
Abstract
Background/Aims:
Long-term high altitude residence leads to a sustained increase in pulmonary vascular resistance and elevation of pulmonary artery pressure due to chronic alveolar hypoxia. However, living at high altitude is also associated with other environmental factors such as cold. There is still little experimental evidence suggesting detrimental effects of low temperatures on the pulmonary vasculature. Therefore, our objective was to investigate acute effects of cold exposure on the pulmonary circulation in Kyrgyz high altitude natives.
Methods:
Responses of the pulmonary circulation during acute exposure to controlled cold conditions (4°C–6°C) for 60 minutes were measured in highlanders using Doppler echocardiography. Based on the Doppler echocardiography-derived tricuspid regurgitant systolic pressure gradient (TRG), subjects with TRG ≥40 mmHg were allocated into the pulmonary hypertension (PH) group. Participants from the PH group were compared with volunteer control subjects with TRG <40 mmHg. All baseline measurements were evaluated in a warm room during 60 minutes (22°C–28°C). Following baseline echocardiography, the subjects were assigned to either warm or cold exposure for an additional 60 minutes.
Results:
Acute cold exposure significantly increased TRG both in the control (ΔTRG, 4.93 mmHg) and in the PH (ΔTRG, 8.15 mmHg) group, compared to the respective warm exposure conditions (ΔTRG, −0.14 and −0.05 mmHg). No changes in cardiac output were observed upon cold exposure.
Conclusion:
Thus, acute exposure to cold leads to elevation of pulmonary artery pressure in high altitude residents.
Introduction
The consequences of the long-term high altitude residence, such as augmentation of pulmonary vascular resistance and increase in pulmonary artery pressure, are well-known phenomena. High altitude pulmonary hypertension (PH) appears due to chronic alveolar hypoxia exposure, which subsequently causes hypoxic vasoconstriction and development of pulmonary vascular remodeling. There is good evidence that hypoxia-inducible factors (HIFs) are key transcriptional regulators of the hypoxia-induced structural remodeling of the pulmonary vascular bed (Yu et al., 1999; Brusselmans et al., 2003). Furthermore, animal studies suggest that susceptibility to high altitude PH has a genetic basis (Zhao et al., 2001; Rhodes, 2005). Genome-wide selection scans of Tibetan DNA revealed variants in and around two HIF pathway genes, EPAS1 (HIF-2α) and EGLN1 (HIF prolyl 4-hydroxylase 2, PHD2) (Beall et al., 2010; Bigham et al., 2010; Simonson et al., 2010; Yi et al., 2010; Peng et al., 2011). Remarkably, the polymorphisms in EPAS1 and EGLN1 in Tibetans correlate with hemoglobin concentration (Beall et al., 2010; Bigham et al., 2010; Yi et al., 2010; Peng et al., 2011). However, whether the variants in EPAS1 and EGLN1 are also associated with protection from high altitude PH in Tibetans is not known. Importantly, recent meta-analysis did not support the current belief of differences in pulmonary artery pressure levels between different high altitude populations (Soria et al., 2016). Moreover, no differences in pulmonary artery pressure levels were observed between acclimatized Europeans and Bolivian Aymara or Sherpas (Schwab et al., 2008; Foster et al., 2014). Thus, factors other than hypoxia (and HIF) may contribute to development of high altitude PH.
In addition to alveolar hypoxia, the high altitude geographic settings are often characterized by several other challenging abiotic factors, including higher solar radiation, strong winds, and particularly the prolonged periods with low temperatures. There is evidence supporting detrimental effects of cold exposure on the pulmonary circulation in animals (Kashimura, 1993; Watanabe et al., 2007). In rats, chronic cold exposure resulted in the development of the classical pathological features of PH, such as pulmonary vascular remodeling, increased right ventricular (RV) systolic pressure, and RV hypertrophy (Crosswhite and Sun, 2013; Crosswhite et al., 2014). Notably, cold stress has been shown to be one of the environmental factors for development of RV hypertrophy and ascites in broilers (Sato et al., 2002). Interestingly, a recent study compared echocardiography-derived hemodynamic parameters obtained in ambulatory patients during hotter months (June–November) to those obtained in colder months (December–May) and revealed an inverse correlation of climate indices with systolic pulmonary artery pressure (Rozenbaum et al., 2018). Moreover, echocardiograms performed during colder months had a higher chance to show systolic pulmonary artery pressure >40 mmHg compared to those conducted in hotter months (Rozenbaum et al., 2018). Furthermore, signs of PH and RV hypertrophy were revealed in long-term residents and natives of Canadian Arctic and Russian Northeast, as we have previously summarized and discussed (Petrovic et al., 2015). To the best of our knowledge, no studies have investigated effects of cold exposure on pulmonary circulation in high altitude natives. Therefore, the aim of the study was to assess pulmonary hemodynamics in high altitude residents exposed acutely to cold.
Materials and Methods
Study subjects and experimental protocol
This independent project was part of a larger study investigating the cardiovascular system in high altitude natives, which included several high altitude expeditions to Alay region. This study was conducted in Sary-Mogol village (∼3000 m, Kyrgyzstan) during the late winter (February–March 2017) (Fig. 1a).
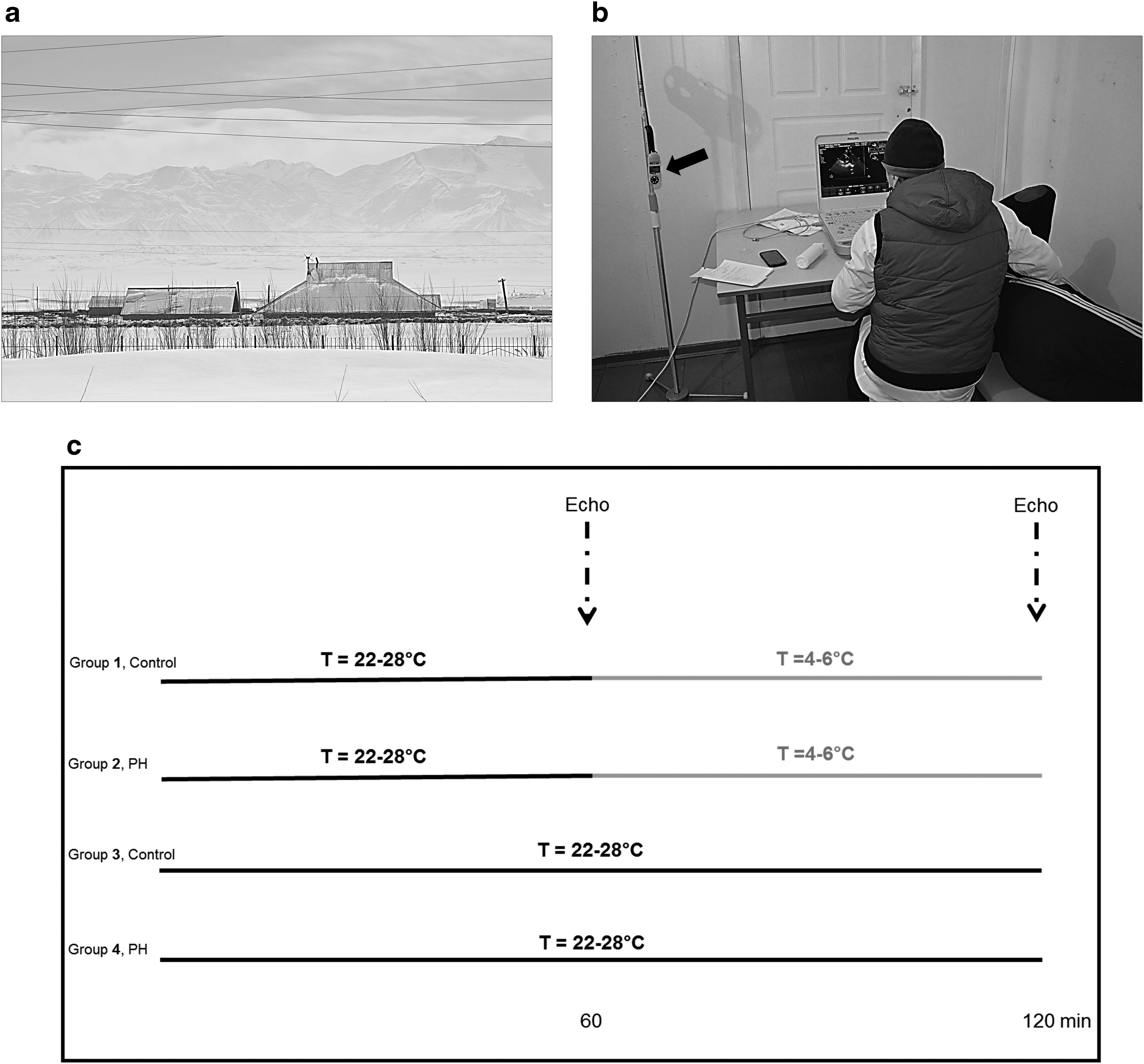
High altitude location and experimental design.
Male and female highlanders of Kyrgyz ethnicity living in the Sary-Mogol and from neighborhood villages were invited to participate in studies performed at the local primary health care center. Criteria of inclusion were age between 18 and 70 years. Inclusion criteria also specified that they were born and had lived permanently at their resident altitude. All patients underwent functional evaluation before inclusion: thorough medical history, physical examination, blood pressure measurement, pulse oximetry, spirometry, complete blood cell count, 12-lead electrocardiography, and echocardiography. Exclusion criteria considered major cardiovascular and respiratory diseases, active infections, pregnancy, and excessive erythrocytosis (hemoglobin concentration in females >19 g/dL, in males >21 g/dL).
Based on the Doppler echocardiography-derived tricuspid regurgitant systolic pressure gradient (TRG), in mmHg, we selected subjects with TRG ≥40 mmHg (most of them had been investigated during previous expeditions), which were then allocated into the PH group. Participants from the PH group were matched with volunteer control subjects with TRG <40 mmHg by age, sex, and body mass index (BMI). After completion of initial screening, participants were familiarized with the study procedures. The participants were randomly assigned to either warm exposure or cold exposure. Detailed characteristics describing the demographic and physiological parameters of the enrolled subjects are provided in Table 1.
Demographic and Physiological Values in Human Subjects
Values are mean (SD). Comparisons are made between the groups.
No significant differences were present between the groups (one-way ANOVA or the Kruskal–Wallis test for non-normally distributed data).
BMI, body mass index; SBP, systolic blood pressure; DBP, diastolic blood pressure; SpO2, arterial oxygen saturation; FEV1, forced expiratory volume in 1 second; PH, pulmonary hypertension; ANOVA, analysis of variance; SD, standard deviation.
Written informed consent was obtained from all study participants. This study was approved by the Ethics Committee of the National Centre for Cardiology and Internal Medicine, Bishkek, Kyrgyzstan and by the Institutional Ethical Review Board of the Faculty of Medicine at Justus-Liebig University, Giessen, Germany. The study was conducted following the Declaration of Helsinki principles.
Acute exposure to cold: experimental design
After baseline echocardiography, which was performed ∼60 minutes following the initial exposure to the warm ambient temperature of 22°C–28°C in a temperature-controlled room, the subjects moved to another room in the same building where the ambient temperature was kept at around 4°C–6°C (Fig. 1b, c). During the exposure period to cold (60 minutes), the subjects were lightly dressed. At the end of the experiment, echocardiography was performed for the second time. In the warm exposure group, the subjects remained in the temperature-controlled room and were exposed to the ambient temperature of 22°C–28°C for additional 60 minutes (Fig. 1c). After that, echocardiographic assessment was conducted again. Therefore, there were four experimental groups: (1 and 2) human subjects with PH or controls exposed for 60 minutes in the warm room (after initial 60 minutes in the same room), and (3 and 4) human subjects with PH or controls exposed for 60 minutes in the cold room (after initial 60 minutes in the warm room).
Physiological measurements
Blood pressure was measured using an automated blood pressure cuff (Omron M300; Omron Healthcare) on the nondominant arm with the subject sitting upright after at least 5 minutes rest. Simultaneous resting recordings of oxygen saturation (SpO2) and heart rate were performed using a finger pulse oximeter (WristOx® 3100; Nonin Medical, Inc., Plymouth, MN). The mean of three measurements separated by a 3-minute interval was used in the analyses for blood pressure, heart rate, and SpO2. In addition, we have performed the spirometry in all subjects using MicroLab 3500 (CareFusion) following the international guidelines. Finally, the BMI values were obtained following the formula: BMI = weight (kg)/height (m)2.
Transthoracic echocardiography
M-mode, two-dimensional, and Doppler echocardiography were performed from the standard parasternal, apical, and subcostal views on fully rested subjects in the supine or left lateral decubitus position using a portable ultrasound system (CX50 CompactXtreme; Philips, Bothell) with the S5-1 sector array transducer. Quantification of cardiac chamber size and function was performed in accordance with the recommendations of the American Society of Echocardiography (Lang et al., 2005, 2006; Rudski et al., 2010). The maximal velocity of the tricuspid regurgitant jet (TRVmax) was measured using continuous-wave Doppler in the apical four-chamber view. TRG was calculated from the TRVmax using the simplified Bernoulli equation: TRG = 4 × TRVmax2. Cardiac output (CO) was determined by measuring the diameter (D) of the left ventricular outflow tract (LVOT) and LVOT time–velocity integral (VTI) of its Doppler signal as follows: CO = π × (D/2)2 × VTI × heart rate (Quinones et al., 2002). LVOT VTI was calculated by placing the pulsed wave Doppler sample volume in the outflow tract below the aortic valve in the apical five-chamber view.
Hemoglobin
Complete blood count was performed by an automated hematology analyzer Mindray BC-2300 (Guangzhou Shihai Medical Equipment Co., Ltd., China) using 15 μL of ethylenediaminetetraaceticacid whole venous blood according to the manufacturer's instructions.
Statistical analysis
All data analyses were performed using GraphPad Prism version 6.05 for Windows (GraphPad Software, La Jolla, CA; www.graphpad.com). Data are expressed as mean value ± standard deviation. Normal distribution of the data was tested using the D'Agostino-Pearson normality test. Demographic and physiological group characteristics were compared using one-way analysis of variance (ANOVA) or the Kruskal–Wallis test for non-normally distributed data. Changes in absolute values of CO were analyzed by two-way ANOVA with Sidak's multiple comparisons test. Finally, the TRG values were compared using the paired and unpaired t test. A p-value <0.05 was considered significant for all analyses.
Results
Demographic and physiological characteristics
Initially, 52 individuals were recruited into the study. During the execution of the study, five participants withdrew because of personal reasons. Of these subjects, one, three, and one participant were from the control warm exposure, control cold exposure, and PH cold exposure groups, respectively. The data from a total of 47 participants were available for analysis. Table 1 shows demographic and physiological characteristics of the participants in each group. There were no significant differences in age, BMI, blood pressure, peripheral oxygen saturation, or spirometric parameters between the experimental groups.
Response of the pulmonary circulation to acute cold
Baseline TRG values measured after initial 60 minutes of exposure to warm ambient conditions and before further exposure to the same conditions for additional 60 minutes were significantly elevated in the PH group (48.6 ± 5.8 mmHg), compared to the respective control group (27.8 ± 3.2 mmHg; p < 0.05) (Fig. 2). Similarly, baseline TRG values measured after initial 60 minutes of exposure to warm ambient conditions and before exposure to cold conditions for 60 minutes were significantly elevated in the PH group (46.8 ± 5.7 mmHg), compared to the respective control group (30.6 ± 4.9 mmHg; p < 0.05) (Fig. 3). Importantly, TRG values did not differ between the control groups or among the PH groups (Figs. 2 and 3). As expected, there were no changes in TRG values in both groups (PH and control) after further exposure to the warm conditions for another 60 minutes (Fig. 2). However, there was a significant increase in TRG values in both groups (PH and control) after 60 minutes of cold exposure (Fig. 3a, b). Finally, CO values did not differ between the experimental groups. There were no significant effects neither by condition (control vs. PH) nor temperature (warm vs. cold) and no interaction effect for CO (Fig. 4).

Effects of warm exposure on pulmonary circulation. Individual responses in highlanders (n = 12–13) with low TRG

Effects of cold exposure on pulmonary circulation. Individual responses in highlanders (n = 10–12) with low TRG

Effects of warm and cold exposure on cardiac output. Changes in absolute values of cardiac output (ΔCO, in L/min) in highlanders (n = 10–13) with low TRG (control) and those with elevated TRG (PH) exposed either to warm or cold temperatures are depicted. Data are expressed as mean ± standard deviation.
Discussion
In this study, we report for the first time the effects of acute cold exposure on pulmonary circulation in human high altitude residents of Kyrgyzstan. In general, we demonstrated that acute cold exposure significantly increased TRG, which was independent of the baseline TRG values.
Earlier observations described severe hypoxic PH and RV failure that develops in susceptible cattle after their exposure to high altitudes (Glover and Newsom, 1915, 1918). Remarkably, animals developed PH with RV failure more often if they were brought to high altitudes during the cold months (Glover and Newsom, 1915). Notably, previous experiments demonstrated a progressive increase in pulmonary vascular resistance during hypothermia in anesthetized dogs (Kuhn and Turner, 1959), suggesting a contributory role of cold condition to PH development. Furthermore, cooling of the skin resulted in augmented pulmonary vascular resistance in cattle living at the moderate altitude of 1524 m (McMurtry et al., 1975). Moreover, cold pressor test (insertion of the left hand and lower third of the forearm in crushed ice for 2 minutes) performed at low altitude led to a rise in pulmonary arterial pressure and pulmonary vascular resistance in subjects with a previous history of high altitude pulmonary edema, a condition characterized by an abnormal pulmonary artery pressure response to hypoxia and exercise in susceptible persons (Masud ul Hasan Nuri et al., 1988). Consistent with these findings, it was noticed that calves with an exaggerated hypoxic pulmonary vasoconstriction experienced more prominent effects of acute environmental cold on the pulmonary circulation (Will et al., 1978).
In contrast, we observed a TRG increase both in control high altitude residents and those with established PH. Lower temperatures, longer duration of exposure to cold of cattle, and lower altitude may account for this discrepancy. Indeed, experiments in awake rats demonstrated a gradual increase in mean pulmonary artery pressures with the drop in ambient temperature from 5°C to −10°C (Kashimura, 1993). In contrast, placement of cold-exposed calves into a warm environment for 24 h led to a decrease in pulmonary artery pressures (Will et al., 1978). Correspondingly, slightly higher pulmonary artery pressures were observed in highlanders at ambient temperatures (19°C) than in a thermoneutral environment (Lockhart and Saiag, 1981).
Experimental evidence demonstrated that cold-induced elevation in pulmonary artery pressures was not accompanied by a rise in CO suggesting that cold-induced PH is primarily caused by pulmonary vasoconstriction (Chauca and Bligh, 1976; Busch et al., 1985). In accordance with these findings, we did not observe any changes in CO in highlanders during acute exposure to cold.
Furthermore, it has been suggested that hyperreactive pulmonary vessels might contribute to PH at high altitude. Interestingly, studies in calves with normoreactive pulmonary vessels suggested that hyperreactive vasculature is not crucial for the cold-induced pulmonary pressure rise (Busch et al., 1985). In addition, no interaction between effects of cold and hypoxia on the pulmonary circulation was shown in animals (Chauca and Bligh, 1976; Busch et al., 1985). We demonstrated that acute exposure to cold environment leads to elevation of pulmonary artery pressure in human high altitude residents. This response is probably due to pulmonary vasoconstriction and independent of the baseline levels of pulmonary artery pressure. Studies in cattle clearly showed that hypoxic pulmonary vasoconstriction due to alveolar hypoventilation contributes to cold-induced pulmonary artery pressure elevation (Will et al., 1978; Busch et al., 1985). In contrast to cattle and other animals, humans respond to cold exposure by hyperventilation (Cogo, 2011). Therefore, we believe that in humans hypoxic pulmonary vasoconstriction plays less significant role if at all in acute cold-induced pulmonary artery pressure rise compared to animals.
In addition, numerous studies implicated sympathetic nervous system in cold-induced changes of the cardiovascular system (Papanek et al., 1991; Sun, 2010). There is a substantial experimental evidence of significantly increased levels of circulating and urinary epinephrine and norepinephrine in cold-exposed animals (Sun, 2010). However, chronic cold exposure has been shown to reduce vascular responsiveness to phenylephrine (Fregly and Brummermann, 1993), suggesting desensitization of vascular α1-adrenoceptors. Consequently, since our study was performed in the end of the cold season (i.e., chronic cold exposure), we assume a minor contribution of epinephrine and norepinephrine to the cold-induced responses. Nevertheless, cold activates a number of other systems, including renin-angiotensin and endothelin-1 systems and dysregulation of the nitric oxide pathway (Sun, 2010), which all play an important role in regulation of pulmonary circulation. However, evidence of their involvement in cold-induced pulmonary artery pressure elevation is still lacking.
Further work is necessary to investigate effects of chronic cold exposure on pulmonary circulation in human high altitude natives and to precisely characterize the development of the pulmonary vascular disease in such environmental conditions.
Footnotes
Acknowledgments
The authors thank the study participants for giving up their time to take part in this study. In addition, the authors thank Professor Martin Wilkins from Imperial College in London for his valuable scientific advice.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Cardiovascular Medical Research and Education Fund, the Ministry of Education and Science of the Kyrgyz Republic (No. 0005823), and the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) - Projektnummer 268555672 - SFB 1213, A09.