
Abstract
Wang, Juan, Jiahui Wang, Xin Li, Wanju Hou, Jie Cao, and Jing Feng. Endothelial dysfunction in a cell culture model exposed to various intermittent hypoxia modes. High Alt Med Biol. 21:388–395, 2020.
Objective:
To construct an in vitro model of endothelial cells exposed to various intermittent hypoxia (IH) modes, and determine whether different frequencies and degrees can cause different effects on endothelial cells.
Methods:
EA.hy926 cells were used to set up the cell model. A program-controlled gas delivery system was designed to regulate the flow of premixed air into the cell culture chamber. The cells were divided into eight groups exposed to various IH modes: standard cell culture group, intermittent normoxia (IN) group (21% O2 15 seconds/21% O2 3 minutes 45 seconds for 12 cycles/h), IH1 group (1.5% O2 15 seconds/21% O2 8 minutes 15 seconds for 6.32 cycles/h), IH2 group (1.5% O2 15 seconds/21% O2 5 minutes 15 seconds for 9.23 cycles/h), IH3 group (1.5% O2 15 seconds/21% O2 3 minutes 45 seconds for 12 cycles/h), IH4 group (1.5% O2 15 seconds/21% O2 1 minute 45 seconds for 20 cycles/h), IH5 group (1.5% O2 15 seconds/21% O2 15 seconds for 40 cycles/h), and IH6 group (10% O2 15 seconds/21% O2 3 minutes 45 seconds for 12 cycles/h).
Results:
Nuclear factor κB (NFκB) p65, c-fos, tumor necrosis factor-alpha (TNFα), malondialdehyde (MDA), and endothelin-1 (ET-1) were higher in the IH3 group or IH6 group than those in IN group, and they were much higher in IH3 group than those in IH6 group. Superoxide dismutase (SOD) and nitric oxide (NO) were the opposite results. In IH1, IH2, IH3, IH4, and IH5 groups, the frequencies increased gradually. NFκB p65, TNFα, and c-fos were the highest in IH3 group. MDA and ET-1 were the highest in IH2 group. SOD and NO were the lowest in IH2 group.
Conclusions:
Different IH frequencies and degrees could cause different effects on endothelial cells. The endothelial responses varied with the duration of reoxygenation. So, the duration of reoxygenation was the key phase for endothelial dysfunction.
Introduction
Obstructive sleep apnea (OSA) is one of the most common public health problems characterized by repetitive episodes of complete or partial obstruction of the upper airway during sleep (Turnbull, 2018). It has been recognized as an independent risk factor for a number of cardiovascular disorders (Uyar and Davutoglu, 2016), particularly such as systemic arterial hypertension, atherosclerosis, coronary artery disease, congestive cardiac failure, and stroke (Song et al., 2015; Mims and Kirsch, 2016; Lombardi et al., 2017). Several mechanisms underlying the association between OSA and cardiovascular diseases have been proposed, including hyperactive sympathetic drive, systemic inflammation, oxidative stress, endothelial dysfunction, and metabolic deregulation (Sforza and Roche, 2016; Javaheri et al., 2017).
Intermittent hypoxia (IH), which is similar with the process of ischemia/reperfusion injury, is the prominent pathophysiologic character of OSA (Dewan et al., 2015). Endothelial dysfunction plays a key role in the development of cardiovascular diseases (Lavie, 2012). Some previous studies have shown the association between IH and endothelial dysfunction (Feng et al., 2012; Badran et al., 2014b). IH can lead to the increased production of reactive oxygen species (ROS) through oxidative stress activation, and ROS can then activate and upregulate the nuclear factor κB (NFκB) pathway (Eisele et al., 2015). The downstream products of NFκB activation, including tumor necrosis factor-alpha (TNFα), interleukin 6 (IL-6), and interleukin 8 (IL-8), have been reported to be implicated in the pathogenesis of endothelial dysfunction (de Lima et al., 2016).
However, the effects of different frequencies and degrees of IH on endothelial cells have not been fully elucidated yet. The aims of this study are to develop an in vitro model of endothelial cells exposed to various IH modes, including different IH frequencies and degrees, and explore whether different frequencies and degrees can cause different effects on endothelial cells.
Materials and Methods
Cell culture
EA.hy926 cells, produced by hybridizing human umbilical vein endothelial cells with the epithelial cell line a549, were used in the IH model. EA.hy926 cells were cultured after resuscitation in the RPMI-1640 medium in 21% O2, 5% CO2, and saturated humidity at 37°C. Once cells reached subconfluence, they were transferred to an antibiotic-free medium for 48 hours and all the following biochemical experiments were performed in a serum-free medium. Before being exposed to IH, cell cultures were regulated to a pH value of 7.4 and 3 × 106 cells/ml on six-well culture plates with each well containing 1 ml culture medium, which overlaid the cell monolayer in a thickness of 3 mm.
IH exposure protocol
A gas control delivering system was designed to regulate the flow of premixed air into a customized chamber housing cell cultures (owned and operated by the Respiratory Department of Tianjin Medical University General Hospital, Copyright: Feng Jing, Tianjin, China, 2005). Briefly, a series of programmable solenoids and flow regulators altered the FIO2 levels by software edited with Visual C++ computer language controlling the numbers and durations of IH cycles (IH and reoxygenation [ROX] protocol, IH/ROX protocol). The chamber was equipped with a humidifier, thermostat, and molecular sieve to maintain the inner temperature of 37°C, humidity of 45%–70%, and germ-free circumstance.
The volume of housing chamber for cell cultures was 1.8 liters, and the gas flow (5.0 l/min) into the chamber needed 30 seconds to substitute inner gas with designed premixed air, changing the gas phase completely and then exposing cell monolayer. For example, when the designed IH/ROX protocol was 1.5% O2 15 seconds/21% O2 3 minutes 45 seconds, the real exposing protocol was 30 seconds, 15 seconds/30 seconds, 3 minutes 45 seconds, namely 1.5% O2 45 seconds/21% O2 4 minutes 15 seconds. This model resulted in cyclic fluctuations of cellular PO2 values during each hypoxic episode as designed. With the model, we could control gas tension and then simulate various IH modes. The actual real-time PO2, PCO2, and pH value levels in the culture medium, 1 mm above the cell layer, were sampled and monitored (n = 5) with fluorescence quenching oximetry, carbon dioxide electrode (Lazar Research), and blood gas analyzer (AVL995, Switzerland). PO2 and PCO2 monitors were used to measure the environmental levels in housing chamber for cell cultures. The grouping and the results of PO2 monitoring are given in Table 1.
Oxygen Tension at the Bottom of a Culture Well in Intermittent Normoxia and Different Frequencies and Degrees of Intermittent Hypoxia
IH, intermittent hypoxia; IN, intermittent normoxia; ROX, reoxygenation; SC, standard cell culture.
Test indicators
The levels of NFκB p65 protein and TNFα were determined in the extracted supernatant with the use of ELISA kits (Cell Signaling Technology and Shenzhen Jing-mei, respectively) according to the kit instructions. The concentrations of superoxide dismutase (SOD), malondialdehyde (MDA), endothelin-1 (ET-1), and nitric oxide (NO) were determined with the use of the chemical reagent method (Nanjing Jiancheng Bioengineering Research Institute) according to the kit instructions. Nuclear extracts were obtained from cells using a nuclear extraction kit according to the manufacturer's instructions (Active Motif). Cell total protein levels were measured simultaneously with the bicinchoninic acid method for standardization (Shenzhen Jing-mei) (n = 12).
Real-time polymerase chain reaction (PCR) was carried out to analyze c-fos mRNA expression levels. Total RNA was extracted using the TRIzol reagent (GIBCO) from exposed endothelial cells. RNA integrity was confirmed by formaldehyde gel electrophoresis. First-strand cDNA was amplified using reverse transcription PCR. The sets of PCR primers used were the following: c-fos, sense primer, 5′-CAGACTACGAGGCGTCATCC-3′, antisense primer, 5′-GTGACCGTGGGAATGAAGTTG-3′, cDNA product, 172 bp; housekeeping gene β-actin, sense primer, 5′-TTCTACAATGAGCTGCGTG-3′, antisense primer, 5′-CACAGCCTGGATAGCAAC-3′, cDNA product, 150 bp. Relative gene expression changes were determined using the 2−△△Ct method.
Statistical analysis
SPSS 18.0 (SPSS, Chicago, IL) software package was used for statistical analysis and illustration. One-way analysis of variance was performed for whole difference among groups, and Bonferroni post hoc multiple comparisons were used to evaluate differences between internal groups. Unless otherwise stated, values were reported as mean ± SD, and p-value of <0.05 was considered statistically significant.
Results
Comparisons among NFκB, c-fos, and TNFα
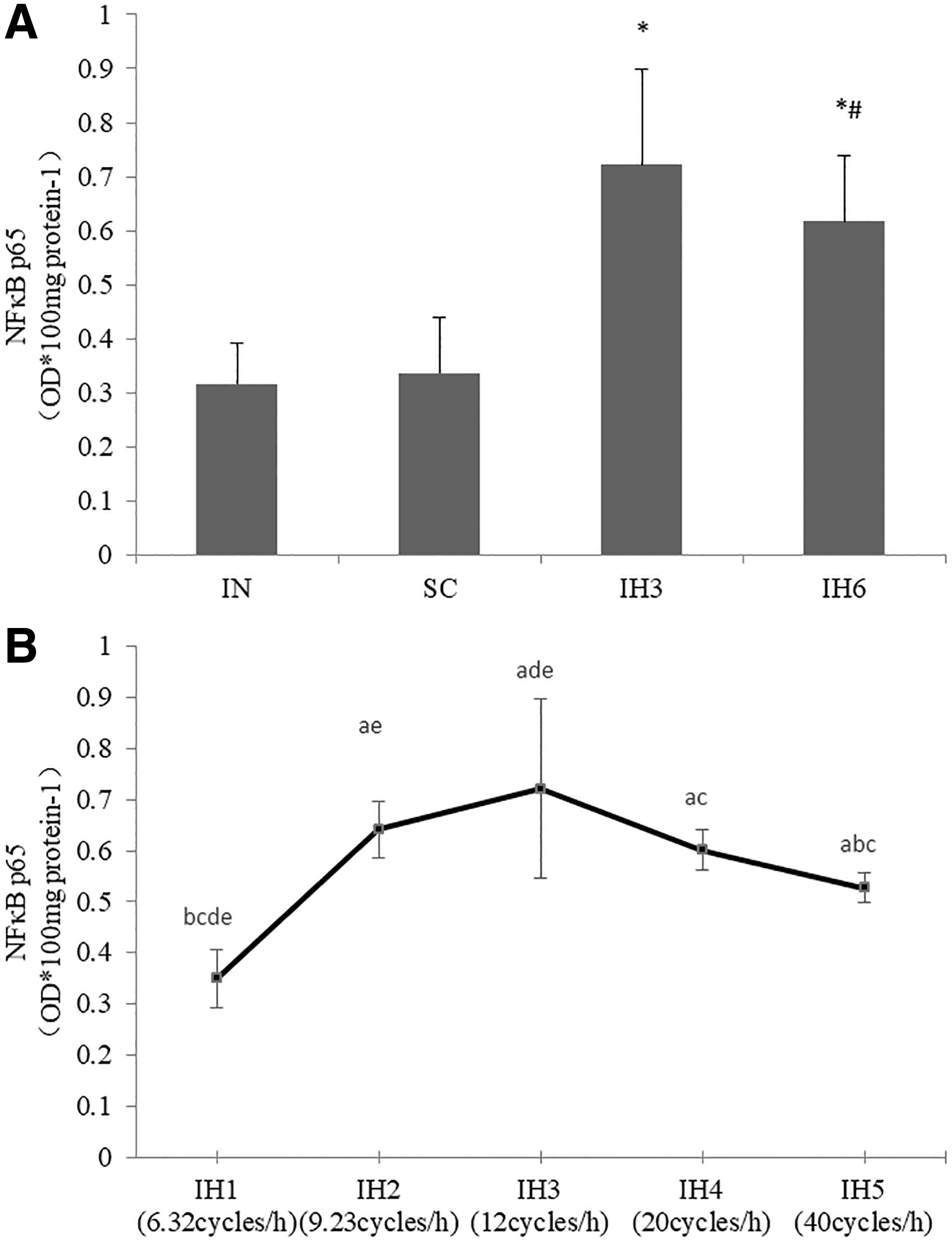
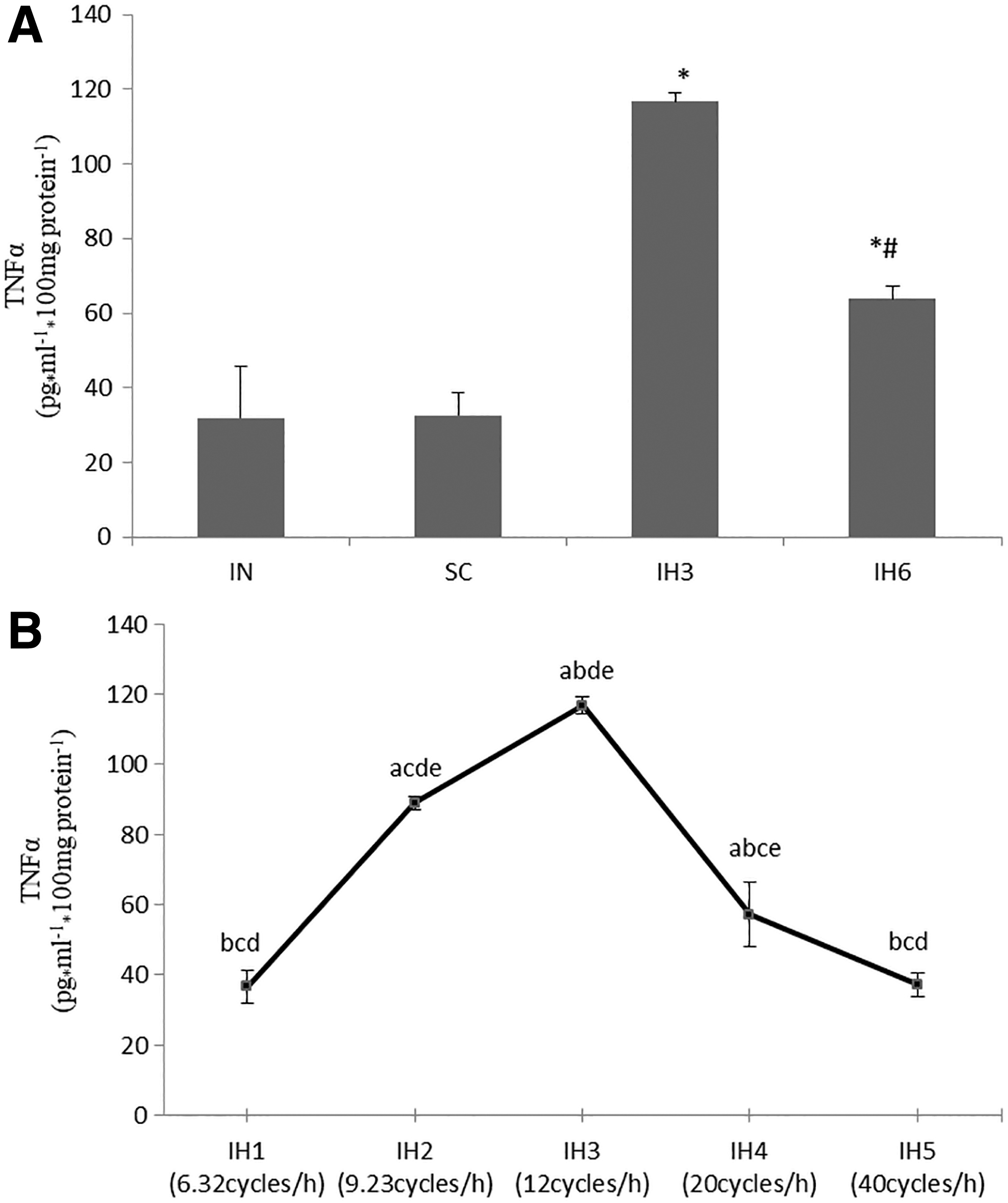
Compared with the intermittent normoxia (IN) group, the levels of NFκB p65 protein, c-fos mRNA, and TNFα were higher in IH3 and IH6 group (p < 0.05). Also, they were much higher in IH3 group than in IH6 group (p < 0.05). There was no statistical difference when comparing IN group and standard cell culture (SC) group. The levels of NFκB p65 protein, c-fos mRNA, and TNFα were measured in IH1, IH2, IH3, IH4, and IH5groups. These groups had different IH frequencies and the frequencies were increased gradually from IH1 to IH5. The levels of NFκB p65 protein, c-fos mRNA, and TNFα increased from IH1 to IH3 group, and then decreased in IH4 and IH5 groups. There was a statistical difference in these groups (p < 0.05). The levels of NFκB p65 protein, c-fos mRNA, and TNFα in IH3 group were the highest than the other groups (p < 0.05) (Figs. 1–3).

Comparisons between SOD and MDA
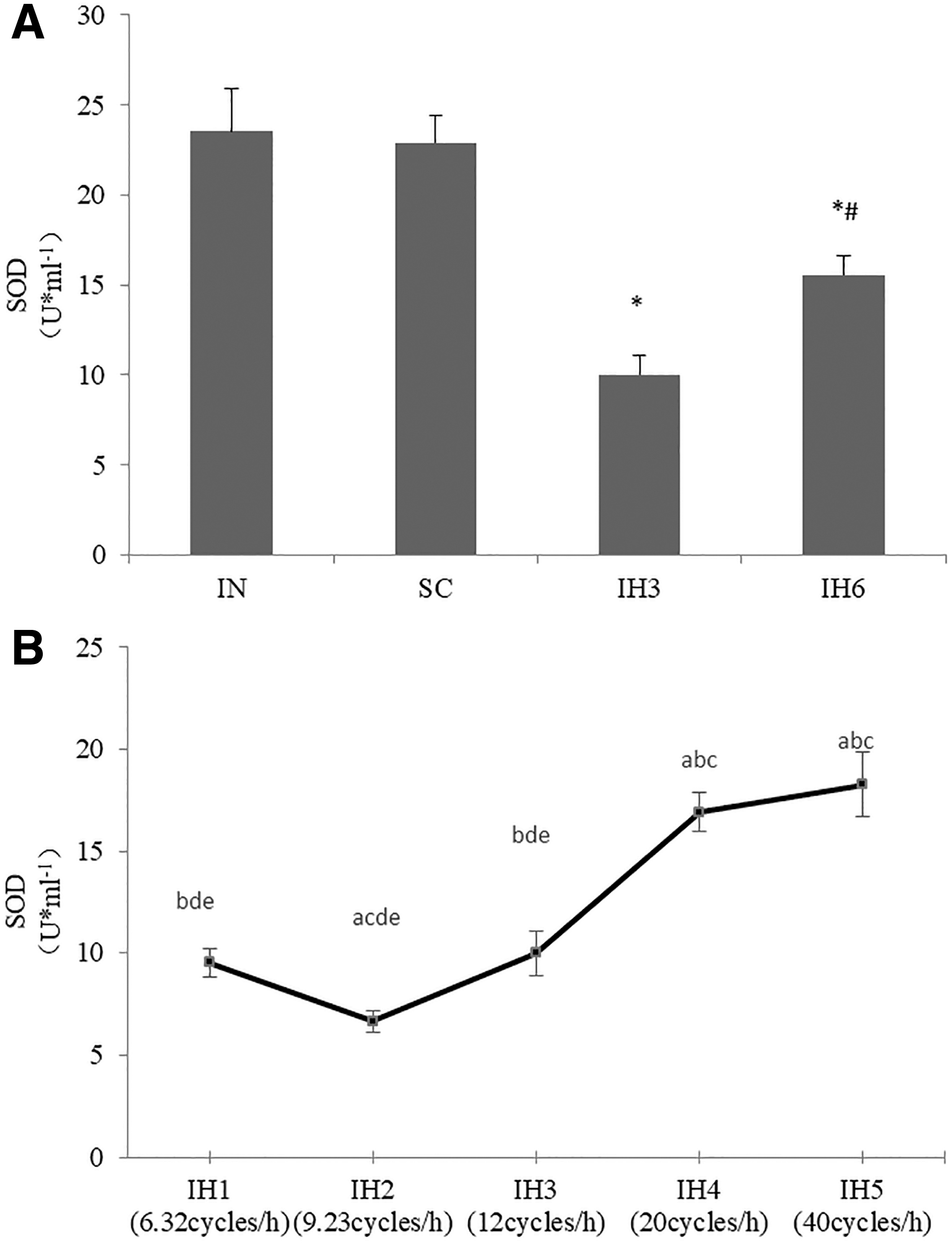
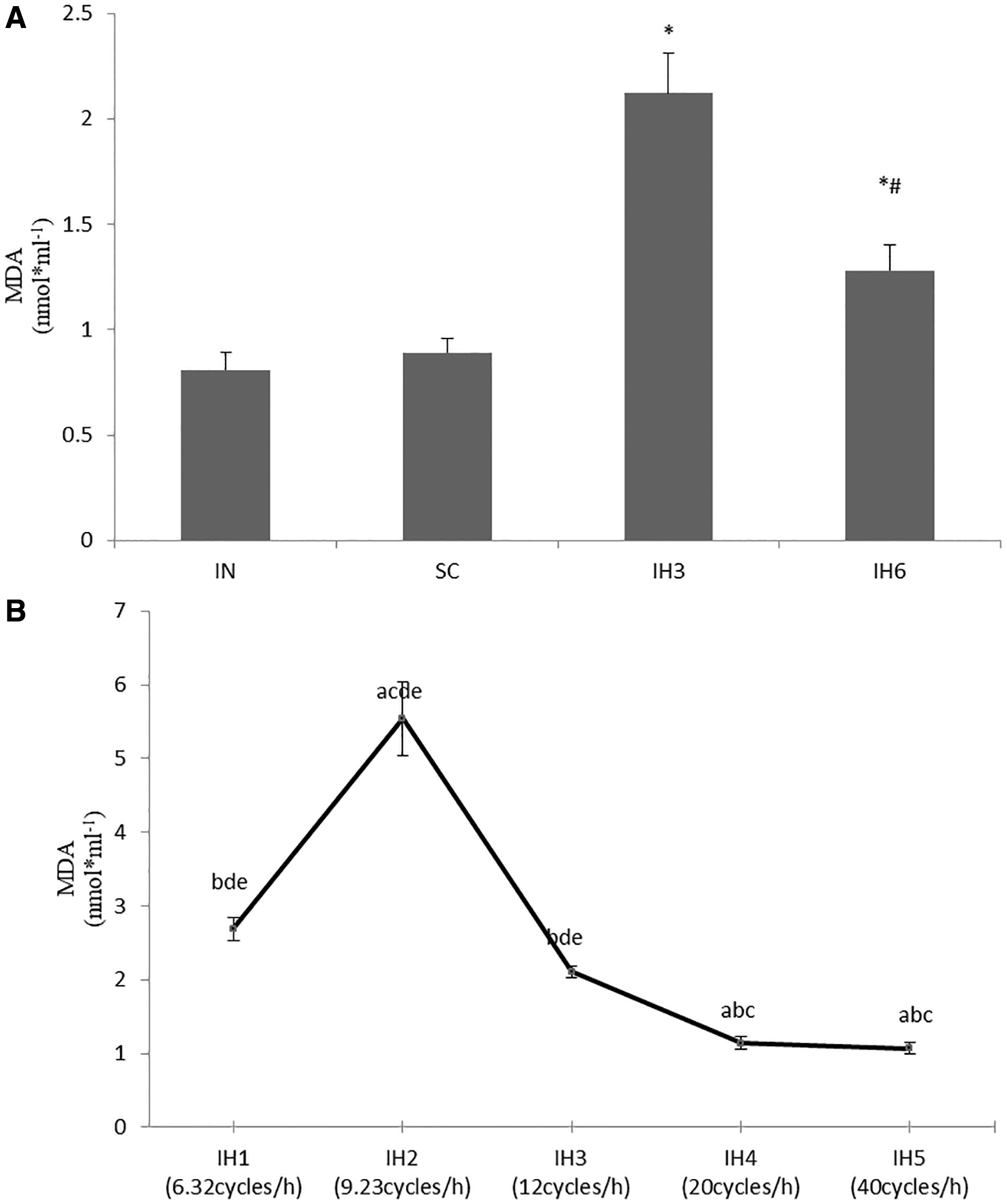
The concentrations of SOD were significantly decreased, and MDA was significantly increased in IH3 and IH6 groups than in IN group (p < 0.05). Also, the concentrations of SOD were decreased more obviously in IH3 group than in IH6 group (p < 0.05). The concentrations of MDA were increased more obviously in IH3 group than in IH6 group (p < 0.05). There was no statistical difference when comparing the IN group and SC group. We examined the concentrations of SOD and MDA in groups with different IH frequencies and there were statistical differences in IH1, IH2, IH3, IH4, and IH5groups (p < 0.05). The level of SOD in IH2 group was the lowest among these groups (p < 0.05), and the level of MDA in IH2 group was the highest among these groups (p < 0.05) (Figs. 4 and 5).
Comparisons between ET-1 and NO
The concentrations of NO was significantly decreased, and ET-1 was significantly increased in IH3 and IH6 groups than in the IN group (p < 0.05). Also, the concentrations of NO were decreased more obviously in IH3 group than in IH6 group (p < 0.05). The concentrations of ET-1 were increased more obviously in IH3 group than in IH6 group (p < 0.05). There was no statistical difference when comparing the IN group and SC group. We examined the concentrations of NO and ET-1 in groups with different IH frequencies and there were statistical differences in IH1, IH2, IH3, IH4, and IH5groups (p < 0.05). The level of NO in IH2 group was the lowest among these groups (p < 0.05), and the level of ET-1 in IH2 group was the highest among these groups (p < 0.05) (Figs. 6 and 7).


Discussion
IH is an important pathophysiologic mechanism and the underlying basis for cardiovascular diseases in patients with obstructive sleep apnea syndrome (OSAS) (Turnbull, 2018). Endothelial dysfunction caused by IH can occur through mechanisms, including oxidative stress and activation of inflammation, based on the evidence from animal models and endothelial cell studies (Ryan et al., 2009; Dumitrascu et al., 2013). It has been recognized that endothelial dysfunction plays a crucial role in OSA-related cardiovascular diseases (Badran et al., 2014a). Some clinical and animal studies have demonstrated that endothelial dysfunction is a precursor of atherosclerosis, and that atherosclerosis is a pathological condition underlying cardiovascular diseases (Song et al., 2015, 2018). The IH mode consists of different IH frequencies and degree patterns. It has been suggested that the frequency and magnitude of hypoxia cycling in IH markedly alter wound healing responses and emerge as key factors determining how cells will respond in OSA (Campillo et al., 2017). In our study, we developed an in vitro IH model that involved exposing EA.hy926 cell cultures to different frequencies and degree IH cycles, simulating various IH modes. The aim of our study is to explore whether different frequencies and degrees can cause different effects on endothelial cells. In our study, the cells were divided into eight groups exposed to various IH modes. In IH3 and IH6 groups, the endothelial cells were exposed to different oxygen concentration levels (1.5% and 10%, respectively) at the same frequency. In IH1, IH2, IH3, IH4, and IH5 groups, IH protocols maintained the same duration of hypoxic episodes and the same oxygen concentration level, while the different durations of ROX made the frequency different. In IH1, IH2, IH3, IH4, and IH5 groups, with ROX durations shortening, the frequencies increased gradually.
NFκB is an important transcriptional regulator of inflammatory mediators. IH could activate subunits of the NFκB transcription factor family, such as p50 and p65, responsible for the activation of inflammatory genes, including IL-6 or TNFα (Taylor et al., 2014). Ryan et al. (2005) reported increased NFκB activity in monocytes derived from OSA patients and this effect was associated with elevated serum TNFα. Recoquillon et al. (2017) reported that IH reproduced in vitro the features of endothelial dysfunction and inflammation observed in OSA, notably the increasing of ROS, the activation of p65-NFκB, and the release of inflammatory cytokines. In vascular endothelial cells, cytokines induce genes that are expressed in classic inflammatory injury partly through the activation of NFκB, the ideal molecular index for inflammation (Toffoli et al., 2009). Once activated, NFκB can bind to promoters of specific genes initiating transcription of many products such as TNFα, adhesion molecules, and acute-phase proteins. These products are all involved in the process of endothelial dysfunction and cardiovascular diseases (Badran et al., 2014a). C-fos is a constituent part of the activator protein complex-1 (AP-1), which is an inflammatory transcription factor. The c-fos mRNA is one of the most extensively studied members of the immediate early gene family (Dampney and Horiuchi, 2003). C-fos and NFκB collaborate to modulate multiple inflammatory responses (Adcock, 1997). Previous studies suggested that the immediate early gene c-fos played an important role in cellular adaptations to hypoxia (Rybnikova et al., 2005). Hypoxia induced c-fos expression upregulation both in animals and in cell cultures (Tsai et al., 2011; Yadav et al., 2017). The study in cell culture (Yuan et al., 2004) showed that hypoxia-induced c-fos expression contributed to AP-1 transcription factor activity and mediated AP-1-regulated downstream genes. This response could cause a series of pathophysiological reactions and plays a role in endothelial cell injury and repair. So, in our study, NFκB p65, TNFα, and mRNA expressions of c-fos were measured to explore the inflammatory response in the endothelial cells exposed to various IH modes. Our data showed that NFκB p65, TNFα, and mRNA expressions of c-fos were higher in IH3 group or IH6 group than those in IN group, and they were much higher in IH3 group than those in IH6 group. So, to a certain extent, we considered that IH induced the inflammatory responses of endothelial cell, which could be dependent on the degree of IH. Also our data showed that NFκB p65, TNFα, and mRNA expressions of c-fos did not increase gradually with the increase of frequency; they all increased from IH1 to IH3 group, and then decreased gradually in IH4 and IH5 groups. The inflammatory responses of endothelial cell caused by IH could be related to the frequency of IH.
Oxidative stress is a pathological state in which excessive production of oxygen free radicals and/or damage to the cellular antioxidant defense system lead to excessive accumulation of oxygen free radicals and their related metabolites, thus causing damage to the organism. Oxidative stress injury is a manifestation of the imbalance of oxidation and antioxidant capacity. ROS and its products activate sensitivity signaling pathways and transcription factors such as NFκB, in turn upregulating the expression of relevant inflammatory factor genes. The production and release of a large number of cytokines and adhesion molecules form an inflammatory cascade effect, leading to endothelial dysfunction. As a lipid oxidation end product, MDA causes tissue damage by affecting mitochondrial respiratory chain complex and key enzyme activities, and is used as an indicator of oxidative damage in the body. SOD is an important oxygen free radical scavenger, and SOD catalyzes oxygen free radical to be hydrogen peroxide and oxygen, thus scavenging excess oxygen free radical in the body, which is regarded as the antioxidant damage index of organisms (Zhao et al., 2016; Chiş et al., 2018). So, in our study, SOD and MDA were measured to explore the oxidative stress response in the endothelial cells exposed to various IH modes. Our data showed that SOD was decreased in IH3 group or IH6 group than that in IN group, and it was much lower in IH3 group than that in IH6 group. The result of MDA was the opposite. MDA was increased in IH3 group or IH6 group than that in IN group, and it was much higher in IH3 group than that in IH6 group. So, we considered that IH induced the oxidative stress responses of endothelial cell, which could be dependent on the degree of IH. Also, our data showed that SOD and MDA did not decrease or increase gradually with the change of frequency, SOD was the lowest in IH2 group and MDA was the highest in IH2 group. The oxidative stress responses of endothelial cell caused by IH could be related to the frequency of IH.
Endothelial cells exert their physiological functions by secreting vasoactive substances to regulate vascular tension and permeability. Vascular endothelial dysfunction is manifested as imbalance of vasoactive substances secreted by endothelial cells, including NO and endothelin (ET). ET is a vasoconstrictive active polypeptide, in which ET-1 is mainly expressed in endothelial cells, and NO is one of the main components of endodermal diastolic factor. OSAS can initiate oxidative stress response, releasing the imbalance of NO and ET, reducing NO level, and increasing ET secretion, leading to endothelial dysfunction, thus promoting the occurrence and development of cardiovascular diseases such as hypertension and atherosclerosis (Manukhina et al., 2006; Wang et al., 2017). In our study, ET-1 and NO were measured to explore endothelial dysfunction exposed to various IH modes. Our data showed that NO was decreased in IH3 group or IH6 group than that in IN group, and it was much lower in IH3 group than that in IH6 group. ET-1 was increased in IH3 group or IH6 group than that in IN group, and it was much higher in IH3 group than that in IH6 group. So, we considered that IH induced imbalance of vasoactive substances secreted by endothelial cells, which could be dependent on the degree of IH. Also, our data showed that NO was the lowest in IH2 group and ET-1 was the highest in IH2 group. The imbalance of vasoactive substances secreted by endothelial cells could be related to the frequency of IH.
Our data showed that the inflammatory responses, oxidative stress, and imbalance of vasoactive substances caused by IH should be related to the frequency and degree of IH. We have previously shown that serum concentrations of proinflammatory factors, including TNFα, IL-6, IL-8, and intercellular adhesion molecule-1 (ICAM-1), were increased in rats exposed to IH and were dependent on the time and level of IH (Li et al., 2011). Wang et al. (2013) suggested that NFκB mediating endothelial dysfunction could be an important mechanism linking OSA to cardiovascular pathologies and could have an exposure time-dependent effect. Some evidence (Dewan et al., 2015) has shown that the major difference between the short intermittent high-frequency hypoxemia, as seen in OSA, and sustained prolonged low-frequency hypoxemia was the cycles of reoxygenation. IH/ROX is similar to ischemia/reperfusion. Rapid ROX at the end of apneas/hypopneas led to the production of free radicals, resulting in oxidative stress and upregulation of NFκB (Javaheri et al., 2017). Endothelial dysfunction needs enough ROX duration; however, it is not that the longer the ROX duration sustained, the more severe the damage that has occurred. When the IH frequencies were further higher, the durations of ROX might be too short, and the endothelial dysfunction went slighter. However, when the durations of ROX were too long, cell repair started and the endothelial dysfunction decreased.
Conclusion
In conclusion, we have detected the inflammatory responses, oxidative stress, and imbalance of vasoactive substances following IH exposure in endothelial cells. The effects are not only dependent on the degree of IH but also the frequency of IH and need enough ROX duration. The duration of ROX is the key phase for endothelial dysfunction. These results validate that IH exposure can lead to inflammatory responses, oxidative stress, and imbalance of vasoactive substances of endothelial cell, and different IH frequencies and degrees may cause different effects on endothelial cells.
Footnotes
Authors' Contributions
Designed the experiments: J.C. and J.F.; performed the experiments: J.W., J.W., and X.L.; analyzed the data: W.H.; drafted or revised the article: J.W. and J.F. All coauthors have reviewed and approved of the article before submission.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.