reports on single plants. Different combinations were tested in vitro for cell viability studies, where the 100:50:1 ratio showed optimum results.
Dose optimization and toxicological safety evaluation
Experimental design and animals
All experimental procedures and protocols for animal studies were approved by the Ethics Committee of Defense Institute of Physiology and Allied Sciences (DIPAS), Defense Research and Development Organization, New Delhi (IAEC No. IAEC/DIPAS/01/08/NB dated January 29, 2008) in accordance with the guidelines of the Committee for the Purpose of Control and Supervision of Experiments on Animals, Government of India. Care was taken to minimize the suffering of the animals during the experiment. Male SD rats weighing 135–145 g were housed in polypropylene cages (3 rats per cage) in an air-conditioned room at 26°C ± 2°C and 60% ± 5% relative humidity with a 12-h light/12-h dark cycle. Rats were provided with water and standard food pellets ad libitum (Lipton India Ltd., Mumbai, India).
The toxicological safety profile of the PC was carried out according to the OECD guidelines. Male SD rats (n = 24) were divided into four groups (n = 6/group): (1) Group A: control; (2) Group B: PC 250 mg/kg body weight (BW); (3) Group C: PC 500 mg/kg BW; (4) Group D: PC 1000 mg/kg BW.
For dose optimization in relation to food intake, male SD rats (n = 48) were divided into eight groups (n = 6/group) and treated for 2 weeks with the following doses of the PC: (1) control, (2) 25 mg/kg BW, (3) 50 mg/kg BW, (4) 100 mg/kg BW, (5) 150 mg/kg BW, (6) 200 mg/kg BW, (7) 250 mg/kg BW, and (8) 300 mg/kg BW. The optimum dose, which resulted in highest food intake, was selected.
Pharmacological administration of PC and observations
The fine PC powder was dissolved in 1% saline water and administered orally at a constant volume of 1 mL to animals by the oral gavage method once daily for 28 days. Toxicological evaluation of PC was performed according to OECD test guidelines (OECD, 1995, 1998). Mean BW gains were calculated for each group at each interval and for the overall testing intermission. Animals were also weighed immediately before sacrifice (fasting BW) for calculation of organ-to-BW ratios.
Blood samples for biochemical and hematological analysis were collected through orbital sinus bleeding after anesthetizing the rats with sodium pentobarbital (35 mg/kg BW, intraperitoneal). Examination of hematological and serum biochemical parameters were carried out through the Mindray BC-2800VET Auto Hematology Analyzer (Shenzhen Mindray Bio-medical Electronics Co. Ltd., Nanshan, China) and Cobas Fara Clinical Chemistry Analyzer (Roche Diagnostics GmbH, Mannheim, Germany) according to the instructions provided by the manufacturer. Gross necropsy and histopathological examination were also performed.
Evaluation of neuroprotective and adaptogenic potential
Experimental design
Male SD rats weighing 220–230 g (3 months old) were divided into four groups (n = 12/group), such as normoxia group (NOR), normoxic group treated with PC (NORPC), 7 day hypoxic group treated with vehicle (7DH), and 7 day hypoxic group treated with PC (7DHPC). Oral administration of PC (250 mg/kg BW) was continued until the end of hypoxic exposure.
Exposure to simulated HH
Animals to be exposed to HH were inducted into a simulated altitude of 7600 m (25,000 ft, 282 mm Hg) in a specially designed animal decompression chamber where the altitude could be maintained by reducing the ambient barometric pressure (Barhwal et al., 2015). The temperature and humidity in the chamber were maintained precisely at 27°C ± 2°C and 63% ± 2%, respectively. The HH exposure was continuous for the stipulated period, except for a 15- to 20-minute interval each day for the replenishment of food and water and changing of cage housing materials. The experimental design for the study denoting time period of memory test, probe trial, simulated hypoxic exposure, and PC intervention has been shown in Figure 1.
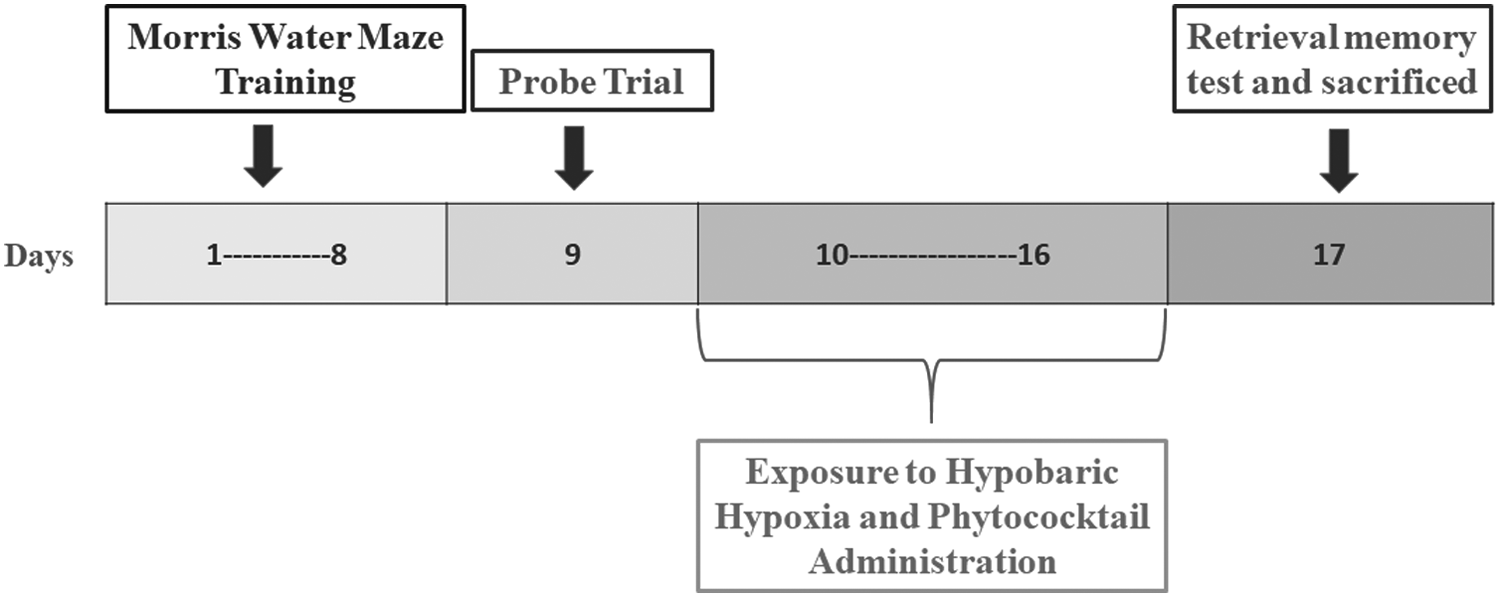
The experimental design for the study representing time period of memory test, probe trial, simulated hypoxic exposure, and PC administration. PC, phytococktail.
Preparation of drug and pharmacological administration
The PC powder was dissolved in 1% saline water. The drug was fed to the rat orally once daily using the feeding cannula at a dose of 250 mg/kg BW during the days of exposure to HH. PC was administered once daily to the requisite experimental groups.
BW and food intake
Each animal was weighed once every day during the study. Food and water consumptions were measured weekly per cage and mean food and water consumptions for individual rats were calculated.
Exercise protocol of treadmill test
To evaluate the endurance performance, the rats were introduced to a treadmill (DB030l Device; Zhishuduobao Biological Technology Co., Beijing, China) running with 15- to 25-minute exercise at 12–20 m/min for 1 week to familiarize them to running. On the day of the exercise test, rats were required to run to exhaustion on the treadmill at a ramp speed adjusted to 22 m/min, 45° ramp inclination with a current repetition rate of 7 ms and current intensity of 9 mA (Brooks and White, 1978; Liu et al., 2005).
Oxidative stress markers and antioxidant status
On completion of hypoxic exposure, rats were sacrificed and the blood samples were collected in the heparin-coated blood collection vial. Blood plasma was isolated by centrifugation at 2000 rpm at 4°C and was taken for reactive oxygen species (ROS) estimation, lipid peroxidation, and reduced glutathione (GSH) estimation. The total protein content per 10 μL of each of the samples was estimated using bovine serum albumin (BSA) as the standard (Bradford, 1976).
Estimation of ROS
ROS, mainly hydrogen peroxide and peroxynitrite, in the blood plasma were estimated spectrofluorimetrically using 2,7-dichlorofluorescein-diacetate (DCFHDA) as described in earlier reports (LeBel et al., 1990; Myhre et al., 2003). In brief, 1.494 mL of 0.1 M phosphate-buffered saline (pH 7.4) was added to 25 mL of the crude homogenate, followed by addition of 6 mL of DCFHDA. The sample was then incubated for 15 minutes at 37°C in the dark and readings were taken at 488 nm excitation and 525 nm emission. The readings were expressed as fluorescent units/mg protein and converted into a percentage by taking the normoxic value as 100%.
Estimation of lipid peroxidation
Lipid peroxidation was measured through the thiobarbituric acid test for malondialdehyde following the method described previously (Das and Ratty, 1987; Colado et al., 1997). Blood plasma was deproteinized with 40% trichloroacetic acid and 5 M hydrochloric acid, followed by addition of 2% (w/v) thiobarbituric acid in 0.5 M sodium hydroxide. The reaction mixture was heated in a water bath at 90°C for 15 minutes and centrifuged at 12,000 g for 10 minutes. The pink chromogen formed was measured at 532 nm spectrophotometrically and expressed in mmol/mg protein. The results were then converted into a percentage considering the normoxic value as 100%.
Measurement of GSH level
The GSH in plasma was measured according to the protocol described previously (Hissin and Hilf, 1976). The amount of GSH was calculated using a standard curve, expressed in mmol/mg of protein and converted into a percentage by taking the normoxic value as 100%.
Adenosine triphosphate level in muscle tissue
The adenosine triphosphate (ATP) content in muscle tissue was determined by using an ATP Chemiluminescence Assay Kit (Calbiochem, San Diego, CA) according to the manufacturer's instructions. In brief, the tissue homogenate was treated with nuclear-releasing buffer for 5 minutes at room temperature with gentle shaking. To the lysate, ATP-monitoring enzyme was added and the luminescent reaction was immediately analyzed in a microplate reader (Spectramax M5e; Molecular Devices, Germany).
The absolute ATP content was calculated by running an ATP standard curve with known ATP concentrations. Protein concentration of samples were determined through the Bradford assay. The calculated total ATP concentration was expressed in nanomolar ATP/mg protein and converted into percentages by taking the normoxic value as 100%.
Lactate dehydrogenase activity in muscle tissue
The lactate dehydrogenase (LDH) (EC 1.1.1.2) activity in muscle tissue was estimated by using the LDH Assay Kit from RANDOX (RANDOX Laboratory Ltd., UK) according to the procedure suggested by the manufacturer, with minor modifications. The decrease in absorbance at 340 nm due to the utilization of nicotinamide adenine dinucleotide (NADH) is directly proportional to the enzyme activity. The assay was started by adding 10 μL of sample to 250 μL of the reconstituted reagent containing pyruvate and NADH in phosphate buffer. Phosphate buffer was added in place of the sample as blank.
The absorbance was then taken at 340 nm in an enzyme-linked immunosorbent assay plate reader (SpectraMax M5e; Molecular Devices) after 30 seconds, which was considered 0 minute, and then after 1, 2, and 3 minutes from the initial reading. The temperature was maintained at 25°C during the experiment. The enzyme activity was calculated from the equation: U/1 = 4127 × ΔA 340 nm/min. The results obtained were then expressed in U/mg of protein. The result thus obtained was expressed in percentage taking the mean control value as 100%.
Memory assessment by Morris Water Maze test following exposure to simulated HH
Morris Water Maze (MWM) test was conducted to examine the spatial reference memory of rodents (Morris, 1984). The video recording was performed through an overhead camera and computer-assisted tracking system by using ANY-maze software (Stoelting Co., USA). After completion of exposure (16th day), all four groups of rats were immediately tested for memory performance in MWM. The latency, path length, time spent in the target quadrant, and number of crossings were recorded.
Assessment of morphological alterations in the CA3 region by cresyl violet staining
The Cresyl Violet (CV) staining was performed by obtaining serial frozen sections of 15 μm thickness from cryoprotected brains (n = 6 per group) and sections corresponding to bregma −4.16 mm were used for CV staining (Das et al., 2015, 2016). Six serial sections were taken from each brain sample and neurons showing staining pattern characteristic of pyknosis were counted in an area of 0.1 mm2 in six different views of the hippocampal CA3 region in each section using Stereo Investigator software (MBF Bioscience, USA). The results were expressed as percentage with the normoxia values being considered as 100%.
Chromatin condensation study by Hoechst staining
The occurrence of chromatin condensation, which is an indicator of apoptosis, was studied by Hoechst 33258 staining (Hota et al., 2008a). Sections were permeabilized in 0.1% Triton and stained with Hoechst 33258 (0.5 μg/mL) for 30 minutes in the dark condition. The stained sections were visualized by using blue filter (4′,6-diamidino-2-phenylindole) in an Olympus BX-51 fluorescence microscope and the brightly fluorescing cells were scored and expressed as percent of control.
Assessment of dendritic arborization in the hippocampal CA3 region by Golgi-Cox staining
After the stipulated period of HH exposure and PC supplementation, the rats were deeply anesthetized with sodium pentobarbital and sacrificed. The brains were removed and processed by Golgi-Cox stain using the procedure described previously (Das et al., 2015, 2016). The apical dendritic branching of pyramidal neurons of the hippocampus was quantified by using the Neurolucida software and results were expressed as percent of control with the control values being considered as 100%.
Protein expression study by Western blotting
The protein content of the hippocampal tissues was calculated by the method of Bradford by using BSA as the standard. The protein expression study of BDNF, TrkB, AKT, PI3K, GSK-3β, pGSK-3β, pCREB, Caspase3, and beta actin (Abcam, Cambridge, MA) was performed by western blotting (Das et al., 2015). The protein expression was quantified by the densitometric analysis of the membranes in each group by using Quantity 1 software in a gel documentation system (Bio-Rad, Hercules, CA).
Statistical analysis
All the experimental results were expressed as mean ± standard deviation mentioned in the experiment section by using statistical analysis with SPSS 17.0 software (Statistical Program for Social Sciences, SPSS Corporation, Chicago, IL). Analysis of variance in a completely randomized design, Duncan's multiple range test, and Pearson's correlation coefficients were performed to compare the data. Post hoc analysis was performed using Newman–Keuls test, and values with p < 0.05 were considered significant.
Results
Dose optimization and toxicological safety profile
General information from repeated dose in a 28-day oral toxicity study
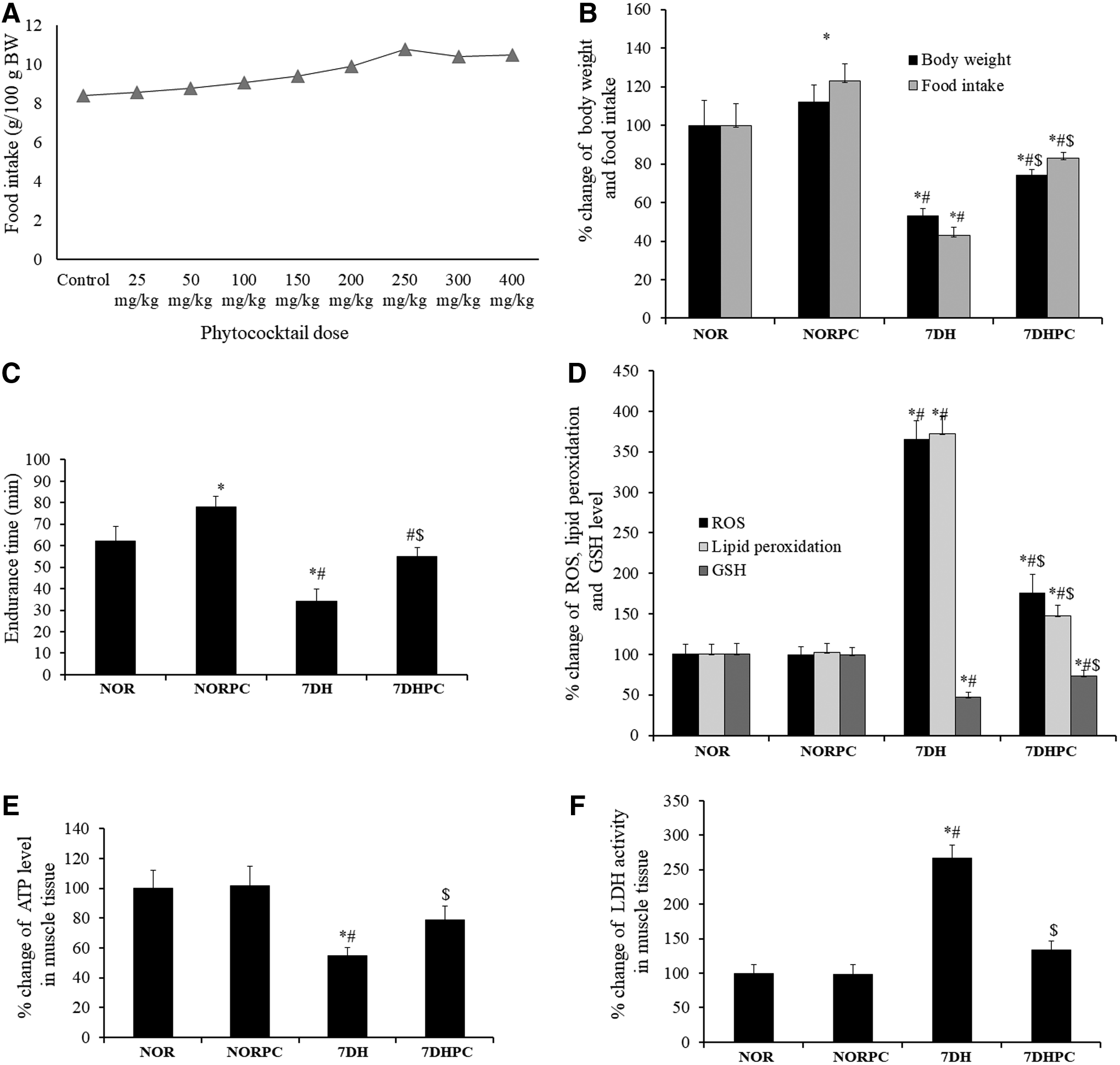
No abnormality was observed over the period of the toxicological study. All animals survived until they were sacrificed at the endpoint of the experiment. The mean BW and mean BW gain of PC-treated rats were increased in comparison with the control group (Table 1). Food and water intake were also increased in the PC-treated groups when compared with the control group (Table 2). The dose of 250 mg/kg BW was found to be the optimum dose in relation to food intake among all treated groups (Fig. 2A).
Body Weight (g) (n = 6/Group)
Values are mean ± standard deviation.
p < 0.05: compared with control.
BW, body weight.
Food and Water Consumption (g/100 g Body Weight) (n = 6/Group)
Values are mean ± standard deviation.
p < 0.05: compared with control.
{}, food intake; (), water intake.
There was no significant difference in hematological parameters among the PC-treated groups compared with the control (Table 3). There was no significant difference observed in biochemical parameters among the study groups (Table 4). Mean absolute and relative (ratio of organ weights to BWs) organ weights for all organs estimated were comparable with the respective control values (Table 5). There was no evidence of pathological abnormality found in rat hearts and livers after PC treatment.
Hematology Parameters (n = 6/Group)
Values are mean ± standard deviation.
BAS, basophile granulocyte; EOS, eosinophil; HCT, hematocrit; HGB, hemoglobin concentration; LYM, lymphocyte; MCH, mean corpuscular hemoglobin; MCHC, mean corpuscular hemoglobin concentration; MCV, mean corpuscular volume; MON, monocyte; PLT, platelet count; RBC, erythrocyte count; RET, reticulocyte count; WBC, total white blood cell.
Serum Biochemical Parameters (n = 6/Group)
Values are mean ± standard deviation.
A/G, serum albumin to globulin ratio; ALB, serum albumin; ALP, serum alkaline phosphatase; ALT, serum alanine aminotransferase; AST, serum aspartate aminotransferase; BUN, serum urea nitrogen; Ca, serum calcium; Cl, serum chloride; CREA, serum creatinine; GLU, serum glucose; HDL-C, serum high-density lipoprotein cholesterol; K, serum potassium; LDL-C, serum low-density lipoprotein cholesterol; Mg, serum magnesium; Na, serum sodium; TBil, total bilirubin; TCHO, serum total cholesterol; TG, serum triglyceride; TP, serum total protein; UA, serum uric acid.
Organ Weights (n = 6/Group)
Values are mean ± standard deviation.
Neuroprotective and adaptogenic potential of the PC following exposure to HH
Gross changes observed during PC supplementation
Food intake in the normoxic group treated with PC increased significantly (p < 0.05) when compared with the normoxic group without PC administration. BW and food intake reduced significantly (p < 0.05) in days of hypoxia exposure with vehicle treatment in comparison with all other study groups (Fig. 2B). The PC-treated groups showed a significantly increased (p < 0.05) endurance time in treadmill running to exhaustion compared with the untreated groups (Fig. 2C).
Changes in ROS, lipid peroxidation, and GSH level in plasma
Plasma ROS and lipid peroxidation level increased significantly (p < 0.05) in the 7-day hypoxic vehicle-treated group in comparison with normoxic and hypoxic groups with PC supplementation. Plasma GSH levels decreased significantly (p < 0.05) in the hypoxic group with vehicle treatment in comparison with all other study groups (Fig. 2D).
Alteration in ATP level and LDH activity in muscle
ATP level in rat muscle tissue decreased significantly (p < 0.05) in 7-day hypoxic vehicle-treated groups treated with the vehicle in comparison with normoxic and hypoxic groups treated with PC (Fig. 2E).
Alteration in LDH activity in muscle
LDH activity in the muscle tissue increased significantly (p < 0.05) in 7-day hypoxic vehicle-treated group when compared with normoxic and hypoxic groups treated with PC (Fig. 2F).
PC supplementation ameliorates HH-induced memory impairment
In MWM test, vehicle-treated hypoxic rats showed a significant increase (p < 0.05) in path length and latency, with decreased number of platform crossings and time spent in the target quadrant compared with the vehicle-treated normoxic group rats. PC-supplemented 7 day hypoxia (7DH) rats showed decreased (p < 0.05) path length and latency along with increased (p < 0.05) number of platform crossings and time spent in the target quadrant when compared with vehicle-treated 7DH rats (Fig. 3A–F).

Assessment of spatial memory through Morris Water Maze following exposure to HH and PC supplementation.
Changes in pyknotic cell counts following exposure to HH and PC supplementation
The number of pyknotic neurons in the CA3 region of the hippocampus increased significantly (p < 0.05) in the 7DH group compared with the normoxic and normoxia-with-PC supplementation groups. The neuronal pyknosis in the 7-day hypoxic group with PC supplementation was significantly lower (p < 0.05) compared with animals subjected to similar duration of exposure to hypoxia (Fig. 4A). No significant reduction in neuronal pyknosis in the CA3 region of the hippocampus was observed between normoxia and normoxia-with-PC groups.
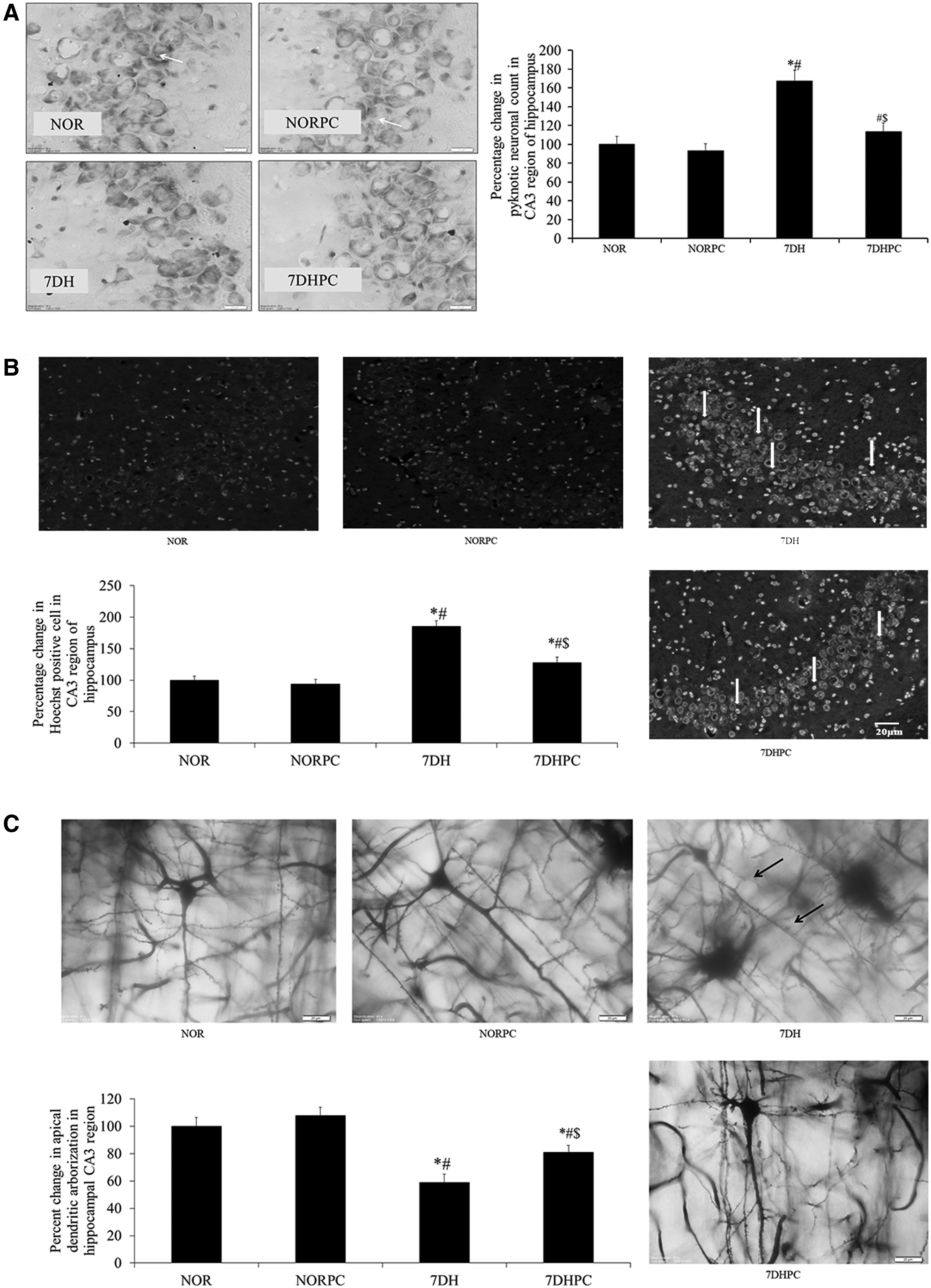
Representative images of Cresyl Violet staining depicting pyknotic cells in the CA3 region of hippocampus
Estimation of chromatin condensation by Hoechst staining
There was a significant increase (p < 0.05) in the number of Hoechst-positive cells in the CA3 region of the hippocampus of the vehicle-treated hypoxic group compared with the normoxic group. Supplementation of the PC during exposure to HH for 7 days resulted in a significant reduction (p < 0.05) in the number of Hoechst-positive cells compared with the vehicle-treated hypoxic group (Fig. 4B).
Assessment of alterations in apical dendritic arborization by Golgi-Cox staining
Exposure to HH for 7 days significantly decreased (p < 0.05) the apical dendritic arborization in the CA3 region of the hippocampus when compared with the normoxic group. However, the administration of PC during exposure to HH significantly increased (p < 0.05) the apical dendritic arborization in the CA3 region compared with the HH group (Fig. 4C).
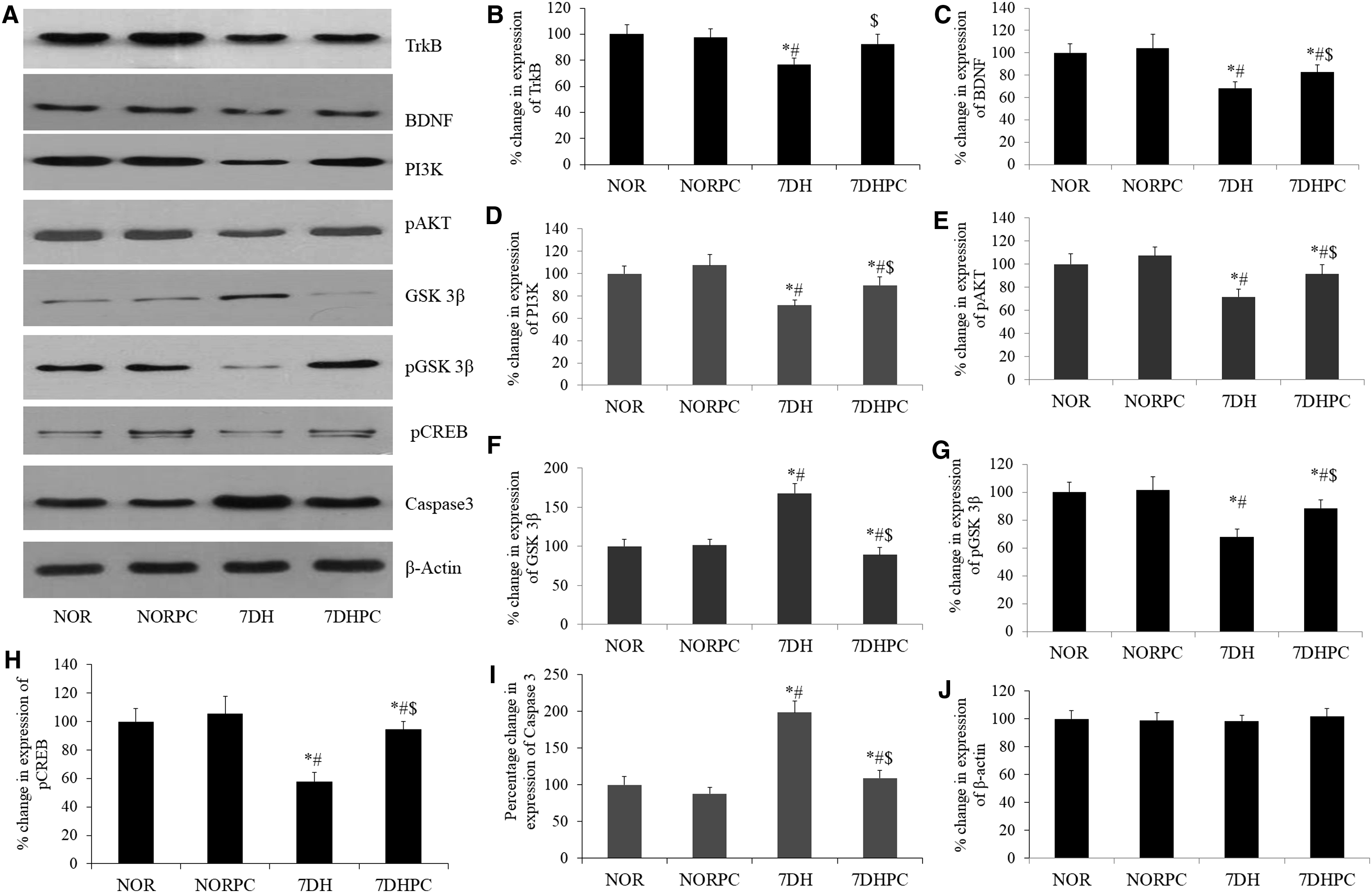
Assessment of protein expression by Western blot
Western blot densitometry analysis showed that PC supplementation significantly upregulated (p < 0.05) the expression of BDNF, TrkB, AKT, PI3K, pGSK-3β, and pCREB in the hippocampal region during exposure to HH for a period of 7 days when compared with the 7DH group. Furthermore, there was a significant reduction (p < 0.05) in the hippocampal expression of GSK-3β and Caspase3 following exposure to HH was also ameliorated by PC supplementation (Fig. 5).
Discussion
HH in high altitude and in simulated conditions is known to cause cognitive and mental dysfunctions along with memory deficits, motor impairment, and hypophagia (Hamilton et al., 1991; Bahrke and Hale, 1993; Singh and Selvamurthy, 1993; Shukitt-Hale et al., 1998). Diverse arrays of physiological and psychological responses are instigated by the unique physical and environmental factors of cold arid high-altitude regions of the world (Dhar et al., 2014; Sharma et al., 2014). The adverse geo-climatic conditions prevailing in the high-altitude region of the Trans-Himalayan cold desert cause a threat to human survival, and there is ample evidence that in this unfavorable hypoxic environment, several pathophysiological events could occur due to the alteration in physiological function.
Our laboratory is also situated in the Trans-Himalayan high-altitude region at 3500 m above msl, where we investigated the effects of high-altitude stay in healthy humans and amelioration of high-altitude maladies by Trans-Himalayan plants. Prolonged high-altitude exposure causes various health ailments, including cognitive impairment and loss of memory. To ameliorate these high-altitude maladies, we planned to develop a novel prophylactic using the indigenous botanical resources of this region.
Among the native Trans-Himalayan plants, sea buckthorn, apricot, and roseroot have long been used in the traditional medicinal system of India, China, and many other countries. Several herbal products have also been developed from our institute and accepted for human consumption as a health supplement (Ballabh et al., 2008; Dhar et al., 2013a, 2013b). Therefore, a formulated PC with potent antioxidant capacity and adaptogenic potential can be useful against HH-induced neurodegeneration.
Hypoxia-induced neurodegeneration and memory impairment is widely reported in experimental animal models (Hota et al., 2008a, 2008b; Barhwal et al., 2015). The hippocampal CA3 region is associated with cognitive aspects, and thus hippocampal neurodegeneration is correlated with cognitive decline (Prasad et al., 2013). Therefore, the present study was conducted on a rat model to elucidate the neuroprotective property of the PC (Fig. 1). The assessment of memory through the MWM test advocated that memory impairment during exposure to chronic HH was significantly ameliorated by PC supplementation in male SD rats. The PC was also found to provide hippocampal neuroprotection against hypoxia-induced neurodegeneration by significant reduction in pyknotic neuronal counts.
The CA3 region of the hippocampus has attracted foremost interest in recent years for its specific role in memory processes, susceptibility to seizures, and neurodegeneration. Furthermore, the CA3 region's structural and functional integrity is required for the memory retrieval process after consolidation of memory (Lee et al., 2004; Leutgeb et al., 2005; Ji and Maren, 2008; Rebola et al., 2017). The augmented Hoechst-positive neurons in the hippocampal CA3 region following exposure to HH for 7 days were significantly downregulated by PC supplementation. Golgi-Cox staining of the CA3 pyramidal neurons showed significant reduction in apical dendritic arborization following exposure to HH. However, PC supplementation showed significant restoration of apical dendritic arborization in 7DH group rats.
The immunoblotting study showed upregulated expression of BDNF, TrkB, AKT, PI3K, pGSK-3β, and pCREB in the rat's hippocampal region after PC supplementation during exposure to hypoxia, which confer significant neuroprotection by the activation of the PI3K/Akt signaling pathway through BDNF/TrkB interaction. The reduced expression of GSK-3β and Caspase3 following exposure to HH was also augmented by PC supplementation.
In addition, PC supplementation also sequestered the CREB phosphorylation in the hippocampal region and conferred neuroprotection through expression of survival genes. Our results were in agreement with previous reports, where sea buckthorn, apricot, and roseroot plants, which were used for developing the PC, showed promising neuroprotective activities through these signaling pathways (Chen et al., 2012; Chiang et al., 2015; Liu et al., 2016; Zhang et al., 2016a, 2016b; Heitman and Ingram, 2017).
In the present study, we also evaluated the adaptogenic potential of the PC in SD rats following exposure to chronic HH. A significant reduction in the BW and food intake was observed in the 7DH group compared with control (normoxia). Our results are in agreement with previous reports on the feeding behavior of rats exposed to chronic HH (Singh and Selvamurthy, 1993; Singh et al., 1997). Previous reports have also advocated the recovery of stress-induced BW loss in rats after sea buckthorn (Hippophae rhamnoides) supplementation (Vijayaraghavan et al., 2006; Diandong et al., 2016; Du et al., 2017). R. imbricata also showed a significant function in BW gain in experimental animals and the adaptogenic potential of this plant has been well established (Gupta et al., 2008; Chiang et al., 2015).
In the line, our study also showed the potential role of the PC supplementation in BW gain, augmented food and water intake in hypoxic rats. The underlying causes for the loss of BW may be attributed to a higher metabolic rate, different energy output, and reduced perception of hunger together with decreased food intake (Jain et al., 2012). The hypophagia and BW loss were recovered with PC supplementation during exposure to HH, which showed its adaptogenic and appetizing potential. Furthermore, our study did not record any significant difference in hematological parameters in hypoxic groups as compared with the control and the findings were in agreement with earlier reports (De Franceschi et al., 1999).
However, a recent study on experimental fish showed that acute hypoxic exposure leads to significant increase in hematological parameters (Chen et al., 2017). The findings of our study showing no significant change in hematological parameters of experimental rats following hypoxia may possibly be due to the experimental animals used for simulated hypoxic exposure.
Previous reports on HH exposure showed increased oxidative stress (Nakanishi et al., 1995; Maiti et al., 2006) and depleted endogenous antioxidant enzyme level in rat tissue as well as in human blood (Sairam et al., 1998). In corroboration with the previous reports, our results also documented an elevated level of free radicals and lipid peroxidation following 7-day exposure to HH along with reduced level of endogenous antioxidants in rat. Administration of PC during exposure to HH was found to reduce the free radical level, which further diminished the incidence of lipid peroxidation. The antioxidant capacities and radical scavenging properties of the PC could be attributed to the rich content of polyphenols, flavonoids, glycosides, phenylpropanoids, vitamins, and other bioactive components with strong antioxidant effect (Dhar et al., 2012, 2013a, 2013b).
The PC-induced augmentation of endogenous GSH level in blood indicated the efficacy of active biomolecules in modulating glutathione biosynthesis and stabilization under hypoxic conditions. However, the underlying mechanisms of such a regulatory effect of PC on endogenous glutathione biosynthesis need further investigation.
We observed that exposure to HH for 7 days depleted the ATP level in the rat's muscle tissue. However, PC supplementation during hypoxic exposure restored the ATP level in the muscle. We also observed significantly increased muscle LDH activity in rats exposed to 7-day hypoxia that was reduced after PC supplementation. An earlier report also advocated that increased LDH activity in rat skeletal muscle is associated with hypoxic exposure (Marti et al., 1994). The improved endurance time in the treadmill test was further confirmed by reduced accumulation of lactate in the rat muscle tissue.
These findings indicate strong neuroprotective and adaptogenic potential of the PC against HH. The neuroprotective effect may be attributed to the synergistic effect of diverse bioactive phytochemicals, such as glycosides, phenylpropanoids, alkaloids, phytosterols, aromatic aldehydes, ascorbic acid, etc. present in the PC (Dhar et al., 2012, 2013a). These compounds possess strong antioxidant and anti-inflammatory properties, which exhibited protective effect on neuronal cells under hypoxia (Dunn et al., 2011; Kumar and Khanum, 2012; Yu et al., 2013; Spagnuolo et al., 2016; Zhou et al., 2018). However, further clinical studies should be conducted to establish the therapeutic potential of the PC that could be used as a dietary supplement by humans exposed to HH conditions persisting at high altitude.
Conclusions
Considering the prevalence of stress-induced pathophysiological conditions in HA, we aimed at developing a PC that could not only prevent the onset of chronic diseases but also improve human performance in HA environments. The findings of the present study demonstrated the neuroprotective potential of the PC against hypoxic damage to organ systems, including the brain, and accelerate the onset of adaptive changes following hypoxic stress in rats. The PC comprising sea buckthorn, apricot, and roseroot, the three indigenous plants of Indian Trans-Himalaya, is rich with a diverse group of bioactive phytochemicals and nutritional attributes that may contribute to the neuroprotective and adaptogenic properties.
However, the in vivo bioavailability and other pharmacokinetic parameters of the PC need to be investigated for further results. Future research toward understanding the signaling cascades of the neuroprotective role of the PC may provide new insights for developing novel prophylactic and therapeutic strategies against HH-induced neurodegeneration.
Footnotes
Acknowledgment
The authors acknowledge the contribution of Dr. Ritendra Mishra, Mumbai, India, who helped in the copyediting and proofreading of the article.
Author Contributions
S.B.S. conceived the project and designed experiments. P.D., S.K.D., and K.B. carried out experiments and acquisition of data. S.K.D. and P.D. wrote the article. S.K.H. and K.P.M. provided crucial inputs for experiments and edited the article. The corresponding author (S.B.S.) has given her consent that all co-authors have reviewed and approved the article before submission.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The study was entirely supported by Defense Research and Development Organization, Ministry of Defense, Government of India.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.