
Abstract
Background:
In high-altitude animals, brain cell resilience against hypoxia stress is one critical evolutionary step that has promoted individual survival and species adaptation to the environment. Aquaporin-4 (AQP4) is implicated in a number of physiopathological processes, particularly in the development of brain edema, and other functions such as the regulation of extracellular space volume, potassium buffering, waste clearance, and calcium signaling. Still, the role of AQP4 in the adaptation to high-altitude hypoxia remains unknown. The yak (Bos grunniens) is the only large mammal that is currently known to have adapted to the high-altitude hypoxic environment of the Qinghai-Tibet Plateau (>4000 m above sea level).
Methods:
In this study, we cloned the complementary DNA (cDNA) for yak AQP4 and analyzed structural differences of AQP4 between yak and cattle. We used reverse transcription quantitative polymerase chain reaction and western blot to investigate whether the expression of AQP4 mRNA and protein was different in brain of yak and cattle. In addition, immunohistochemistry was use to analyze the localization and expression of AQP4 in brain of yak and cattle.
Results:
Immunohistochemical results have shown that AQP4 is expressed in many regions of the yak brain, and both protein and messenger RNA (mRNA) levels are significantly lower than those of low-altitude cattle (Bos taurus). Phylogenetic analysis revealed that yak AQP4 is evolutionarily conserved. Interestingly, a substitution of Ala (cattle) to Ser in position 82, and eight additional amino acid residues composing an α-helix region are present in yak AQP4 protein. These sequence modifications potentially modulate the function of AQP4 in distinct environments.
Conclusions:
Our findings suggest that AQP4 may have an important role in the resistance to cerebral edema through low expression and maintenance of normal physiological function in the yak brain.
Introduction
A series of studies have demonstrated that hypoxia is a major stress factor that induces various diseases. In the context of homeostasis, the brain is the most sensitive organ to changes in oxygen levels. In fact, when the oxygen demand for the brain is not met, hypoxia acts as an initial trigger for physiopathological changes that include oxidative stress and edema formation.
High altitude is the most important ecological challenge for plateau animals. The yak (Bos grunniens), one of the large native high-altitude animals, have developed a unique brain adaptation mechanism that ensures successful survival to the harsh environment (Ding et al., 2019).
Aquaporin-4 (AQP4) is one of the most abundant molecules in the brain. The expression of AQP4 is particularly prevalent in astrocytic membranes at the blood–brain barrier and brain-liquor interface (Amiry-Moghaddam et al., 2003; Badaut et al., 2011). AQP4 has been shown to be involved in a number of physiological processes, including (1) regulation of extracellular space volume (Badaut et al., 2011; Jin et al., 2013), (2) potassium buffering (Eid et al., 2005), (3) cerebrospinal fluid circulation (Nagelhus and Ottersen, 2013; Papadopoulos and Verkman, 2013), (4) waste clearance (Iliff et al., 2012), (5) cell migration (Fan et al., 2013), and (6) calcium signaling (Thrane et al., 2011). Importantly, it has been reported that AQP4 also plays a critical role in regulating the formation of brain edema (Amiry-Moghaddam et al., 2004; Assentoft et al., 2015; Ikeshima-Kataoka, 2016).
AQP4 is a predominant water channel molecule found in the mammalian brain. Since brain edema is an inflammatory condition closely related to changes in water transport, we hypothesized that AQP4 may have a differential expression in high-altitude animals like the yak. As a result, in this study, we cloned the open reading frame sequence of yak AQP4 and determined its expression pattern in vivo. Our results indicate that AQP4 has distinctive tissue and altitude-specific expression patterns in yak, suggesting a potential role controlling brain homeostasis in hypoxic conditions.
Materials and Methods
Sample collection
All research protocols used in these experiments were approved by the Animal Ethics Committee of the Gansu province, China. The brains of 10 healthy yak (5 males and 5 females, 4- to 7-year-old) with a mean body weight of 250 kg (standard deviation [SD] ±20 kg) were collected immediately after slaughter from a local abattoir, Maqu County (>3500 m above sea level), Gannan Tibet Autonomous Prefecture, Gansu Province, China. For comparison, the brains of 10 healthy cattle (Bos taurus; 5 castrated and 5 female, 3- to 6-year-old) of similar body weight (275 kg, SD ±25 kg) were collected immediately after slaughter from a local abattoir, Guanghe County (<1500 m above sea level), Linxia Hui Nationality Autonomous Prefecture, Gansu Province, China (Ding et al., 2018). Every sample was divided into two parts on average, one part was fixed with 4% paraformaldehyde in 0.03 M sodium phosphate buffer, pH 7.4, and the other part was divided into different encephalic regions, including cerebral cortex (frontal lobe), thalamus, caudate nucleus, and hippocampus, snap-frozen in liquid nitrogen, and then stored at −80°C before use.
RNA and protein isolation
Total RNA of 100 mg tissue from the abovementioned samples was extracted using Trizol reagent (Invitrogen, Carlsbad, CA) as described by the manufacturer's instructions, its purity and concentration were measured using NanoDrop 2000, the integrity of the RNA was examined by electrophoresis, and complementary DNAs (cDNAs) were obtained from 2 μg total RNA using the Revert Aid™ Reverse Transcriptase (Takara, Dalian, China) according to the manufacturer's instructions.
For protein isolation, 100 mg frozen samples was ground in liquid nitrogen and transferred to centrifuge tubes with RIPA/PMSF (Solarbio; Beijing Solarbio Life Science and Technology Co. Ltd., Beijing, China). After fully blending and mixing until a pink color was obtained, the sample tubes were incubated on a spiral oscillator for 2 hours (200 r/h) on ice. After centrifugation at 4°C for 10 minutes (12,000 r/h), the complete divided protein was obtained. Total protein concentration of samples was determined using a BCA protein assay kit. The total protein concentration was then adjusted to the same level, and 4 × sample buffer at 100°C was added for 12 minutes to completely denature the protein.
cDNA cloning and sequence analysis
The messenger RNA (mRNA) sequence of yak AQP4 (KM609432) was previously determined in our laboratory and deposited in the GenBank sequence database provided by the National Center for Biotechnology Information (NCBI). The specific steps are as follows: the primer pairs for yak AQP4 CDS amplification (Table 1) were designed based on the cattle AQP4 sequence (BC118415) by using Primer 5.0 (PREMIER Biosoft International, Palo Alto, CA). The polymerase chain reaction (PCR) was conducted by using Taq DNA polymerase (TaKaRa), the PCR cycling parameters were denatured for 2 minutes at 98°C, 35 cycles of 98°C for 10 seconds, 60°C for 15 seconds, and 72°C for 1 minute; and a final extension for 5 minutes at 72°C. The predominant PCR product was purified by gel (Eppendorf, Hamburg, Germany) and subsequently cloned into the pEGFP-C1 vector (Promega, Madison, WI) at EcoRI and KpnI sites to produce plasmid pEGFP-AQP4. The recombinants were identified through blue–white color selection in ampicillin-containing LB plates and confirmed by PCR. Positive clones were sequenced in both direction (Sangon, Shanghai, China), and DNAMAN software (Lynnon Biosoft, Vaudreuil, Canada) was employed to align the amino acid sequences among nine different species (Supplementary Data).
Primer Pairs Designed for the Yak Gene Aquaporin and GAPDH
AQP4, aquaporin-4.
Structure analysis of AQP4 protein
Amino acid sequence was aligned by using DNAMAN software. Kyte-doolittle hydrophobicity was predicted with ProtScale. The transmembrane regions were predicted with TMHMM. The secondary structure was predicted by using PSIPRED Protein Sequence Analysis Workbench. Prediction of transmembrane structure was calculated using the ExPASy tool ProtScale. Phosphorylation sites were predicted by NetPhos 2.0 Server. Tertiary structures were modeled using the SWISS-MODEL Workspace (Arnold et al., 2006; Kiefer et al., 2009).
RT-qPCR to determine the expression of AQP4 mRNA
The yak and cattle AQP4 gene mRNA levels in different encephalic regions were detected, including cerebral cortex, thalamus, caudate nucleus, and hippocampus. The reverse transcription quantitative polymerase chain reaction (RT-qPCR) was performed on the FastStart Universal SYBR Green Master (Rox) reagent mix (Roche, Basel, Sweden) with the following program: 50°C for 2 minutes, 95°C for 10 minutes, followed by a cycle of 95°C for 15 seconds, and 60°C for 30 seconds for 40 cycles. The special express primer of yak AQP4 was designed according to the sequences we cloned (Table 1). The relative mRNA expression of AQP4 was normalized to GAPDH using the relative quantification method 2−ΔΔCt method.
Protein expression analysis
Protein samples were thawed, mixed, and separated into beads using a spin column (Bio-Rad), and then separated on a 5% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) gel for Western blot (WB) analysis. After electrophoresis, the protein was transferred from the gel onto nitrocellulose membranes (Millipore Corporation, Billerica, MA). The membrane containing protein was blocked with 5% fat-free milk in tris-HCl buffer twain (TBST) at room temperature for 2 hours and hybridized using AQP4 (1:800; Abcam, Hong Kong, China) at 4°C overnight. The membrane was then washed four times with TBST and labeled with horseradish peroxidase-conjugated secondary antibody (1:2000; Bioss, Beijing, China) for 2 hours at room temperature. After washing five times with 1 × TBST, AQP4 was detected on the membrane using an enhanced chemilumescent detection kit (Beyotime, Shanghai, China). The intensities of the bands on the blots were measured using a densitometric analysis system (Bio-Rad). The intensity of the GAPDH band was used for normalization (Tang et al., 2017).
Immunohistochemical analysis
Paraformaldehyde-fixed paraffin-embedded tissues from cerebral cortex, thalamus, caudate nucleus, and hippocampus of yak and cattle were deparaffinized in xylenes and hydrated in a graded series of alcohols. Heat-induced epitope antigen retrieval using citrate buffer in the microwave oven was used. The above sections were stained with anti-AQP4 antibody, and standard immunohistochemical procedure was followed. The immunohistochemical stained sections were captured using Olympus BX50 epifluorescence microscope (Olympus Optical Co., Ltd.). The sections were used for assessing qualitative expression of AQP4, and the evaluators used the double-blind method to assess the expression.
To analyze the localization and expression of AQP4 in yak accurately, this study applied serial section to determine the localization according to the delineation of Modern Histology (Lingzhong et al., 2003).
Ultrastructural analysis
To detect the ultrastructure of cerebral cortex with prolonged ischemia and function of AQP4 in ischemia injury, the brains of yaks and cattle that were bled to death by the butcher were collected 5 minutes later after slaughtering. For comparison, the brains of animals were collected immediately after slaughter. So, there were four groups: yak-prolonged ischemic, yak-normal, cattle-prolonged ischemic, and cattle-normal group. For each group, three biological replicates were collected. Serial, coronal, ultrathin sections of the tissues were obtained and cut into 1 mm spaced pieces, placed in 3% glutaraldehyde phosphate buffer (pH = 7.4), and fixed for 2–4 hours, followed by 1% osmium tetroxide fixation, ethanol dehydration, and embedding in 812 epoxy resin. Ultrathin sections (60–70 nm) were then obtained from blocks of interest using a ultramicrotome, stained with Uranyl Acetate-Lead Citrate stain, and observed with a transmission electron microscopy-1220 (TEM-1220) operated at an accelerating voltage of 80 kV.
Measurement and statistical analyses
The brain in the WB images and immunohistochemical assays was measured using integrated optical density (IOD) and Image-Pro plus 6.0 (Media Cybernetics, Rockville, MD). Data were analyzed using SPSS 16 (SAS Institute, Cary, NC). Other data were analyzed by one-way analysis of variance and Duncan's post hoc test. p < 0.05 between groups was considered statistically significant (Liu et al., 2018).
Results
Bioinformatics characteristics of yak AQP4
The cDNA sequence of the yak AQP4 was cloned and submitted to GenBank (Accession number KM609432). The predicted amino acid sequence of the yak AQP4 was used to construct a phylogenetic tree to further evaluate its evolutionary relationship to other potential homologs (Fig. 1I). According to the multiple sequence alignment with homologs found in other species, including B. taurus (GenBank NM_181003), Pantholops hodgsonii (GenBank XM_005982605), Ovis aries (GenBank NM_001009279), Equus caballus (GenBank XM_001494280), Equus asinus (GenBank XM_014828373), Ochotona princeps (GenBank XM_004579580), Homo sapiens (GenBank NM_001650), and Mus musculus (GenBank NM_009700), we found that AQP4 protein is well conserved among these species. The tree structure indicates that yak, cattle, and sheep AQP4 homologs were phylogenetically closer to each other, which is consistent with the zoological classification. Amino acid sequence comparison between the AQP4 in yak and cattle, Tibetan antelope, sheep, horse, donkey, pika, human, and mouse revealed a high homology of 99.69%, 99.07%, 98.81%, 98.14%, 97.83%, 96.26%, 96.28%, and 93.19%, respectively.
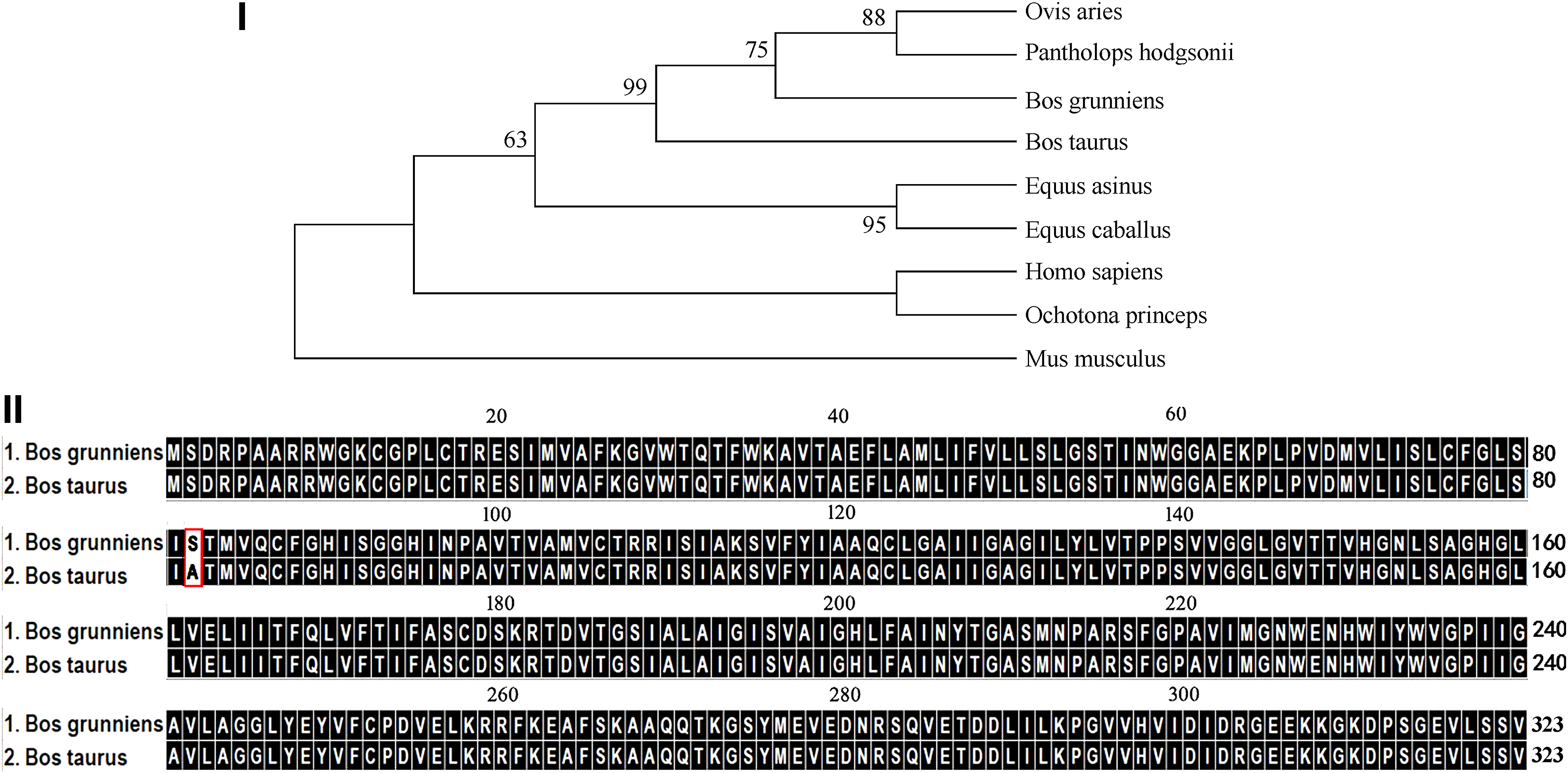
Comparison of AQP4 bioinformatics features of yak and cattle.
The mammalian AQP4 homologs described in this study consist of 323 amino acid residues. These are strongly conserved throughout different species. Nevertheless, an amino acid substitution at position 82 was detected, where in the yak AQP4 was Ser (a polar amino acid) instead of Ala (a nonpolar amino acid) present in cattle (Figs. 1II and 2III).
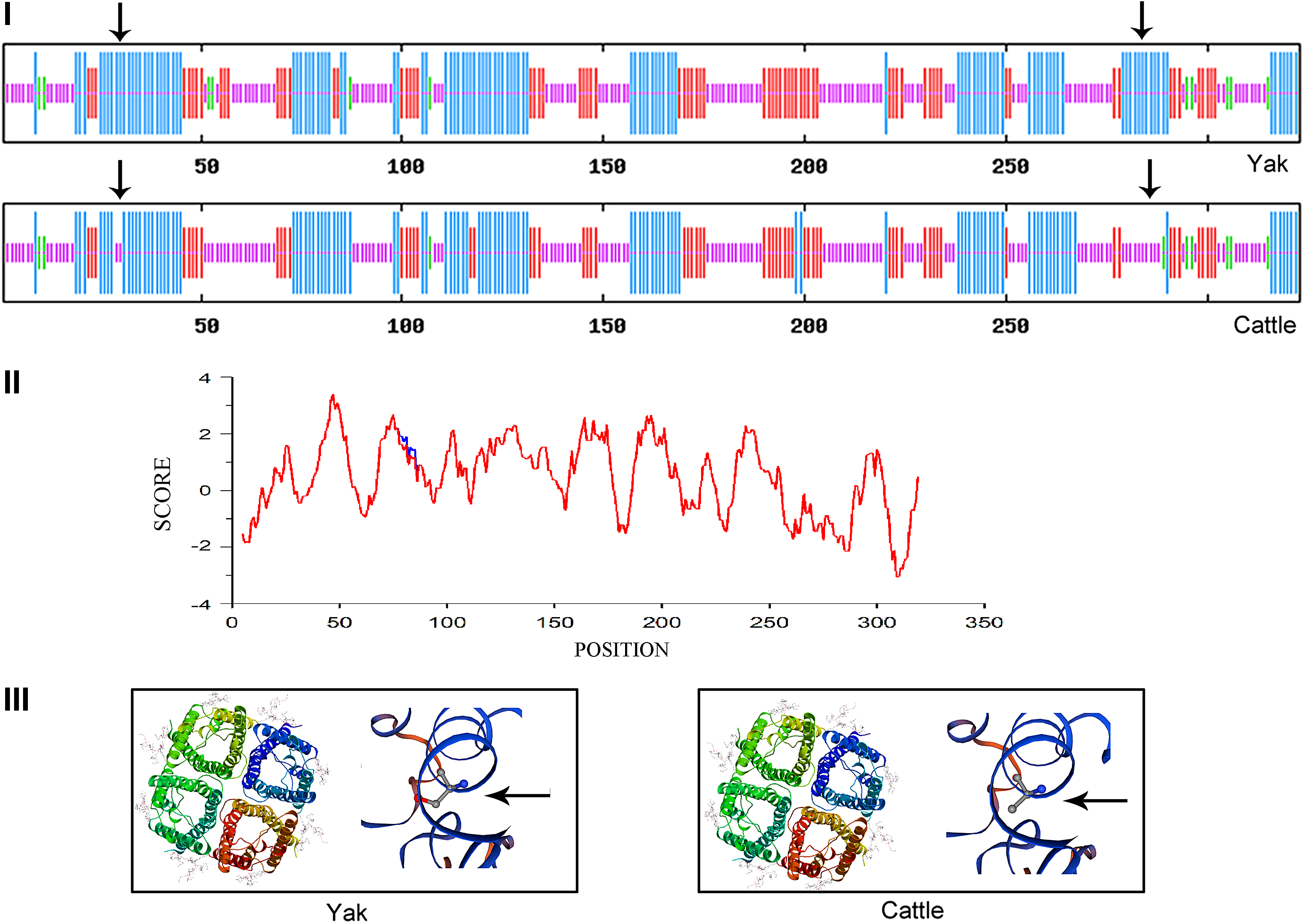
Predicted structure and function for AQP4 of the yak and cattle.
AQP4 is a tetrameric protein (Fig. 2III), and each monomer consists of six transmembrane regions with corresponding amino acid sites as shown in the Table 2. The secondary structure of AQP4 protein in both yak and cattle is composed of α-helices, extended strands, β-turns, and random coils, and the proportion of each of these secondary structures is quite distinct between the two species (Table 3). It is noteworthy that the α-helix region of yak AQP4 is composed of 130 amino acids (40.12% of the total length). Moreover, the yak ortholog has eight more amino acid residues than the cattle AQP4, and these residues are largely distributed along the cytoplasmic tail (Fig. 2I).
Analysis of Transmembrane Domain of Yak and Cattle
The Secondary Structure on Aquaporin-4 Protein of Yak and Cattle
Hydrophilicity/hydrophobicity analysis showed that the hydrophobicity of yak AQP4 at position 82 was lower than that in the cattle homolog, while other regions were apparently identical between these species. In other words, Ser82 in yak AQP4 had stronger hydrophobicity than Ala82 in cattle (Fig. 2II).
Expression of AQP4 in yak brain
The levels of AQP4 mRNA and protein in the cerebral cortex, thalamus, caudate nucleus, and hippocampus of both yak and cattle were investigated by RT-qPCR and WB, respectively. As shown in Figure 3, AQP4 protein levels were higher in the cerebral cortex (Fig. 3I, II), but contrarily, AQP4 mRNA levels were lower in this same tissue when compared to other central nervous system compartments in yak (Fig. 3III). In the yak, the highest AQP4 mRNA expression was detected in the thalamus (Fig. 3III). Still, both protein and mRNA levels in the above encephalic regions of the yak were significantly lower than those of cattle (Fig. 3III).
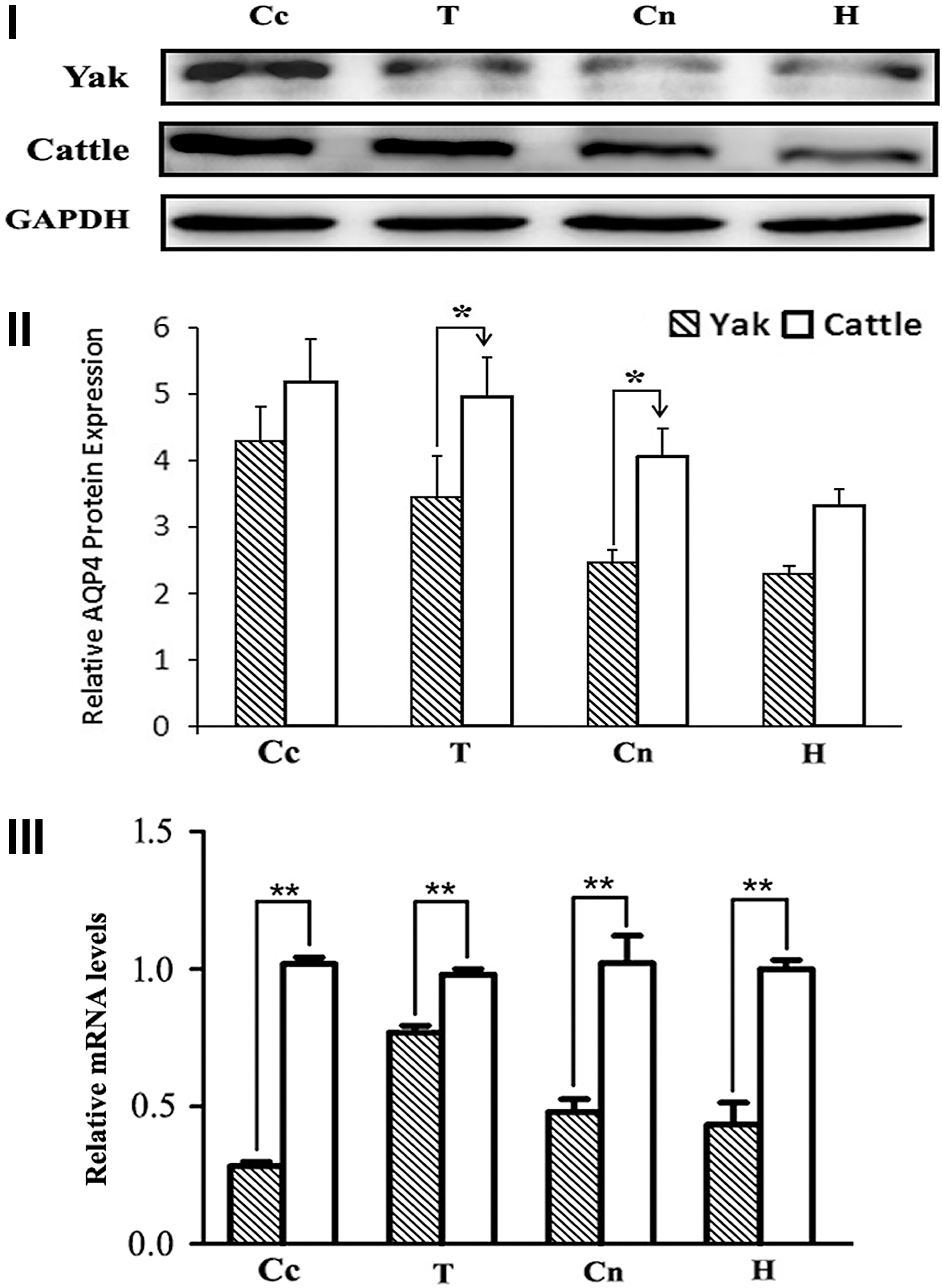
The protein and gene expression of AQP4 in the brain of yak and cattle.
Localization and quantitative analysis of AQP4 in yak brain
To analyze the localization and expression of AQP4 in different encephalic regions of yak and cattle brain, four compartments (i.e., cerebral cortex, thalamus, caudate nucleus, and hippocampus) were selected for further immunohistochemistry analysis. As shown in Figure 4, AQP4 staining was detected in all four regions, but with the highest expression in cerebral cortex. At the cellular level, both yak and cattle AQP4 were mainly detected and largely distributed in respective neurons, capillaries, and glial cells (Fig. 4I). Nevertheless, the IOD levels of AQP4 in the encephalic regions of yak were significantly lower than those of cattle (p < 0.01) (Fig. 4II).
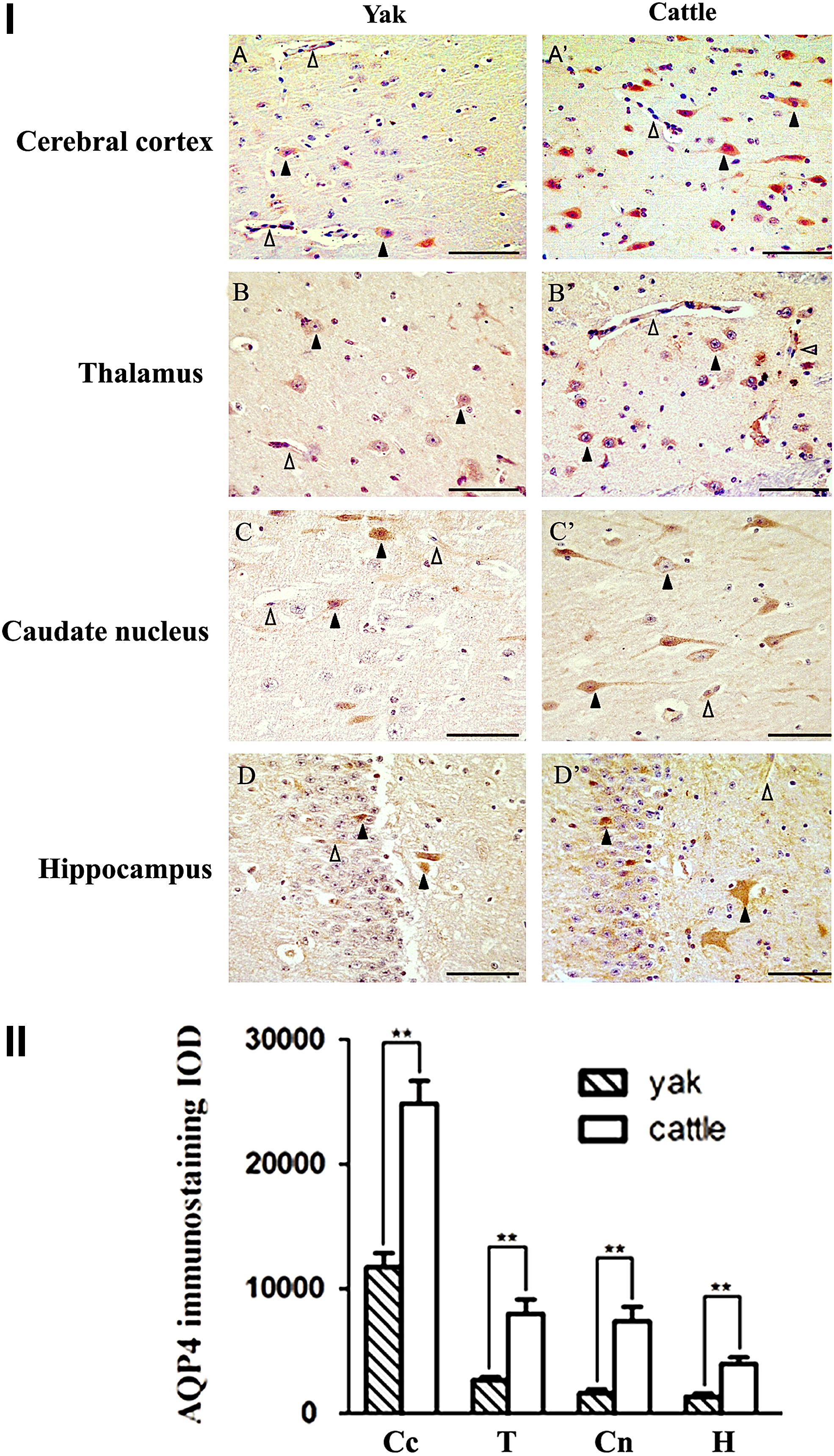
Immunohistochemistry localization of AQP4 in the brain of of yak and cattle.
Ultrastructure characteristic of the yak brain after prolonged ischemia
To test whether yak AQP4 may have a role driving cerebral edema, we further analyzed the effect of prolonged cerebral ischemia on the ultrastructure of yak brain and compared it with the cattle counterpart. As shown in Figure 5A and A′, in prolonged ischemic groups, a vasogenic edema around the cerebral capillaries appeared in both yak (Fig. 5A) and cattle brain (Fig. 5A′). However, the vasogenic edema detected in the cattle brain was apparently more severe (indicated by an asterisk). In the normal group, both yak (Fig. 5B) and cattle (Fig. 5B′) have no vasogenic edema.
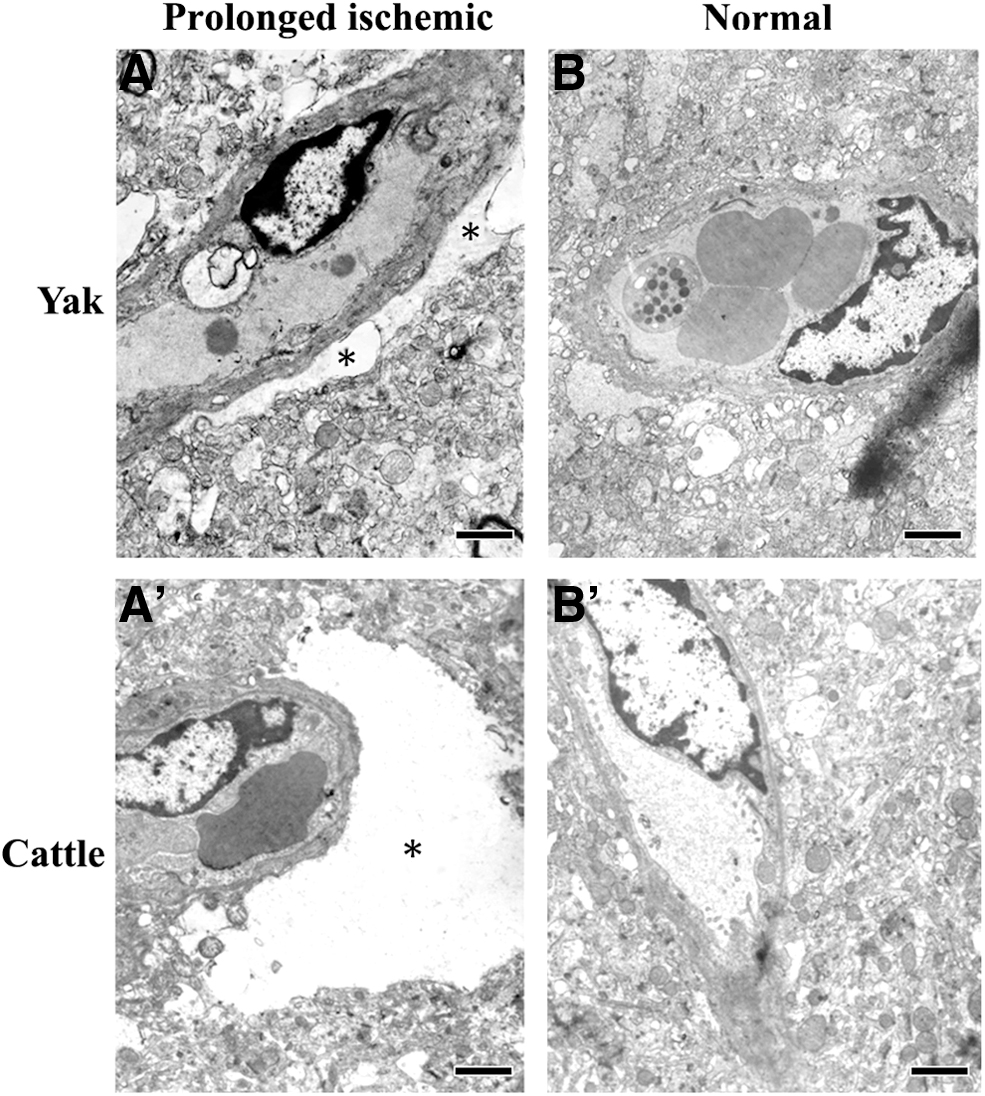
Ultrastructure comparison of the yak and cattle brain in prolonged ischemic group and normal group.
Discussion
To adapt to a high-altitude environment, plateau animals have developed some distinct characteristics. The yak, a typical plateau mammal, has been extensively studied to better understand this level of adaptation. Studies have shown that a higher expression of both HIF-1α and VEGF is possibly linked to the high-altitude adaptation of the yak (Dolt et al., 2007; He et al., 2016). Both HIF-1α mRNA and protein in yak have a tissue-specific expression (Wang et al., 2006). The yak HIF-1α cDNA has been cloned (Dolt et al., 2007), and its broad mRNA expression revealed an evolutionary conservation through multiple sequence alignment, where 15 bases are modified and result in 8 amino acid substitutions in the translated cattle protein (Xiong et al., 2015). As an upstream modulator of AQP4, HIF-1α plays an important role in brain edema (Assentoft et al., 2015). Our group has recently cloned the yak ghrelin and examined its expression in the hypothalamus-pituitary-gastrointestinal tract axis, providing some basis for unique adaptation system of yak to high-altitude environment.
For the first time, in this study, we have isolated yak AQP4 and further characterized its potential role in hypoxia-induced brain edema. Protein sequence analysis as well as expression profiling of AQP4 helped us to initially understand the mechanism of high-altitude hypoxic stress response.
AQP4 is a high-capacity membranous water channel protein that is heterogeneously expressed in brain tissue parenchyma. Compared to other aquaporin proteins, AQP4 is the most abundant in the mammalian brain (Zhang et al., 2019), which shows high expression in fluid transport, including pial and ependymal surfaces in contact with cerebrospinal fluid, subarachnoid space, and the ventricular system (Tamtaji et al., 2019). Based on its location and expression, it was hypothesized that AQP4 is involved in bidirectional fluid exchange between both the blood and cerebrospinal fluid compartments and the brain (Hubbard et al., 2015). Recent studies have reported that brain edema progression may be altered by genetic deletion of AQP4. In fact, edema formation and animal mortality can be reduced after AQP4 deletion in a number of edematous conditions, such as ischemia, water intoxication, acute liver failure, and meningitis (Tamtaji et al., 2019; Zhang et al., 2019). In contrast, AQP4 overexpression in mice glia cells may lead to increased edema formation due to water intoxication (Assentoft et al., 2015). In addition, increased levels of AQP4 mRNA have been detected in numerous conditions, including bacterial meningitis, subarachnoid or intracerebral hemorrhage, brain tumors, and hydrocephalus (Zhang et al., 2019). Altogether, these studies have suggested that AQP4 may play a specific role in cerebral edema.
In this study, we show the expression levels of the yak AQP4 homolog, and how it is compared to those of closely related low-altitude cattle. In our study, immunohistochemical analyses have revealed that the expression of AQP4 is well detected in the cerebral cortex, thalamus, caudate nucleus, and hippocampus, in both yak and cattle. These data are consistent with previous reports (Li et al., 2013). It has been suggested that the role of AQP4 in cerebral cortex, caudate nucleus, and hippocampus is mainly related to the regulation of extracellular volume and potassium concentration (Yang et al., 2011), In particular, AQP4 may act as an osmotic receptor in the thalamus, regulating osmotic pressure and participating in water metabolism throughout the body (Venero and Vizuete, 1999). Furthermore, our results have shown that the levels of yak AQP4 protein and mRNA in cerebral cortex, thalamus, caudate nucleus, and hippocampus were differential and significantly lower than those of cattle. The expression of AQP4 and formation of brain edema appear to be positively correlated, and a high expression of AQP4 may directly lead to the formation of cerebral edema through continuous water accumulation. Our results suggest that low expression of AQP4 plays a critical role in the resistance of cerebral edema, therefore reducing transportation of water in the yak brain.
In ischemic stroke, sudden disruption of blood supply causes cellular metabolic failure, ionic imbalance, and BBB breakage, leading to brain edema (Walberer et al., 2008; Mamtilahun et al., 2019). AQP4 has been suggested to be a critical player in both edema formation and resolution following cerebral ischemia (Tait et al., 2008). Some studies have indicated that ischemic brain injury elevates HIF-1α expression, which leads to upregulation of MMP9, MMP2, and AQP4, resulting in BBB disruption (Assentoft et al., 2015; Mamtilahun et al., 2019). These changes accelerate blood plasma component diffusion by exosmosis to the extracellular space after acute ischemia, causing vasogenic brain edema, and AQP4 would then act as a water “exit route” in the brain edema (Mamtilahun et al., 2019). Interestingly, in this study, when cerebral cortex ultrastructure was analyzed with prolonged cerebral ischemia, the cattle brain showed more severe vasogenic edema than the yak. Our results suggest that AQP4 may be an optimal therapeutic target for the prevention of high-altitude cerebral edema. It is worth mentioning that acetazolamide, a high-altitude tolerance drug, is also an aquaporin blocker, which fits nicely with our findings of AQP4 downregulation in yak (Swenson, 2016).
Besides modulating water transport, AQP4 also has important roles in many other physiological activities. Since both protein and mRNA levels of yak AQP4 were lower than those in cattle, we asked the how this decreased the expression of AQP4 could meet the physiological needs in the yak brain to overcome hypoxic conditions. To figure this out, the coding region of yak AQP4 gene was cloned, and some characteristics of the encoding protein structure were predicted and analyzed in silico. Sequence alignment with a number of mammalian AQP4 orthologs suggested that the amino acid sequence of yak AQP4 is evolutionarily conserved. In fact, the homology of yak AQP4 was over 99%, at the protein level, when compared to the many of the orthologs analyzed in this study. It is noteworthy that there was only one amino acid difference between yak and cattle; the yak AQP4 has the polar Ser82, while the cattle AQP4 has a nonpolar Ala82.
Protein phosphorylation is a major post-translational modification by which cells can promote internalization of particular proteins, affecting trafficking to the plasma membrane and/or directing molecules to certain intracellular degradation pathways (Farr et al., 2019). Recently, a range of phosphorylated residues have been detected by mass spectrometry at the C-terminus of AQP4 (Ser276, Ser285, Ser315, Ser316, Ser321, and Ser322) (Assentoft et al., 2015). Moreover, studies have shown that the function of AQP4 can be inhibited by Serl80 acidification (Han et al., 1998; Moeller et al., 2009; McCoy et al., 2010), while it might be also activated by Ser111 acidification (Zelenina et al., 2002). Therefore, we suggest that the phosphorylation status of Ser82 in the yak AQP4 could modulate the normal physiological function of brain cells under extreme hypoxia. Indeed, previous studies have shown that a higher folding degree of the AQP4 cytoplasmic tail can improve the protein function (Moe et al., 2008; Fenton et al., 2010).
Altogether, in this study, we hypothesize that, during high-altitude hypoxic adaptation, a lower expression of AQP4 may effectively inhibit the occurrence of cerebral edema through reducing water transport, while maintaining its physiological functions by phosphorylation-dependent gating and adjusted folding of AQP4. Our study suggests an important role for AQP4 in adaptation to hypoxia by the plateau yak, in a high-altitude environment. In addition, as a predominant water channel molecule in the mammalian brain, AQP4 plays a critical role in brain edema formation and brain water clearance; it could be an optimal therapeutic target for the prevention and treatment of high-altitude cerebral edema, but more detailed research is needed.
Footnotes
Acknowledgments
We thank Jialong Li, and Yuanzhang Zheng (School of Life Sciences, Lanzhou University, Lanzhou, China) for assistance with sample collection. The authors would like to express their gratitude to EditSprings for the expert linguistic services provided.
Author Disclosure Statement
The authors declare there are no conflicting financial interests.
Funding Information
This work was greatly supported by a grant from the National Natural Science Foundation of China (No. 31760271).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.