
Abstract
Ji, Weizhong, Yaqing Zhang, Ri-li Ge, Yaqi Wan, and Jie Liu. NMDA receptor-mediated excitotoxicity is involved in neuronal apoptosis and cognitive impairment induced by chronic hypobaric hypoxia exposure at high altitude. High Alt Med Biol. 22:45–57, 2021.
Aim:
Exposure to chronic hypobaric hypoxia at high altitude can lead to cognitive impairment; however, its underlying mechanism is still unclear. Excessive expression of glutamate and its receptors can induce excitotoxicity and cause neuronal necrosis, which is an important causative factor for the occurrence of various diseases in the nervous system. Therefore, excitotoxicity may also occur after exposure to a chronic hypobaric hypoxic environment. This study concentrates on the action mechanism of glutamate and its receptor-mediated excitotoxicity in cognitive impairment, induced by chronic hypobaric hypoxic exposure.
Methods:
Sprague-Dawley rats were fed at regions with three different altitudes (4,300, 2,260, and 450 m) for 8 weeks, and had their behavioral changes assessed by the Morris water maze test. Morphological, molecular biological, and biochemical tests were used to determine the role of N-methyl-
Results:
We found that exposure to chronic hypobaric hypoxia at high altitudes could cause cognitive impairment, damage the neurons in the hippocampus and cortex, increase apoptosis, and lead to abnormal Caspase-3 protein expression. The expression of NMDA and a-amino-3-hydroxyl-5-methyl-4-isoxazole acid (AMPA) receptors increased significantly, as did the levels of oxidative stress and free radicals as well. However, no change in cognitive function was observed in the chronic hypobaric hypoxia environment at the middle altitude, there were no statistically significant differences in neuronal apoptosis and related protein expression compared with the rats in the flatland environment group.
Conclusion:
We show that high-altitude chronic hypobaric hypoxic environment could cause obvious cognitive impairments, which is related to the excitotoxicity mediated by glutamate and its receptors, in contrast to the chronic hypobaric hypoxia environment at middle altitude group and flatland environment group.
Introduction
Hypobaric hypoxia is the most primary feature of the plateau environment and has a complex impact on the human's physiological functions. In recent years, the impact of acute and chronic exposure to hypobaric hypoxia on brain health has attracted widespread attention. At present, the impact of acute hypoxia exposure on cognitive function is well known. However, few studies have been focused on the cognitive impairment induced by chronic hypobaric hypoxia exposure (over 1 month). Whether there is any effect on the cognitive function, is also controversial. For the population residing at different altitudes (500, 2,500, and 3,700 m), the response speed and the cerebral blood flow velocity of the population with different ages living at the middle and high altitudes, were significantly decreased (Hogan et al., 2010). A study showed an increased prevalence of mild cognitive impairment (MCI) in populations with a longer stay in high-altitude areas, which was different from the MCI caused by Alzheimer's disease in neurophysiology (Hota et al., 2012). However, another study found no significant changes in the cognitive function after living at an altitude of 2,260 m for 7 months, suggesting that the hypobaric hypoxic environment at middle altitude did not affect cognitive function (Zhang et al., 2011). The reason for the contradiction between these two findings was the difference in the altitude and the time of exposure. A report on the cognitive function of 28 long-term residents in the high-altitude area whose residence time was >18 years showed that subjects presented a longer reaction time and lower accuracy in the behavioral performance (Yan et al., 2011). At present, there have been relatively few studies on the lowest altitude and related exposure time of the cognitive impairment caused by chronic hypobaric hypoxia exposure, and the results that do exist are inconsistent. In general, cognitive impairment caused by exposure to the chronic hypobaric hypoxic environment mainly include delayed cognitive reaction time, decrease in attention, executive ability decline, and working memory loss, which is consistent with the cognitive impairment range of chronic hypoxia induced by nocturnal apnea (Wang et al., 2015). The degree of cognitive impairment is positively correlated with altitude and exposure time. Attention should be paid to the evaluation method of working memory, as it may have a certain effect on the results of cognitive impairment (Kent, 2016). Researchers in the United States compared the shooting performance of soldiers exposed to a hypobaric hypoxic environment at an altitude of 4,300 m for 30 days and found that their shooting performance decreased significantly compared with that at plains (Kryskow et al., 2013; Roach et al., 2014). Some scholars performed the event-related potentials test in a long-term exposure population, and the results indicated that long-term hypobaric hypoxia environment could affect people's attention and conflict control ability (Ma et al., 2015).
At present, the mechanism of cognitive impairment induced by chronic hypobaric hypoxia remains unclear. Long-term high-altitude hypobaric hypoxic environment leads to the anoxic condition of the brain tissue, which consumes a large quantity of oxygen, and induces neuronal damage. After exposure to hypoxia or ischemia, the release of excitatory and inhibitory synaptic transmitters was increased in neurons, especially the activation of the glutamate-gated channels (Revah et al., 2016). Subsequently, neurons in the cortical tissue undergo a great amount of anoxic depolarization causing excitability, neuronal damage, and even brain cell death. The longer the hypoxia time the more obvious the neuronal damage. Oxidative stress-induced neuronal injury induced by long-term hypobaric hypoxia may also be one of the main reasons for cognitive impairment from chronic hypobaric hypoxia exposure. A study showed that the content of hypoxia-inducible factor (HIF-1 alpha) and nitric oxide was significantly increased in the hypoxia state, resulting in the production of peroxides and other cytotoxic metabolites that damage the brain cells (Barhwal et al., 2009). At the same time, antioxidants acetyl-
Glutamate is an excitatory neurotransmitter and its ionic receptors mainly include the N-methyl-
In this study, field experiments were carried out in environments with different gradients of altitude, thereby establishing the chronic hypoxia rat models. The cognitive function of rats was assessed using Morris water maze, and rat brain slices were prepared for further analysis. The morphological and ultrastructural changes of neurons after chronic hypoxic cognitive impairment were investigated. At the same time, both the messenger RNA (mRNA) and protein expression levels of glutamate and its receptor protein were explored. We further clarified the presence of excitotoxicity, such as lipid peroxidation, calcium overload, and excessive production of free radicals.
Materials and Methods
All animal experiments were approved by the Ethics Committee of Qinghai University. The male Sprague-Dawley rats (90 individuals, 6 weeks old) were provided by the Beijing Charles River Co., Ltd, and were randomly divided into three groups: the high-altitude hypoxia (HH) group, mid-altitude hypoxia (MH) group, and plain control (PC) group, with 30 rats in each group. They were reared continuously for 8 weeks in the Maduo County Qinghai province (altitude 4,300 m), Xining, Qinghai province (altitude 2,260 m), and Xi'an, Shaanxi province (altitude 450 m). The feeding conditions of the animal housing in three different altitudes were the same, the temperature of the animal house was controlled at 18°–22°C, the light/dark time was 12:12 hours, and enough food and water were guaranteed every day.
The Morris water maze experiment
Morris water maze test facility was provided by the Shanghai Xinruan InfoTech Ltd, with a diameter of 1.6 m, a height of 0.6 m, and a water depth of 0.31 m. Four quadrants were set up with special software and they were termed as the first, second, third, and fourth quadrant, respectively. The platform was placed in the third quadrant, which was 0.01 m below the surface. Each rat was put into water daily from different quadrants. The time and the movement trace of rats from swimming to climbing up the platform were recorded. If the rats could not find the platform within 60 seconds, they were manually guided to the platform. For training, rats could only be placed from each quadrant once and the training was performed four times a day, for four consecutive days. This experiment tested the learning and spatial memory ability of rats. On the fifth day, the platform was removed and rats were placed into the water from the first quadrant. The timing was recorded for 60 seconds. The number of times the rat crossed the quadrant of the platform and the percentage of swimming time crossing the target quadrant in the total swimming time were recorded. This experiment tested the memory, retrieval, and application ability of the rats.
Rat cerebral perfusion and sampling
After the water maze test, rats in each group were given an intraperitoneal injection of 10% chloral hydrate for deep anesthesia (1.5 ml/100 g body weight). Twelve rats in each group were randomly selected and fixed on the operating table. The abdominal cavity was cut open and the heart was carefully exposed. The perfusion puncture needle was punctured from the apex of the heart, passing through the left ventricle to the aortic root. After fixing with hemostatic forceps, intraperitoneal infusion of normal saline and 4% paraformaldehyde solution was carried out. After successful perfusion, the brain was removed from the head, and the coronal slices between the optic nerve and the superior cerebellar margin were taken. The 4 mm thick slices were placed and blocked in a 4% paraformaldehyde solution overnight. The fixed tissue blocks were dehydrated, embedded in paraffin, and stored for staining the next day.
Morphological changes in chronic hypoxia rats
Nissl staining
Slides were treated to prevent deactivation and the brain slices were routinely dewaxed in water, placed in 1% toluidine blue water solution preheated at 50°C, and dyed for 20 minutes at 56°C in an incubator. This was followed by distilled water washing, 70% alcohol soaking for 1 minute, and 95% alcohol differentiation. The microscope was controlled till Nissl bodies were clear. Anhydrous alcohol was used for rapid dehydration, xylene for transparency, and neutral gum for fixation. For each slice, three visual fields were randomly selected for observation and counting, images with 400 × were collected and saved, and the final results were averaged.
TUNNEL staining
Routine sections in paraffin blocks were routinely dewaxed in water. Trypsin K working fluid was used for treatment at 37°C for 25 minutes, followed by repeated rinses, alcohol dehydration, xylene transparency, neutral gum seal sealing, and finally a microscopic examination.
Electron microscope special staining and observation
After anesthesia, six rats were taken from each group for decapitation and brain collection. The cortex and hippocampus (HC) were stripped rapidly on ice. They were then fixed in 3% glutaraldehyde electron microscope stationary liquid and preserved in dark. The next day, they were dehydrated by gradient alcohol, then embedded with epoxy resin, sheared by semi-membrane, and stained by lead citrate. Finally, the ultrastructure of neurons in the cortex and HC was observed using a transmission electron microscope.
Immunofluorescence detection of NEUN and Caspase-3
The fixed tissues were dehydrated by automatic dehydrator and then embedded. After slicing, they were dewaxed and rinsed repeatedly followed by the addition of primary antibody (NEUN, mouse monoclonal antibody, concentration: 1:100; Caspase-3, rabbit polyclonal antibody, concentration: 1:100), biotinylated secondary antibody: rhodamine-labeled goat anti-mouse immunoglobulin G, and DAPI; they were incubated for 10 minutes at room temperature. After washing, the anti-fluorescence attenuation patch-sealing agent was used for sealing followed by microscopic examination.
Real-time fluorescence quantitative polymerase chain reaction
The total RNA was extracted from the rat HC and cortex using the FOREGENE kit. The reverse transcription was then carried out. In the 20 μl reverse transcription system, 4 μl 2 × PrimeScript Buffer 2, 1 μl PrimeScript RT Enzyme Mix l, 1 μl RT Primer Mix, 10 μl RNA, and 4 μl RNase Free dH2O were added. Twenty microliters polymerase chain reaction (PCR) reaction system contained 10 μl 2 × Real PCR EasyTM Mix-SYBR, 0.8 μl Forward Primer (10 μM), 0.8 μl Reverse Primer (10 μM), 2.0 μl Template (DNA), and 6.4 μl ddH2O. Reaction conditions were 95°C, 30 seconds; 95°C, 5 seconds; 55°C, 30 seconds; 72°C, 30 seconds; 45 cycles. The primer sequences are given in Table 1, and the β-actin was used as an internal reference. Thermo Scientific PikoReal software (Thermo) was used to analyze the threshold cycle (CT) values of each test sample obtained from PCR. The relative expression level of gene toward mRNA was calculated using the 2-▵▵CT method (Table 1).
Primers and Base Sequences for Real-Time Fluorescence Quantitative Polymerase Chain Reaction Detection
Western blot
BCA kit was used to quantify the total protein (TP) levels of Caspase-3, and the NMDA and AMPA receptors. After 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis separation, they were transferred to the polyvinylidene fluoride membrane. Five percent skim milk powder was used for blocking at room temperature for 1 hour, followed by a rinse for 5 minutes. Rabbit antihuman primary antibody with a 1:1,000 dilution ratio was added and incubated for 1.5 hours at room temperature, then kept at 4°C overnight. Tris-Hcl, NaCl, and Tween20 (TBST) solution was used for rinsing. Horseradish peroxide-labeled goat anti-rabbit secondary antibody solution was added for incubation at room temperature for 1 hour. TBST was rinsed and the luminescent liquid was added for lighting, followed by X light film exposure, development, and fixing. The internal reference used was the GAPDH protein.
Biochemical detection
The TP assay kit, glutathione reductase (GR) kit, hydroxyl free radical test kit, and lipid peroxide assay kit were provided by the Nanjing Jiancheng Bioengineering Institute. The HC and cortex tissues were homogenized in 0.15 M KCl to obtain 10% homogenates. The crude homogenate was centrifuged at 10,000 rpm for 10 minutes. The supernatant was used to estimate the GR, glutathione peroxidase activity, lipid peroxide content, and free radical levels. The temperature for sample preparation was kept at 48°C. The TP content was determined by the Coomassie brilliant blue method and the content of each sample was determined using ultraviolet spectrophotometry.
Statistical methods
All data are expressed as mean ± standard deviation. The water maze test groups were compared using a single-factor analysis of variance (ANOVA). The percentage of apoptosis was measured by the Image-Pro Plus 6.0 image analysis system, and the integrated optical density and area of all images were collected to calculate the percentage of positive expression (%). Thermo Scientific PikoReal software (Thermo) was used to analyze the CT values of each test sample generated in the PCR process. The relative expression level of gene toward mRNA was calculated using the 2-▵▵CT method. The variance analysis among groups was performed using the one-way ANOVA, and the least significant difference test was used to test the homogeneity of variances. The heterogeneity of variance was analyzed by Tamhane's T2. The value p < 0.05 indicated the significant difference among the groups. SPSS 22.0 statistical software was used for analysis.
Results
Behavioral changes in chronic hypoxic rats
Water maze test indexes of each group showed that on the first and second day of training, the average escape latency of each group was slightly different; however, without any statistical significance (p > 0.05). From the training state observation, the limb movement ability and daily activity ability of rats were not affected. On the third and fourth day, the average latency of the HH group was significantly longer than that of the MH and PC group (p < 0.05). On the fifth day, the space exploration experiment showed that the number of crossing platform times and crossing target quadrant time in the HH group were significantly lower than those in the MH and PC group (p < 0.05). However, there was no significant difference in the three indexes between the MH altitude group and the PC group (p > 0.05), as given in Figure 1.
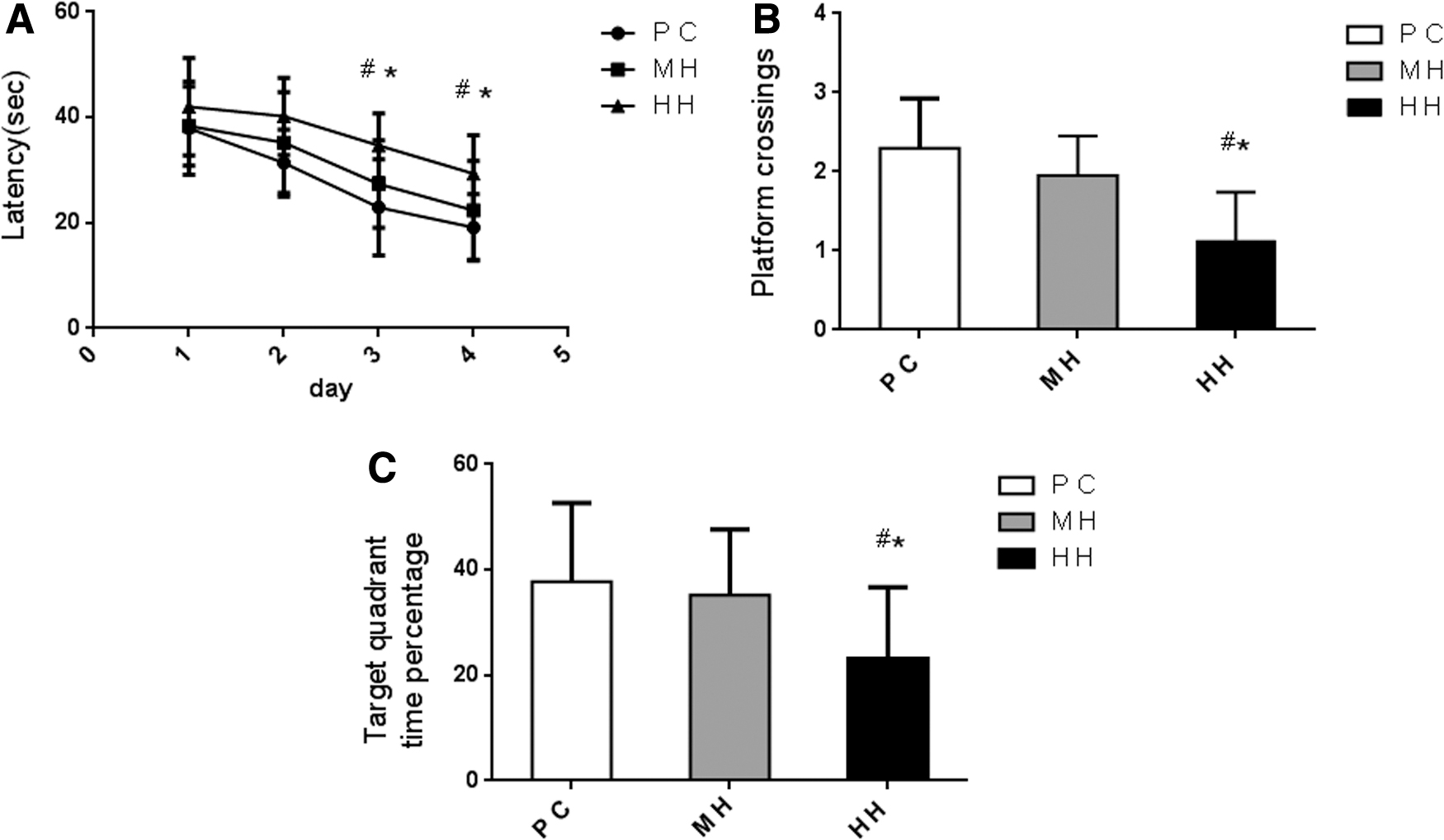
Evaluation of the learning and memory ability of rats in each group (n = 20/group) by the water maze test.
Morphological changes of brain tissue in chronic hypoxia rats
Nissl staining
Compared with the PC and MH groups, neurons in the HC and frontal cortex (FC) of the HH group were damaged, exhibiting a disordered cell arrangement and reduced number of Nissl bodies, with a statistically significant difference (p < 0.01). The morphology of neurons in the MH group was normal, exhibiting an ordered arrangement and slightly reduced Nissl bodies. There was no significant difference between the MH group and the PC group (p > 0.05), as given in Figure 2.

The Nissl staining of the hippocampal and cortical neurons ( × 400) (n = 8/group). The arrangement of the hippocampal and cortical neurons in the HH group was disordered.
Tunnel staining
Compared with the PC group, the apoptotic cells in the HC and FC of the HH group were increased significantly, and the apoptosis percentage was increased significantly (p < 0.01). Compared with the PC group, the apoptotic cells in the HC of the MH group were increased significantly and the percentage of apoptosis was increased (p < 0.05); however, no significant difference was observed in the cortical apoptotic cells (p > 0.05). Compared with the MH group, the apoptotic cells in the HC and cortex of the HH group were increased significantly alongside the significantly increased percentage of apoptotic cells (p < 0.01) (Fig. 3).
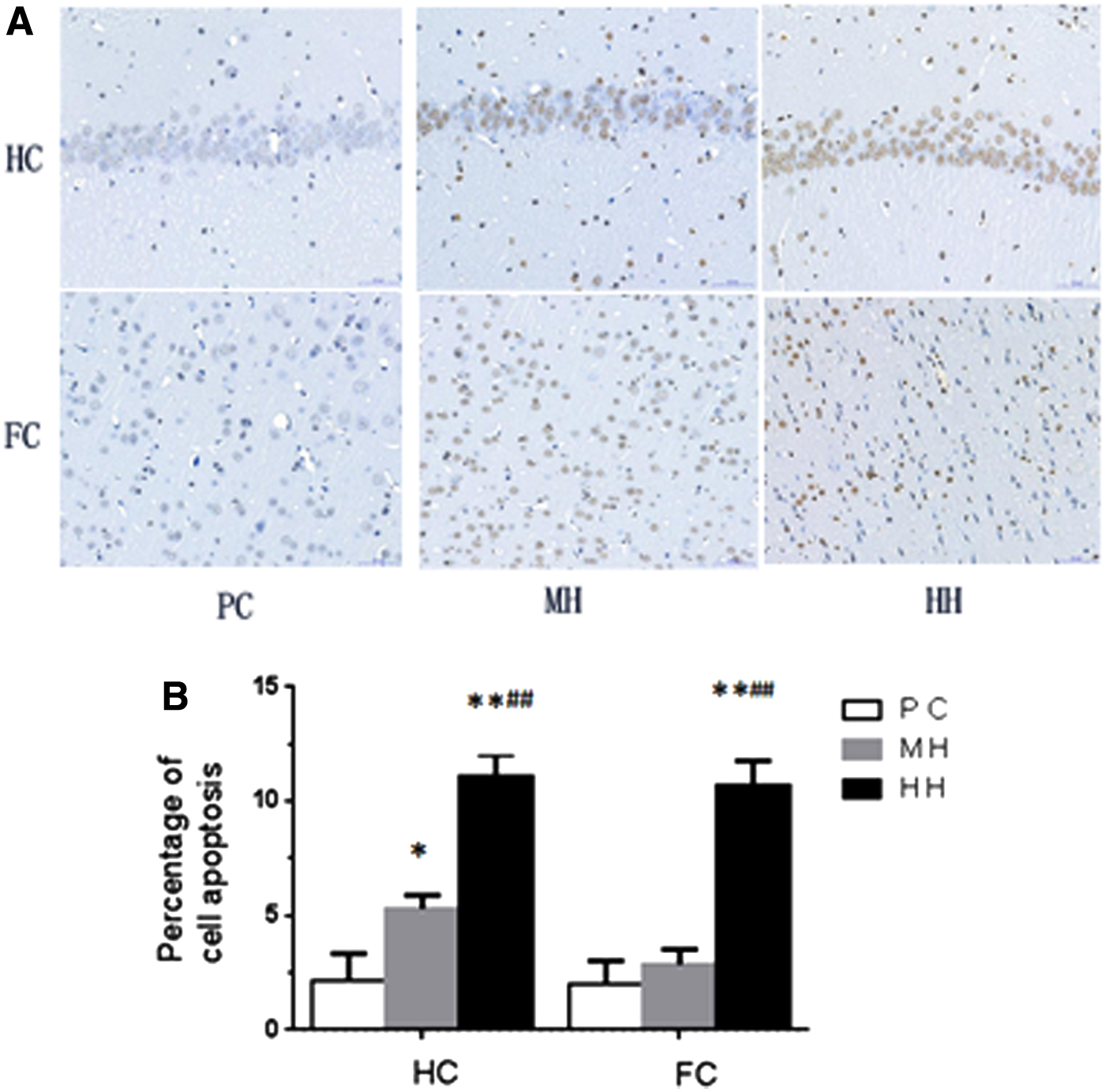
Electron microscopic observation
Compared with the PC group, the cells in the HH group were in an irregular shape, slightly agglutinated in chromatin, marginally arranged, and with their nucleolus visible. Swollen mitochondria, partial dissolved mitochondria cristae, slightly dilated rough endoplasmic reticulum, slightly swollen synaptic structure, and loose nerve myelin structure, were observed as well. In the MH group, unsmooth nuclear membrane, slightly swollen mitochondria, slightly dilated rough endoplasmic reticulum, slightly swollen synaptic structure, and slightly loosened myelin sheath were observed, as given in Figure 4.

Ultrastructural observation of hippocampal and FC in each group (n = 6/group). The cells in the HH group presented irregular shape, swollen mitochondria, dilated rough endoplasmic reticulum, and slightly swollen synaptic structure. In the MH group, the nuclear membrane was not smooth with slightly swollen mitochondria, slightly dilated rough endoplasmic reticulum, and slightly swollen synaptic structure being observed. In the PC group, mitochondria and rough endoplasmic reticulum displayed little change and the synaptic structure was normal.
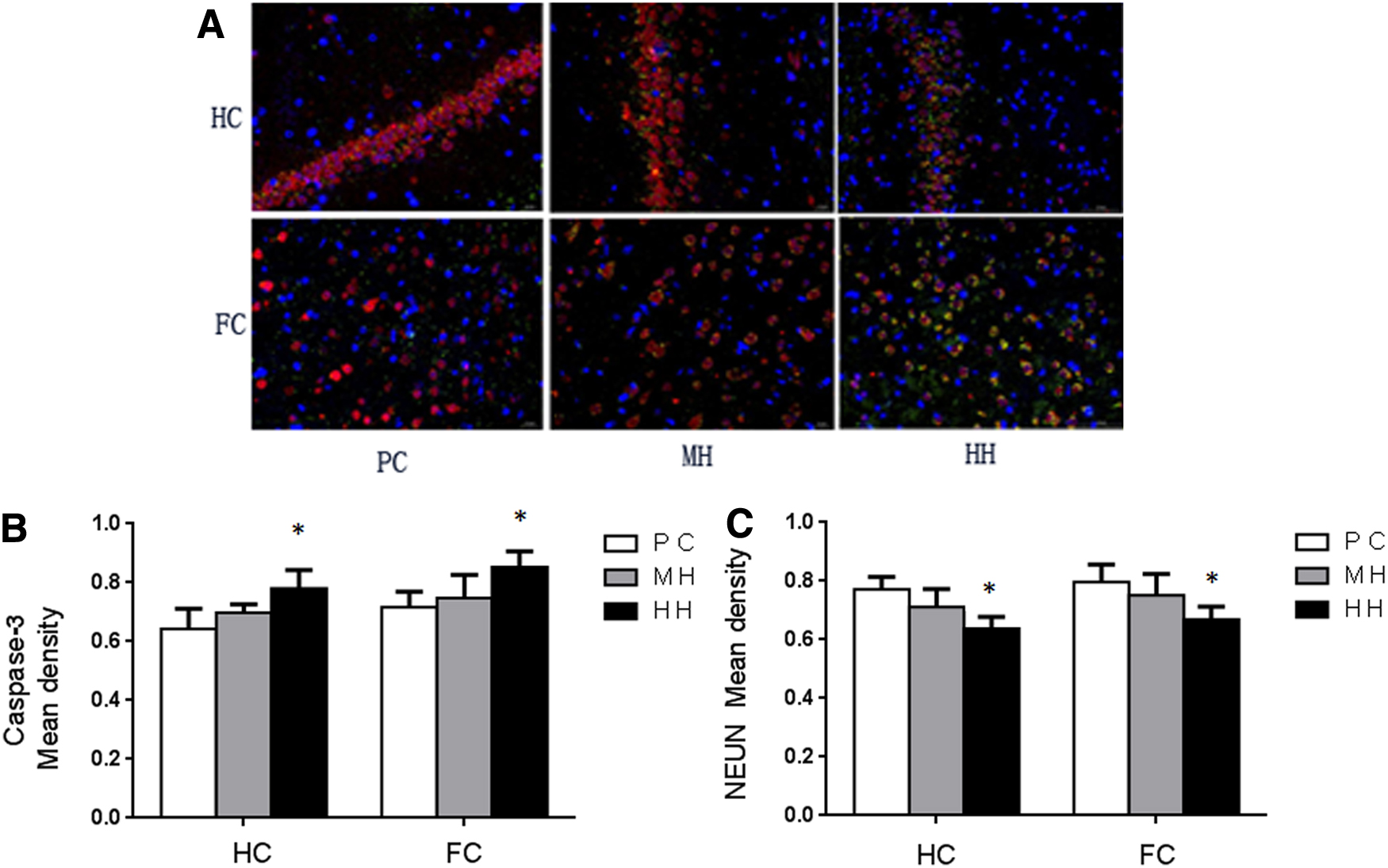
Immunofluorescence double staining of neuronal apoptosis-related proteins in chronic hypoxia rats (Fig. 5A)
Compared with the PC group, the content of Caspase-3 protein in the HC and FC of the HH group was increased significantly (p < 0.05). In the HH group, the content of NEUN protein in HC and cortex was significantly decreased (p < 0.05). There was no significant change in the content of Caspase-3 and NEUN protein in the HC and cortex tissues of the MH group (Fig. 5B, C).
Expression of apoptosis-related protein and glutamate receptor mRNA in brain tissue of chronic hypoxia rats
The expression of Caspase-3 gene in the HC and FC of the HH group was significantly higher than that of the MH group and PC group (p < 0.01); however, there was no significant difference between the MH group and PC group (p > 0.05, Fig. 6A). Compared with the PC and MH groups, the expression of the NR1 receptor gene in the HC was significantly increased in the HH group (p < 0.05). Compared with the PC group, the expression of the NR1 receptor in the HH and MH groups was increased significantly (p < 0.01; Fig. 6B). The expression of the GLU2 receptor gene in the HC of the HH and MH groups was significantly increased compared with the PC group (p < 0.05). The expression of the GLU2 receptor gene in the cortex of the HH and MH groups was significantly increased compared with the PC group (p < 0.01; Fig. 6C).
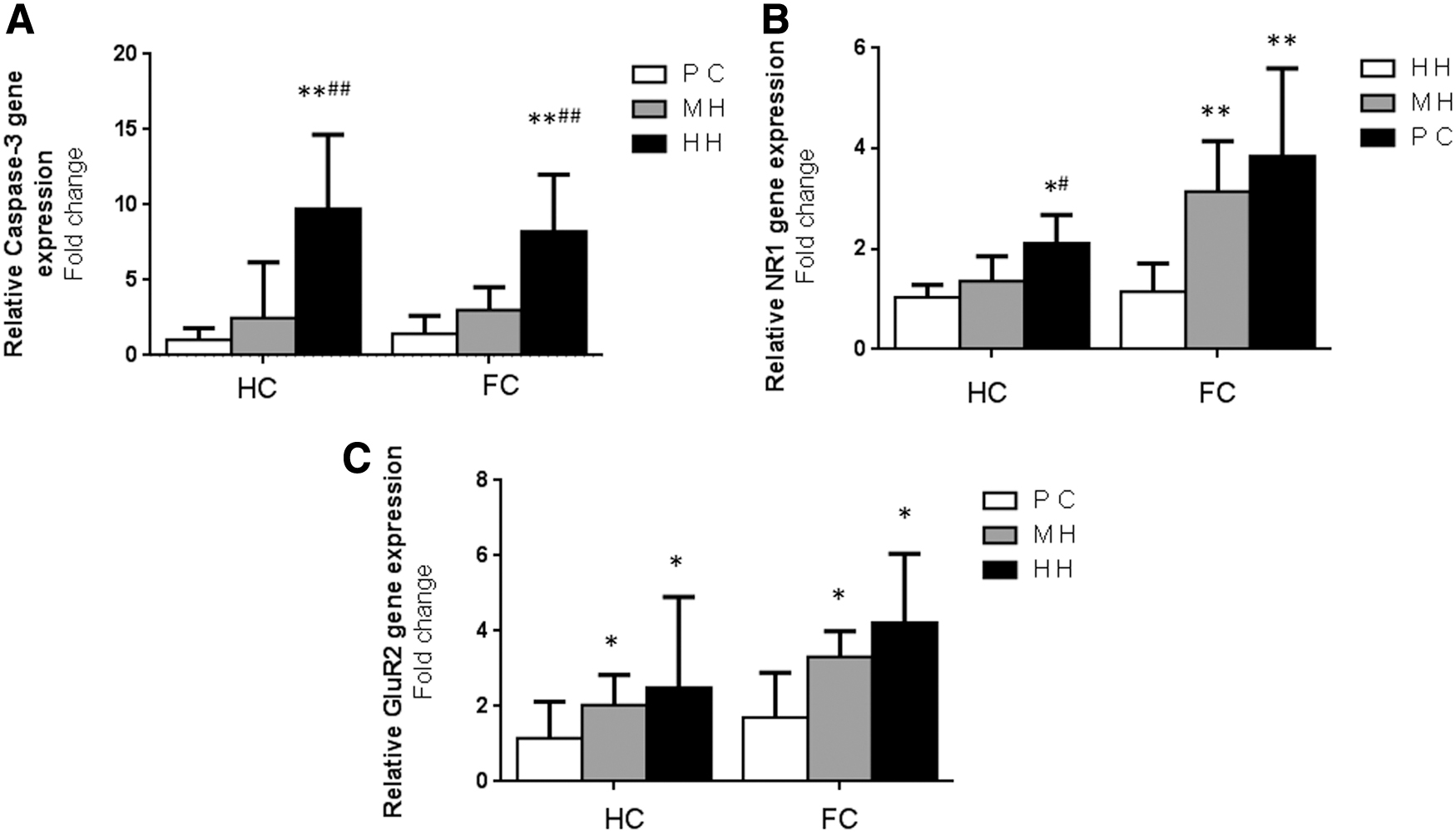
RT-PCR of glutamate receptor and Caspase-3 (n = 6/group).
Expression of apoptosis-related protein and glutamate receptor protein in brain tissue of chronic hypoxia rats
The expression of the cleaved Caspase-3 protein (17 kDa) in the HC and cortex of the HH group was significantly higher than that in the MH and PC groups (p < 0.01) (Fig. 7A). There was no significant difference between the MH group and PC group (p > 0.05, Fig. 7B). Compared with the PC and MH groups, the expression of the NR1 receptor protein in the HH group was increased significantly (p < 0.05). No obvious difference was observed between the MH group and PC group (p > 0.05). Compared with the PC group and MH group, the expression of the NR1 receptor protein of cortex in the HH group was increased significantly (p < 0.01), However, no significant difference was observed in the NR1 receptor expression between the MH group and PC group (Fig. 7C). Compared with the PC and MH groups, the GluR2 receptor protein expression in the HC and cortex of the HH group was significantly increased (p < 0.05), and there was no significant difference between the MH group and PC group (Fig. 7D).
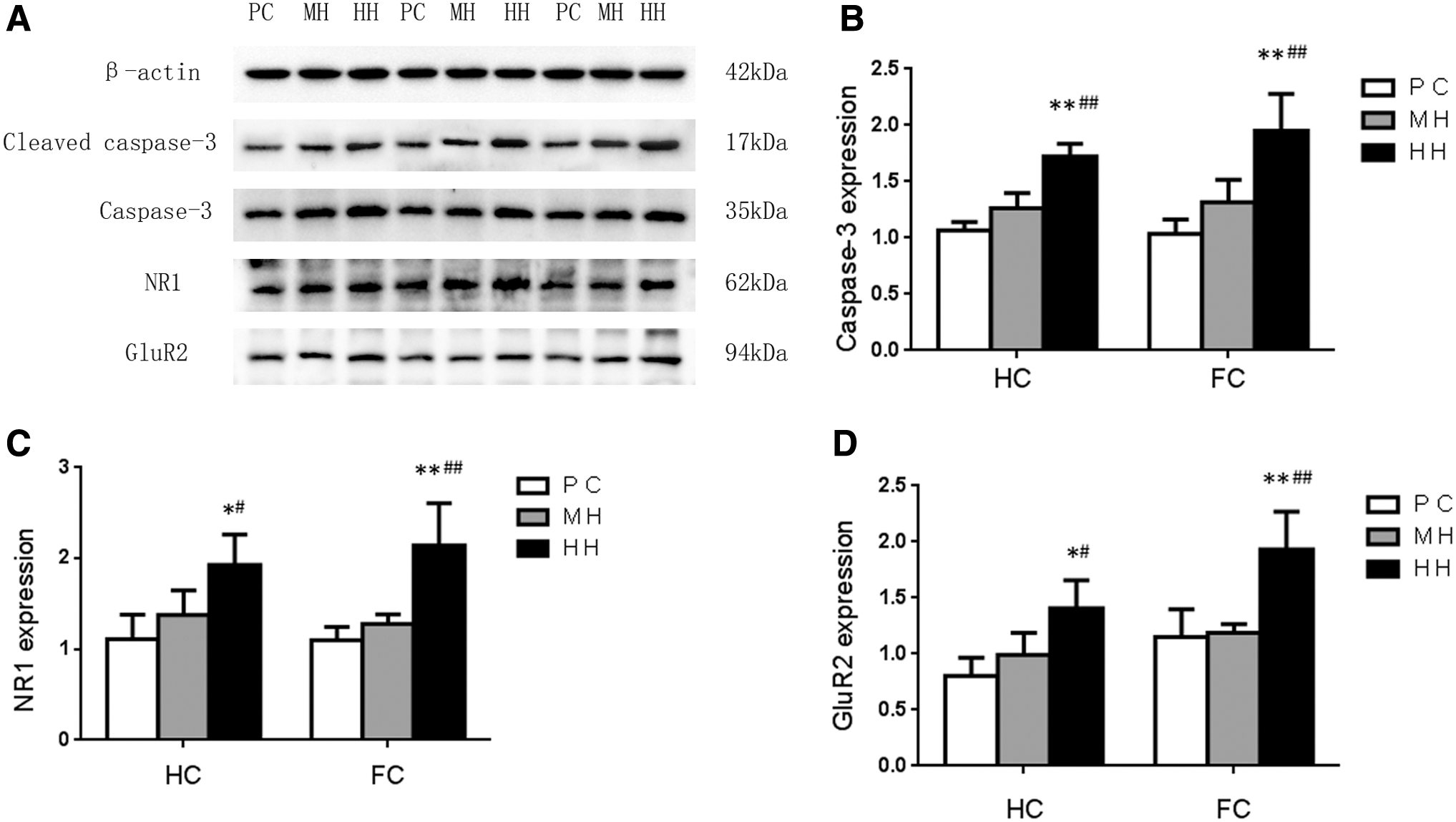
Biochemical detection of excitatory toxicity in brain tissue of rats with chronic hypoxia
Compared with the PC group, the activity of GR in the HC and FC of the HH group was significantly decreased (p < 0.01). The activity of GR in the HC and cortex of the HH group was decreased compared with the MH group; however, the difference was not statistically significant (p > 0.05). Compared with the PC group, the activity of GR in the HC and cortex in the MH group was decreased; however, the difference was not statistically significant (p > 0.05, Fig. 8A). Compared with the PC and MH groups, the activity of glutathione peroxidase in the HC and cortex of the HH group was significantly increased (p < 0.05). However, there was no significant difference in the glutathione peroxidase activity in the HC and cortex between the MH group and PC group (Fig. 8B). Compared with the PC and MH groups, the inhibitory ability against the free radicals of the HC and cortex in the HH group was significantly decreased (p < 0.05). However, there was no significant difference between the MH group and PC group (p > 0.05, Fig. 8C). Compared with the PC and MH groups, the lipid peroxide content in the HC and cortex of HH group was increased significantly (p < 0.05). However, there was no significant difference between the MH group and PC group (p > 0.05, Fig. 8D).

Biochemical detection of excitotoxicity (n = 6/group).
Discussion
Studies on the cognitive impairment in animals exposed to chronic hypobaric hypoxia at different altitudes with an exposure time of more than 4 weeks have rarely been reported. Most of the relevant studies have been performed in a simulated low-pressure and low-oxygen cabin, with an intermittent hypoxia situation instead of sustained hypoxia. With the increase of altitude in the plateau environment, the content of oxygen in the unit volume of air gradually decreases, and the atmospheric pressure drops, leading to more damage to the brain function. To investigate the effects of chronic hypobaric hypoxia on the cognitive function and possible glutamate receptor-mediated excitotoxicity in rats at different altitudes, this study first conducted the chronic hypobaric hypoxia experiment in natural environments at three different altitudes. Rats exposed to chronic hypobaric hypoxia conditions were examined before the experiment and had normal limb movements. According to the water maze experiment, the average escape latency, the number of crossing platforms, and time of crossing the target quadrant of rats in the HH group were significantly different from that of the MH and PC groups, indicating that the chronic hypobaric hypoxia environment at a high altitude significantly impaired the learning and memory ability of the rats. A study on the cognitive test of adults and children exposed to high altitude for longer periods found that the cognitive function was decreased significantly at high altitude and in a hypobaric hypoxic environment (Rimoldi et al., 2016), which is consistent with the current results. In addition, the results also showed that there was no statically significant difference between the MH group and PC group, indicating that the chronic hypoxia at moderate altitude had no obvious effect on the cognitive function in rats, consistent with the results of a clinical study (Roach et al., 2014). Another study also demonstrated that moderate altitude and hypoxic environment presented no significant effect on the human cognitive function (Pilmanis et al., 2016), which was consistent with the results of this study.
Hippocampal and FC neurons form the most important structural basis for learning and memory. After the damage occurs in the HC, the cognitive function is affected. Because the HC and cortex are one of the most sensitive brain tissues to hypoxia (Kim et al., 2013), it can cause irreversible neuronal damages such as neuronal apoptosis, degeneration, and necrosis (Miles and Kernie, 2008). According to the results of Nissl staining and electron microscopy observation, significant changes in the morphology of the HC and cortical brain were found in the HH group. The number of neurons decreased, exhibiting a disordered arrangement, partially denatured and necrotic neurons, reduced nucleus volume, chromatin agglutination, local inflammatory cells infiltration, swollen mitochondria, synapses, and other structures. In the MH group, degeneration was observed in a small number of neurons in the rats; however, there was no significant difference compared with the PC group. Studies explored the effects of acute hypobaric hypoxic environmental exposure on neurons reported a reduction in the cell number, disordered arrangement, and necrosis of a small number of neurons (Brochu et al., 2011). This was consistent with the research in chronic hypobaric hypoxia environment. However, some studies suggested that after exposure to hypobaric hypoxia for 3 days, the morphological changes of the rat brain tissue were the most obvious (Nair et al., 2011). However, with the prolongation of exposure time, the damage in the HC might be gradually alleviated. In this study, chronic persistent hypoxia was still harmful to the hippocampal and cortical neurons. Therefore, it is considered that chronic persistent hypoxia is a form of a cumulative and irreversible damage process. Once the neuronal damage occurs, it is irreversible. Hypoxic acclimatization may improve the damage of acute hypoxia brain tissue to some extent; however, its effect on chronic hypobaric hypoxia damage is not clear and requires further investigation.
The direct cause of cognitive impairment induced by the chronic hypobaric hypoxic environment may be related to the increase of neuronal necrosis and apoptosis. One study found that liraglutide promotes cognitive impairment through inhibition of hippocampal neuronal apoptosis (Yan et al., 2019). Another study on chronic intermittent hypoxia showed that H2 can alleviate neurocognitive impairment induced by intermittent hypoxia through antioxidant and antiapoptotic effects (Li et al., 2018), indirectly proving that the cognitive impairment is associated with the apoptosis of hippocampal neurons. The results of the TUNNEL in this study showed that the number of apoptotic cells and the percentage of apoptosis were increased significantly in the HC and cortex of rats in the HH group. Immunofluorescence results showed that the Caspase-3 protein levels in the HC and cortex were increased in the HH group. The results of real-time fluorescence quantitative PCR and western blot were also showed a significant enhancement of the gene expression and protein level for Caspase-3 in the HC and cortex of the HH group, indicating that the exposure to high-altitude chronic hypobaric hypoxia environment could aggravate neuronal apoptosis, which may be the main cause for cognitive impairment. This result is consistent with the findings of a study on cognitive impairment induced by extracorporeal circulation (Zheng et al., 2019). Brain cognitive impairment was correlated with apoptosis, oxidative stress, and so on (Shwe et al., 2018). It is believed that prolonged exposure to high-altitude chronic hypobaric hypoxic environment can aggravate neuronal apoptosis, which may be an important cause for cognitive impairment.
The exact mechanism of how chronic hypobaric hypoxia leads to increased neuronal apoptosis and induces cognitive impairment is still unclear. In recent years, the role of excitotoxicity mediated by glutamate and its receptors in the cognitive impairment induced by hypoxia has gained notoriety among researchers. It is generally acknowledged that the NMDA receptor is a hetero-tetramer composed of two NR1 subunits and two NR2 subunits (Paoletti, 2011). The NR1 subunit is composed of 938 amino acids and the two NR1 subunits constitute the main structure of the ion channel, which determines the main properties of the NMDA receptor (Pagadala et al., 2013). The AMPA receptor is composed of four subunits including the GluR1, GluR2, GluR3, and GluR4. Wherein GluR1, GluR3, and GluR4 could increase the permeability of AMPA receptor to Ca2+, the GluR2 subunit mainly reduces the permeability of Ca2+. Some researchers believe that apoptosis and cell necrosis mainly depend on the severity of the stimulation to the NMDA receptor. Overexpression of the NMDA receptors was discovered to be an important mechanism for cognitive impairment in chronic intermittent hypoxia (Jagadapillai et al., 2014; Sasaki et al., 2015). At present, there have been few studies on the animals exposed to chronic hypoxia at different altitudes. The results of this study demonstrated that the expression of subunit NR1 receptor of glutamate NMDA receptor at the gene and protein level was significantly increased in the HH group, and the expression of NR1 receptor in the cerebral cortex of the MH group was significantly higher than that in the PC group, indicating that the enhancement degrees of NMDA receptor expression was different in the hypoxic environment at different altitudes. However, it was also found that the expression of the subunit GluR2 receptor of the AMPA receptor in the HC and FC of the HH and MH groups was higher than that of the PC group, which is inconsistent with some reports (Beppu et al., 2013). Some researchers hypothesize that GluR2 could alleviate Ca2+ overloading (Chen et al., 2012). The inadequacy of this study lies in that it fails to address the effect of GluR2 on Ca2+ permeability under hypobaric hypoxia environment and it should be investigated in future studies.
Under chronic hypobaric hypoxia condition, whether the enhanced expression of glutamate receptor NR1 leads to excitotoxicity and induces cognitive impairment has been the critical problem addressed in this study. The excitotoxicity of the NMDA receptor is mainly determined by the Ca2+ influx (Afanador et al., 2013). Meanwhile, the accumulation of Ca2+ in mitochondria can lead to metabolic acidosis and free radical production (Lau et al., 2009). However, free radicals play an important role in glutamate excitotoxicity (He et al., 2011; Astori and Lüthi, 2013). Many studies have found that antioxidants play a neuroprotective role in the glutamate-induced excitotoxicity (Gonzalez-Zulueta et al., 1998; Muscoli et al., 2004; Peluffo et al., 2006; Yoon et al., 2013), indirectly proving the crucial role of free radicals in excitotoxicity. This study found that the activity of GR in the HH group was significantly reduced, whereas the glutathione peroxidase activity was significantly enhanced, indicating that the ability of glutathione as an oxidant to scavenge free radicals was decreased significantly. Further investigation on the ability to inhibit free radicals found that the ability to inhibit free radicals in the HH group was significantly lower than that in the PC and MH groups, indirectly proving that in the chronic hypobaric hypoxic environment at high altitude, the antioxidant capacity was decreased and that the free radicals were excessively generated. This could at least partly explain the existence of glutamate-mediated excitotoxicity in a chronic hypoxic environment. A study on chronic intermittent hypoxia found that the excitotoxicity mediated by glutamate could be observed after 7 days of intermittent hypoxia (Huo et al., 2014). In another study, rats exposed to simulated hypobaric oxygen chamber for 5 days at 6,000 m altitude, exhibited spatial memory impairment. At the same time, the activity of lactate dehydrogenase was enhanced, whereas the activity of glutathione and superoxide dismutase was decreased, suggesting the existence of oxidative stress, which was consistent with this study on chronic hypoxia (Shi et al., 2012).
The study also found no significant difference in the morphology of hippocampal and FC damaged by the chronic hypobaric hypoxic environment. The expression and oxidative stress of Caspase-3 and glutamate receptors in the hippocampal and cortical tissues were consistent, indicating that there was no regional difference in the damage of chronic hypobaric hypoxia to the brain tissue. This is consistent with other reports (Franco-Enzástiga et al., 2017; Thomas et al., 2017; Ko et al., 2019).
Conclusions
This study investigated the behavior, morphology, and molecular biological aspects of glutamate excitotoxicity in rats exposed to a chronic hypobaric hypoxic environment. We found that exposure to high-altitude chronic hypobaric hypoxia can produce glutamate excitotoxicity and eventually induce cognitive impairment. However, glutamate excitotoxicity and cognitive impairment were not observed in the hypobaric hypoxic environment at middle altitude. Similarly, there was no regional difference in brain damage induced by chronic hypobaric hypoxia exposure.
Footnotes
Author Contributions Statement
G.-R.l. designed the experiment, W.Y. and L.J. participated in the experiment operation, and Z.Y. participated in the statistical analysis.
Acknowledgments
The authors thank the Basic Research Program of Qinghai Provincial Science and Technology Department for funding of this project.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Basic Research Program of Qinghai Provincial Science and Technology Department (2020-ZJ-757).