
Abstract
Background:
Both dynamic and static visual performances are essential for safety and motoric performance at altitude. There is a lack of information regarding alterations in dynamic visual performance (DVP) in oxygen-reduced environments. The purpose of this study was to analyze DVP in natural hypoxic conditions in a group of young, healthy hikers.
Methods:
DVP in four parafoveal subfields was analyzed using the computer-assisted Düsseldorf Test for Dynamic Vision. Measurements were performed twice at altitudes above 3500 m during an 8-day alpine hike.
Results:
On day 5 (3647 m), no changes in DVP were detected. On day 6 (4554 m), however, we found a significant reduction in DVP in the superior parafoveal retinal subfield, partly representing the lower visual field. The observed changes did not correlate with oxygen saturation, hematocrit, or cardiovascular parameters. We found no interrelation between symptoms of acute mountain sickness and DVP at altitude.
Conclusions:
Our data suggest that hiking at altitudes above 4500 m results in lower DVP in the visual field of healthy young people. The alteration might affect motor performance and coordination, increasing the risk of accidents.
Introduction
Vision is a key sense for safety and motor performance in sports (Jendrusch et al., 1999, 2010; Senner et al., 1999), and for recreation at high altitude (Ellerton et al., 2009). Alterations to the human eye during exposure to high altitude can include structural, functional, and physiological changes, such as altered intraocular pressure, optic disc swelling and increased optic nerve sheath diameter, an increase of corneal thickness, altered retinal arterial and venous diameter, and retinal blood flow velocity (Cymerman et al., 2000; Morris et al., 2007; Somner et al., 2007; Bosch et al., 2008, 2009, 2010a, 2010b; Sutherland et al., 2008; Willmann et al., 2011, 2014a, 2017; Keyes et al., 2013). Studies have been published on changes in color vision, contrast sensitivity, visual acuity, night vision, and the visual field (Kobrick et al., 1988; Davis et al., 1995; Yap et al., 1995; Leid and Campagne, 2001; Horng et al., 2008; Tekavcic-Pompe and Tekavcic, 2008; Willmann et al., 2010; Davies et al., 2011; Fischer et al., 2012; Gekeler et al., 2019).
Some studies concerning parts of the spectrum of motion perception (efferent oculomotor dynamic performance) have been performed in recent years (Still et al., 2011; Merz et al., 2013; Di Stasi et al., 2014; Faull et al., 2015). Since visual perception at altitude is of major importance, the quality of motion perception is highly relevant. During exercise, information given by the visual system is used to control movement, regulate balance, and provide environmental orientation (Patla, 1997). Visual motion detection occurs in two ways: as afferent result of the perception of distant retinal images without eye movement and in the form of eye-tracking movements as a result of oculomotor efference. In nearly every mountain sport, moving retinal images are a vital source of visual information. Alongside other characteristics of visual perception and secondary sensory modalities, dynamic visual performance (DVP) is essential for safe travel in high-altitude terrain, as it assists with orientation and obstacle avoidance (Reynard and Terrier, 2015).
Studies have provided ambivalent results regarding the association between acute mountain sickness (AMS), self-reported scores, and ophthalmological changes under hypobaric hypoxia (Bosch et al., 2008, 2009, 2010a; Sutherland et al., 2008; Willmann et al., 2011; Fischer et al., 2012). Therefore, the aim of this study was to analyze DVP of healthy young hikers under natural hypoxic high-altitude conditions and to enhance our knowledge of hypoxia's influence on visual perception. To our knowledge, DVP has not yet been investigated under conditions of high-altitude exposure (including terms of oxygen deficiency and AMS).
Materials and Methods
The study was performed in accordance with the Declaration of Helsinki (1983 revision) and was approved by the medical ethics committee of the Ruhr-University Bochum. All subjects gave their written consent.
Subjects
Eleven young healthy trichromatic hikers (two female and nine male, average age 24.0 ± 2.5 years, and body mass index 22.7 ± 1.3 kg·s−2), normally resident at altitudes between 100 and 500 m, participated in the study. All subjects required no or low spectacle correction (refraction error ≤ ±2.5 diopters) and had no previous eye surgery or reported visual impairment. Before hiking, all subjects showed normal to high DVP in relationship to their age (Wist et al., 2000). None of the subjects visited high altitudes in the 6 months before the study. Before beginning the study, the physical fitness and general health of each subject were checked. The latter was checked by a medical examination and by measuring blood concentrations of relevant clinical parameters like electrolytes, liver and kidney values, blood cells, and inflammatory markers. Physical fitness was evaluated by determining the running velocity corresponding to a blood lactate concentration of 4 mmol/L (V4) in an incremental running field test (start velocity: 2.4 m·s−1, stage increment: 0.4 m·s−1, stage duration: 5 minutes) and revealed a moderate aerobic performance (V4: 3.7 ± 0.5 m·s−1).
Time of testing and ascent profile
Vision tests were performed before and during an 8-day high-alpine hike (d1–d8) in the Valais Alps of Switzerland and Italy (Fig. 1). The first 4 days served as an acclimatization period of hiking and sleeping at altitudes between 1610 and 4223 m. Accommodation during the hike was in alpine huts, where the tests were completed. Adequate hydration and energy supplies were given for the general conditions. Participants abstained from caffeine and physical exercise for 1 hour before experiments (Mewes et al., 2008). The tests were completed before starting the hike in Bochum, Germany (baseline, d0: 154 m), and on the fifth day (d5: 3647 m) and the sixth day (d6: 4554 m) of the hike. On day 5, the test was done in the afternoon in the Refugio Gnifetti after spending one night in the hut (3647 m). Testing on day 6 was also performed in the afternoon in Capanna Margherita (4554 m) after sleeping in Refugio Gnifetti the night before, and ascending to Capanna Margherita in the morning. The climb took 5 hours and subjects rested for 2 hours before the test.
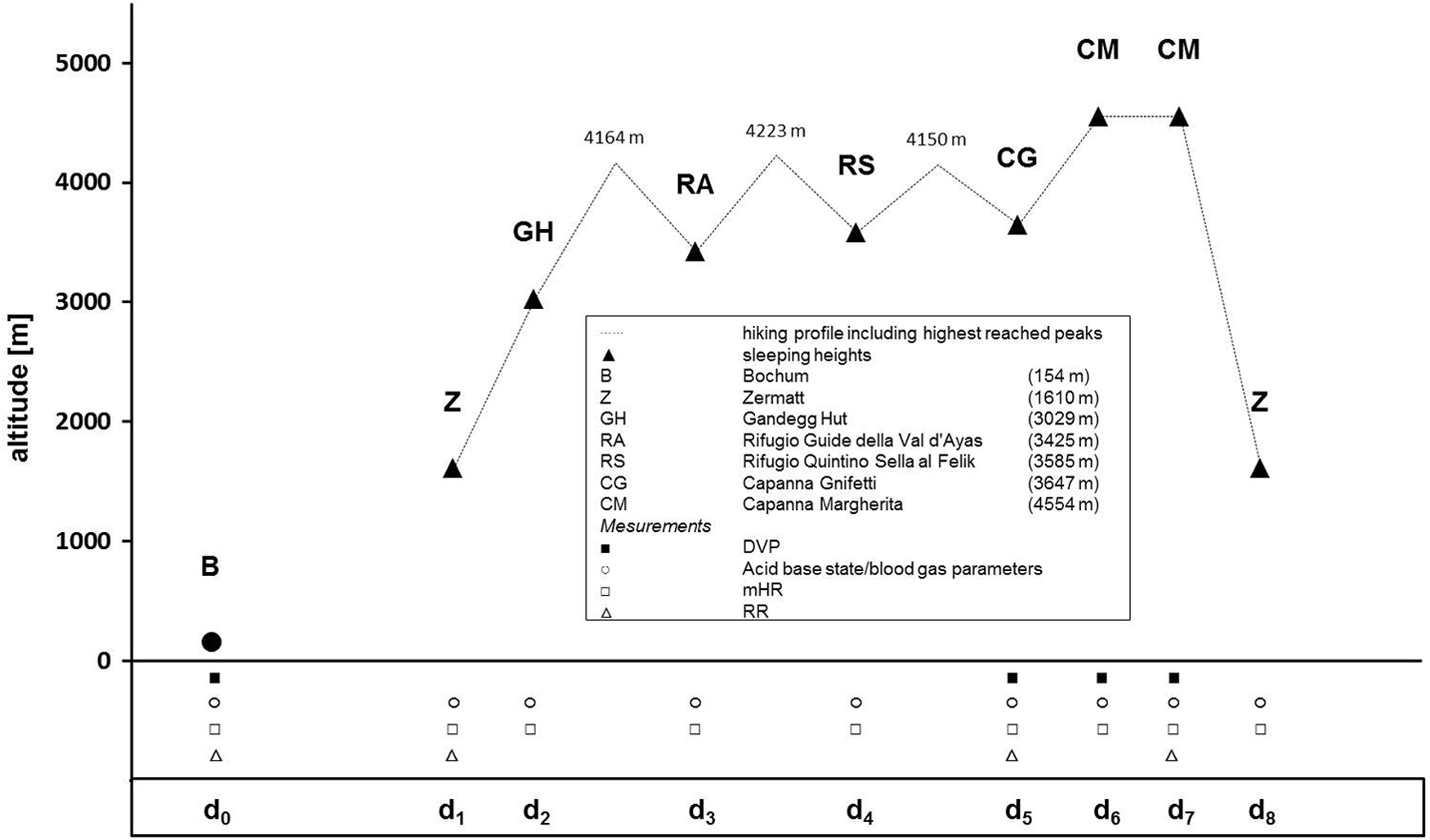
Altitude profile of the high-alpine hike. BL examination (d0) of DVP performed before hiking in Bochum (temperature: 19°C; atmospheric pressure 1019 hPa). Examination of DVP performed during the hike [d1–d8] on day 5 (temperature: 18°C; atmospheric pressure 662 hPa) and day 6 (temperature: 20°C; atmospheric pressure 588 hPa). Legend: d0/d1−d8 = testing/hiking days; ▴ = sleeping heights; ▪ = testing time of DVP; ○ = testing time of acid-base state/blood gas parameters through capillary blood samples; □ = testing time of resting heart rate before getting up in the morning (mHR); ▵ = testing time of blood pressure by Riva-Rocci method (mHR). BL, baseline; DVP, dynamic visual performance.
DVP test and experimental procedure
The computer-assisted Düsseldorf Test for Dynamic Vision (DTDS; Wist et al., 1998; 2000) relies solely on motion cues without differences in luminance, and was used to analyze afferent DVP. The test was done on a laptop with a calibrated high-definition LCD screen (Latitude E6500, 1920 × 1200 pixel spatial resolution; Dell, Inc., Round Rock). To ensure standardized conditions, the testing was performed in a light-impermeable examination tent with low LED luminance (LEDstixx; OSRAM, Munich, Germany) for keyboard orientation (Fig. 2A). The eye-to-screen distance was 59 cm. Each session commenced with a 10-minute adaptation to the lighting conditions of the tent. The optotypes were Landolt rings (constant diameter: 1.8°; Landolt α: 0.36°) with the same average luminance as their surroundings, becoming visible when the dots within the rings were moved briefly, while those in their surroundings remained stationary (motion contrast; Fig. 2B).

(
We used the standard form of the test with 4 different variations of motion contrast (100%, 50%, 30%, or 20%), 20 trails per motion contrast, and 4176, 2088, 1392, and 835 pixels, respectively. Stimulus duration was fixed at 140 ms. The gap of the Landolt rings varied in a randomized order in four different orientations (left, right, top, or bottom). The test started with the highest motion contrast (1.0; 100% pixels moving within the ring) and became more difficult (50%, 30%, and 20% pixels moving within the ring). The DTDS is a forced-choice method without time pressure (Ehrenstein, 2003). Because of the central fixation and short optotype duration, no eye movements can falsify the afferent mode of motion perception. Before each visual test battery, we administered two drops of artificial tears (Blink® Intensive Tears; Abbott, Abbott Park, IL) into the subject's bottom eyelid to prevent altitude-induced “dry eyes” (Gupta et al., 2008; Willmann et al., 2014c). Subjects did not take any medication during the measurements.
Besides testing DVP, we evaluated markers for AMS using the five-item Lake Louise Self-Report (LLS; Roach et al., 1993). LLS was measured daily in the morning, before DVP testing. Prevalence of mild AMS was defined as LLS ≥3, with headaches present. Severe AMS was defined as LLS ≥5 with headaches present (Wagner et al., 2012). Morning resting heart rate (mHR) was measured daily on waking using mobile heart rate monitors (Polar FS2c, Polar Electro Oy, Kempele, Finland). Systolic and diastolic blood pressure (RRs/RRd) was measured in the morning on d0, d1, d5, and d7 using the Riva-Rocci method on the right arm (stethoscope and blood pressure cuff: KaWe, Asperg, Germany; Lewis, 1941; Riva-Rocci, 1896). Capillary blood samples were taken daily on waking from a hyperemic earlobe (Finalgon; Sanofi-Aventis, Paris, France). One sample was taken on day 6 after the hike and before DVP testing. Analysis of blood parameters (epoc® Blood Analysis System; Epocal, Ottawa, Canada) included capillary oxygen partial pressure (pO2), calculated capillary oxygen saturation (cSO2), and hematocrit (Hct). Temperature and atmospheric pressure in the tent were analyzed during DVP tests using the PRO TREK PRW-1300 watch (CASIO, Tokyo, Japan).
Statistical analyses
All statistical analyses were undertaken using SPSS (V. 23.0 for Windows; IBM, Armonk). For all tests, p values less than 0.05 were considered statistically significant. pO2, CSO2, Hct, mHR, and RRs/RRd were calculated as mean ± standard deviation. Effects of high-altitude exposure over time for all variables were evaluated by a two-tailed analysis of variance for repeated measures. Paired student t-tests were performed to further analyze statistical significance between related data over time. Bonferroni adjustment was used to correct the alpha error rate due to multiple testing. Mean LLS scores were analyzed as medians (P50) and the accompanying first (P25) and third quartiles (P75). Effects of high-altitude exposure on LLS over time were evaluated using nonparametric tests (Friedman and Wilcoxon). Alpha adjustment was practiced for multiple testing.
DVP was analyzed three times. The first analysis focused on differences in the overall test results between testing times as the percentage sum of correct answers in DTDS over all test levels (overall DVP). To further analyze specific effects on DVP, the second and third analyses considered variations in motion contrast as the percentage sum of correct answers in DTDS according to motion contrast levels (100%, 50%, 30%, or 20%), and separately for the four different subfields of the parafoveal visual field (left, right, upper, and lower) according to the orientation of the gap in the ring (left, right, top, or bottom).
Because of the small sample size and partly non-normally distributed data, nonparametric evaluations by ranks with repeated measures (Friedman) and comparing related samples (Wilcoxon) were used for statistical analysis. Alpha adjustment was applied for multiple testing. The results are expressed as medians of DVP (P50), and the first (P25) and third quartiles (P75) of correct answers are expressed in DTDS. The effect size for changes in DVP was calculated using the correlation coefficient (r) as described by Rosenthal (1994). To analyze the correlations between pO2, cSO2, mHR, RRs/RRd, LLS, and the isolated LLS item asking the state of fatigue and weakness (LLSFatigue) in relationship to DVP, we used the Spearman's rank correlation coefficient (rs). Explorative analysis using dot plots paired with rs calculations of the interrelation between DVP, V4, and sex was performed.
Results
All participants successfully completed all tests during the hike. No subject showed clinical signs or symptoms of high-altitude pulmonary edema or high-altitude cerebral edema. Only one subject revealed an LLS score of 3 (range = 4–6). On day 2, the mean LLS reached its highest level after one night at an altitude of 3029 m, while incidence of AMS remained 0%. Longer duration at an altitude above 3400 m with ascents above 4000 m (and sleeping levels between 3425 and 3647 m) resulted in an increment of AMS incidence up to 9%–18% between days 3 and 6. Between days 2 and 5, as hikers acclimatized, LLS decreased, before increasing again on day 6 (Friedman's χ2 = 38.84; d.f. = 8; p < 0.01, Table 1).
Oxygen Saturation, Hematocrit, Cardiovascular Parameters, and Lake Louis Self-Report of All Subjects Before and During the Hike

In line with a more or less continuous decline in capillary pO2 from about 85 mmHg (d0) to about 47 mmHg (d6afternoon), cSO2 also decreased during the hike from about 97% (d0) to about 86% mmHg (d6afternoon) with increasing altitude (p < 0.05; pi = α*17, Table 1). At altitudes between 3425 and 3647 m (on days 3 to 6), morning pO2 and cSO2 stabilized around 55 mmHg and around 90%, respectively (p > 0.05; pi = α*17). Hct remained unchanged throughout the hike (d0 = 41.7% ± 2.8%, d1 = 40.6% ± 2.0%, d2 = 41.7% ± 3.2%, d3 = 42.6% ± 2.9%, d4 = 41.1% ± 3.0%, d5 = 40.5% ± 4.7%, d6 = 42.2% ± 3.7%, d6afternoon = 43.6% ± 3.4%, d7 = 42.8% ± 3.3%, and d8 = 41.8% ± 2.5%, respectively; p > 0.05; pi = α*17, Table 1). mHR increased during the first 2 hiking days from 54 to 70 min−1 (p < 0.05; pi = α*15) and then remained slightly elevated during the remaining days (Table 1). Systolic blood pressure remained unchanged (p > 0.05; pi = α*5), while diastolic blood pressure increased from around 11 (d1) to 13 (d7) mmHg (p < 0.05; pi = α*5, Table 1).
DVP of all subfields and different variations of motion contrast remained constant on days 5 and 6 compared to the baseline (p > 0.05; pi = α*2, Fig. 3, Table 2A). No change in overall DVP was found during test conditions of 100%, 50%, 30%, and 20% motion contrast compared to the baseline (p > 0.05; pi = α*2, Fig. 4). Analysis of DVP separated into parafoveal subfields at 20% motion contrast conditions, showing changes for the quality of the lower DVP (Friedman's χ2 = 7.66; d.f. = 2; p < 0.05), but no changes in DVP in the other parafoveal subfields [left (1), right (2), and upper (3)] occurred at both days at altitude (Friedman's χ21 = 1.73, χ22 = 2.00, χ23 = 3.44; d.f. = 2; p > 0.05, Table 2B). However, DVP in the lower subfield tended to decline on day 5 at 3647 m (pi = α*1, z = −1.67, r = 0.36, 2p = 0.096) and was significantly reduced in the lower subfield on day 6 at 4554 m compared to baseline measurements of 154 m (pi = α*3, z = −2.41, r = 0.51, 2p < 0.05, Fig. 5).
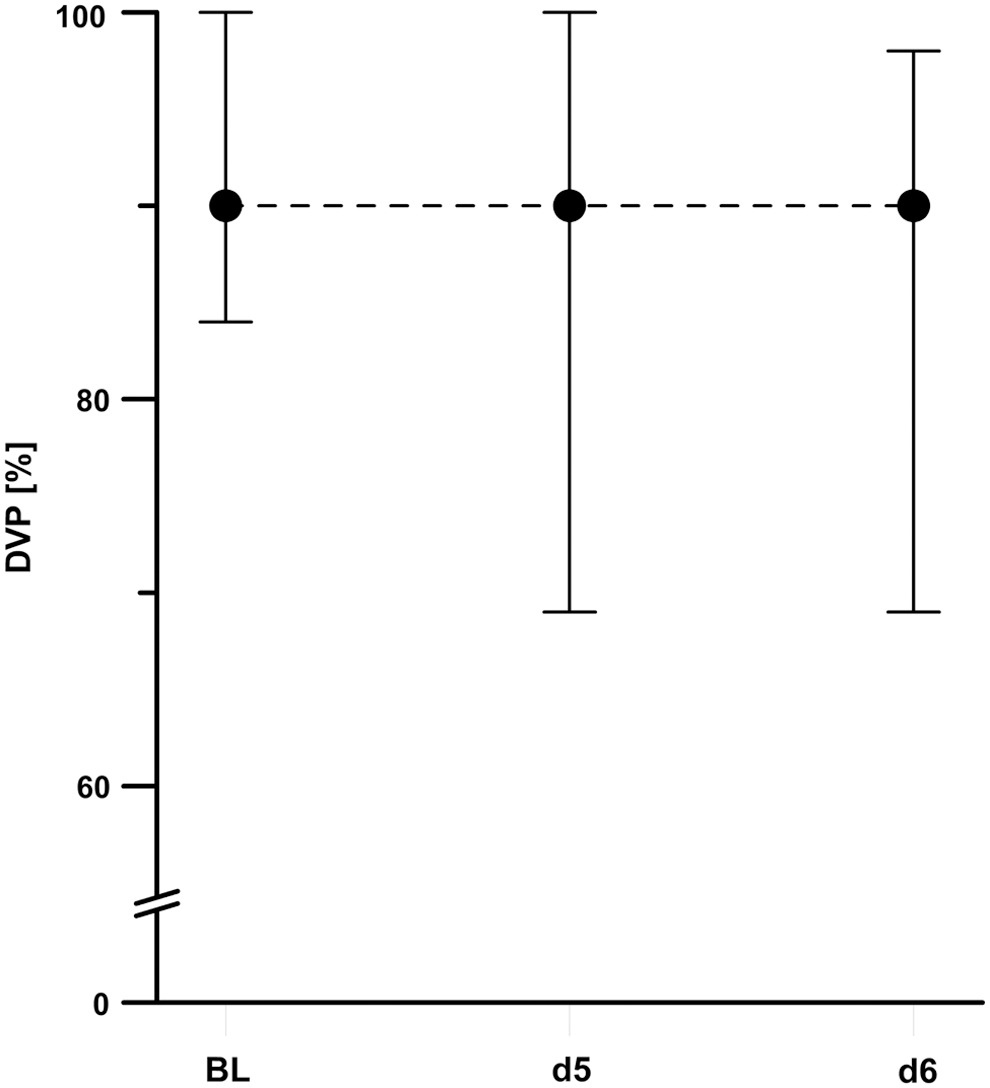
Development of overall DVP on days 5 and 6 compared to BL testing at sea level. The line plots represent the medians (dots) and the interquartile ranges between the first (P25; lower whisker) and third (P75; upper whisker) quartiles (space between the whiskers).

Development of DVP at 100%

Development of DVP at 20% motion contrast condition of all parafoveal subfields of the visual field
Longitudinal Changes of Dynamic Visual Performance
Significant differences in DVP compared to BL (d0): p ≤ 0.05.
We did not find any correlation between DVP and pO2, cSO2, Hct, mHR and RRs/RRd, or LLS, respectively (p > 0.05, Fig. 6). Nor did we find a relationship between endurance performance (1), sex (2), and DVP (rs 1 = 0.02; rs 2 = −0.17; p > 0.05), or presence of AMS (rs 1 = −0.22; rs 2 = 0.21; p > 0.05). As described by Wist et al. (2000), DVP correlates moderately with the age of the subjects (rs = 0.52; p < 0.05), but not with the presence of AMS (rs ≤ 0.00; p > 0.05).
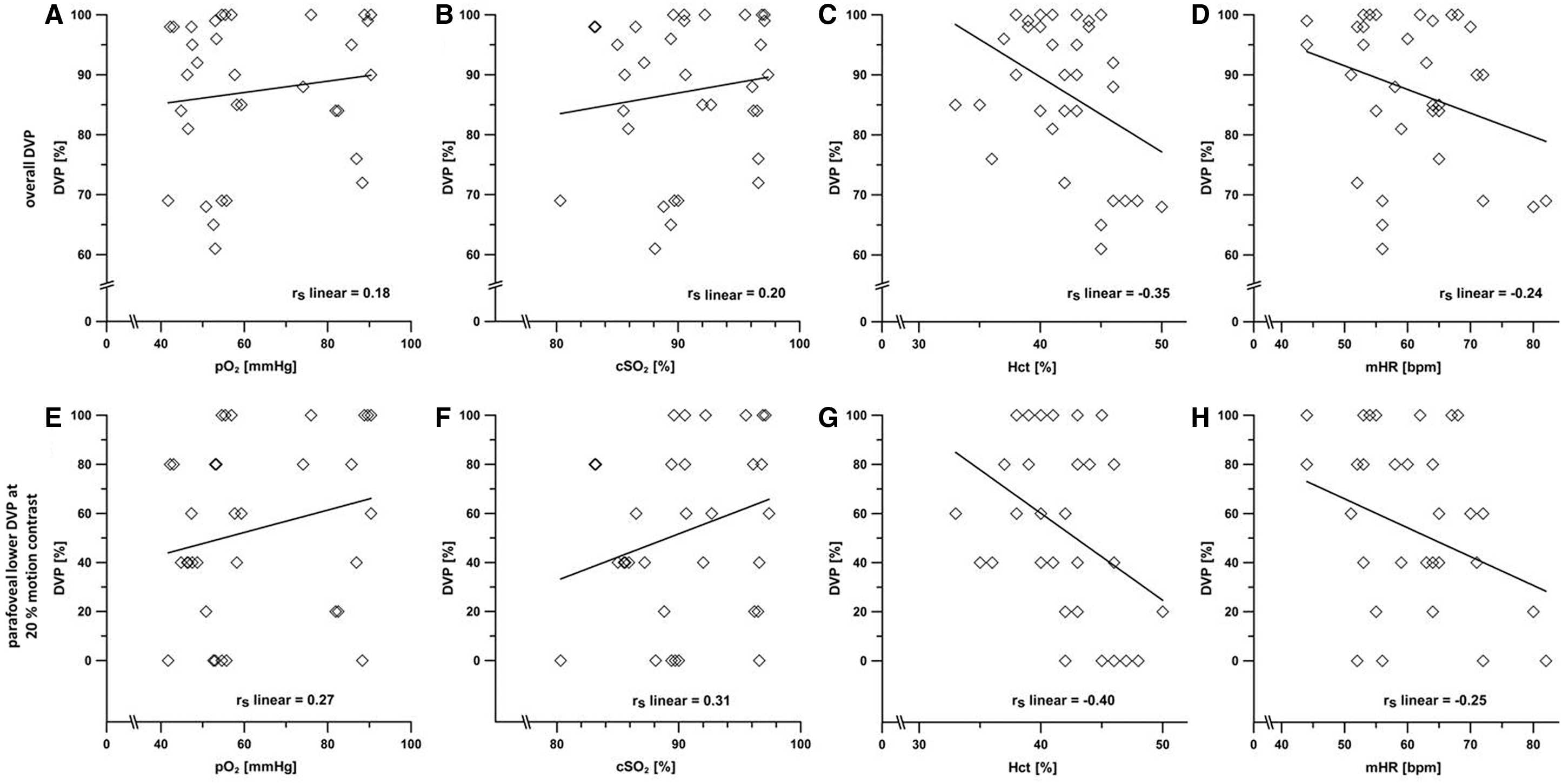
Correlation analyses between overall DVP of all subfields
Discussion
This study aimed to analyze the effect of high-altitude hypoxic conditions on DVP in healthy young people. Generally, and especially at high altitude, a “variety of disturbing factors act on the human body and affect its vital as well as sensory functions, vision being considered the most important sense for providing information” (Daumann and Draeger, 1993). The main finding of the study is a significant reduction in DVP of the lower parafoveal visual field in healthy young hikers at altitudes above 4500 m. Therefore, the common changes in visual perception performance are related to altitude, state of oxygenation, and health. Notwithstanding the mixed results regarding alterations in visual performance and lack of evidence, our results support the hypothesis that visual changes are highly individual and less common, respectively, are of little clinical significance at lower altitude (Virues-Ortega et al., 2004; Wilson et al., 2009). Therefore, the incidence of visual impairments at moderate and high altitudes (below 5000 m) might be lower, particularly with sufficient acclimatization (Willmann et al., 2010, 2014a, 2014c). On the other hand, there are reports of climbers who suffered from clinically significant ocular changes like retinal bleeding or corneal edema well below 5000 m (Hackett and Rennie, 1979; Honigman et al., 2001) and studies have shown the serious consequences of visual impairment on motor control and balance regulation (Paulus et al., 1984).
The ascent profile, including an acclimatization phase, between days 2 and 4, at altitudes between 3000 and 4300 m (sleeping level: 3029 to 3647 m) prevents subjects from experiencing AMS complications and minimizes the physiological effects of acute hypoxia (Hackett and Roach, 2001; Bärtsch et al., 2004; Bärtsch and Swenson, 2013). We hope to learn more about the acute effects of stronger hypobaric hypoxic stimuli by comparing testing not only at different altitude levels but also between different levels of systemic hypoxic stress and different metabolic states. During the acclimatization period, mean cardiovascular parameters mHR and RRs/RRd followed the physiological adaptation mechanism for hypoxia (Young and Reeves, 2002). Between days 2 and 4, mHR remained unchanged (p > 0.05), decreasing on day 6 (compared to day 4) as predicted by acclimatization (p < 0.05). After climbing higher on day 6, cSO2 decreased (p < 0.05). Therefore, the level of systemic hypoxic stress on day 6 was greater than on day 5. Throughout the whole hike, sufficient hydration of the subjects led to equal mean Hct values. On the examination days, one subject showed symptoms of AMS (d5: LLS = 3; d6: LLS = 6), without subjective impairment of their concentration, while reporting mild fatigue (LLSFatigue) on day 5 (LLSFatigue = 1) and severe fatigue on day 6 (LLSFatigue = 3). We found no obvious influence of LLSFatigue on DVP (rs = −0.07; p > 0.05).
Measurements of DVP on the fifth day (d5) showed no changes. Only a trend of reduced lower DVP was found. Day 6 resulted in a reduction of DVP in the lower parafoveal visual field corresponding to the superior central retinal. Previous studies have shown that the retinal cells do not maintain their physiological functions under oxygen-reduced conditions (Brinchmann-Hansen and Myhre, 1989; Sponsel et al., 1992; Tinjust et al., 2002; Kergoat et al., 2006; Feigl et al., 2007). These studies have demonstrated that acute hypoxia impairs the activity of ganglion cells, rod cells, and cone cells, and might lead to impaired visual perception (Brinchmann-Hansen and Myhre, 1989; Sponsel et al., 1992; Feigl et al., 2007). Tinjust et al. (2002) suggests that outer retinal cells are more resistant to systemic hypoxic stress than inner retinal cells based on analyses of photopic flash electroretinogram (ERG) and oscillatory potential recordings in normobaric hypoxia. They used hypoxic stimuli of 12% oxygen over an acute period of 5 minutes. This is in line with studies demonstrating that short-term hypoxia in the central part of the retina reduces post-photoreceptoral depolarizing and hyperpolarizing bipolar cells (Feigl et al., 2007). Recent studies have studied ERG during prolonged hypobaric hypoxia at in the Capanna Margherita at 4559 m (Schatz et al., 2013, 2014).
In the same condition, we found significant reduction in DVP of the lower parafoveal visual field in healthy young hikers; Schatz et al. (2013) were able to show that retinal function of inner, outer, and ganglion cell layer is altered at high-altitude hypoxia. Notably, differences in the severity of the expression of ERG parameters had shown a dominate effect on post-photoreceptoral signaling pathways, including phototransduction and visual processing. In contrast to acute simulated conditions (Tinjust et al., 2002; Feigl et al., 2007), missing differences in ERG responses considered to represent function of amacrine cells, bipolar cells, and interplexiform cells in hypobaric hypoxia lead to the assumption of longer lasting effects inter alia on ganglion cells. Human ganglion cells are overrepresented in the central and superior retina compared to the inferior peripheral retina (Curcio and Allen, 1990). Together, both findings possibly provide a putative anatomically functional explanation to the impairment of the superior retina in particular. Accordingly, systemic hypoxic stress levels at 4500 m or higher (∼12% atmospheric oxygen concentration) might trigger impairment of DVP because of uncompensated oxygen deficits in the superior central inner layers of the retina. In agreement with Schatz et al. (2013), we suggest differences between retinal cells and their adaptation to hypoxia. Ganglion cells and other cells of the inner retinal layers might acclimatize slower or inadequate compared to retinal cells of the outer layer.
In contrast, central controlled efferent DVP (saccadic eye movements) of healthy people is not affected by rapid ascent to 3459 m (Faull et al., 2015) or chronic high altitude (Merz et al., 2013). This might indicate that central nervous processing of visual information is more resistant to decreased systemic oxygen than peripheral parts of the brain. Unlike parafoveal DVP, investigations have shown that static foveal visual performance (such as best-corrected visual acuity) is unaffected by acute or chronic hypobaric hypoxia (Bosch et al., 2008; Fischer et al., 2012; Gekeler et al., 2019; Krusche et al., 2019), supporting the idea that foveal parts of the retina are less sensitive to hypoxic stimuli compared to the parafoveal or peripheral retinal space. In addition, Horng et al. (2008) demonstrated a strong decrease of black-and-white vision in the periphery of the visual field with simulated severe hypobaric hypoxia at 7620 m. Despite dissimilar processing of dynamic and static visual information in the brain (Ungerleider and Pasternak, 2004), central processing might not be affected in the same way the retina do by hypoxic stress.
It is speculated that reduction in parafoveal lower DVP at high altitude adversely affects motor performance and coordination, which might increase the risk of accidents. Vision permits proactive adaptations to guarantee safe and efficient locomotion (Reynard and Terrier, 2015). It allows us to navigate our environment, avoiding obstacles and ensuring correct orientation (Patla, 1997). Occlusion of the peripheral visual field increases postural control more than central occlusion of the fovea (Paulus et al., 1984). Due to a lack of data, it is unclear if alterations to DVP affect balance control or motor performance. Further studies should focus on the suspected correlation between impairments in DVP and stabilization of posture.
There are several limitations to our study, but we do not feel they detract from the main findings. In our study, due to the nature of high-altitude exposure and associated physiological changes, confounding factors make it difficult to compare and reproduce the results following the STAR initiative (Brodmann Maeder et al., 2018). We studied DVP during a representative high-alpine hike closed to recreational sports instead of using a well-established exposure protocol by Bärtsch et al. (1988) to Capanna Margherita like other groups (e.g., the THAO-Project: Fischer et al., 2012; Schatz et al., 2013; 2014; Gekeler et al., 2019). Potential side effects because of prolonged hiking at high altitude like fatigue, exhaustion after climbing, or reduced vigilance as consequence of poor sleep (Pun et al., 2018) did not control in a comprehensive protocol. The setting of the study, therefore, highlighted the practical reference, but at the same time reduces the quality of the correlation analysis of AMS and DVP. Unfortunately, we did not evaluate the effects of acute hypobaric hypoxia on DVP at the beginning of the hike when LLS peaked on d2. In this context, exercise at high altitude has been reported to aggravate retinal alterations (McFadden et al., 1981).
Nevertheless, despite special precautions (such as compliance to resting times before testing), the statistical data should be evaluated prudent. In addition, despite accurate statistical awareness, the results have to be handled with care because the examined cohort in this study was relatively small to assess perception performance. A comparison with previous studies is not able to be performed because of missing data in this field of research. One more limitation was that we examined moderately trained, healthy, and young subjects. The effect of hypobaric hypoxia in a differently trained, aged, or diseased cohort was still unidentified. Because of the active nature of the project (continuously hiking and climbing without the support of any cable car, helicopter, or similar) storage space for research equipment was limited. Thereby, we did not integrate extensive material to measure markers for cerebral activation or oxygenation in the visual areas of the occipital lobe to generate more precise indices for the localization of hypoxia-induced effects on DVP.
Conclusion
To summarize, we demonstrated that an investigation of afferent DVP under standardized conditions is feasible while hiking at high altitude. DVP in the lower parafoveal visual field of young, healthy, physically fit people was affected by altitudes above 4500 m. Properly acclimatized hikers may suffer from significant effects of acute enhanced hypoxia on their lower parafoveal DVP during a long-term stay at high altitudes. In agreement with Willmann et al. (2010, 2014a), we hypothesize that physical adaptation to transient hypoxia by sufficient acclimatization reduces alterations of visual performance in general. Strategies helping mountaineers reaching high altitudes without severe complications might be advisable to ensure the prevention of visual impairments and safe traveling.
Footnotes
Acknowledgments
The authors would like to thank Professor Eugene R. Wist, who provided the Düsseldorf Test for Dynamic Vision, and all participants taking part in the study.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors received no specific funding for this work.