
Abstract
Takezawa, Toshihiro, Shohei Dobashi, and Katsuhiro Koyama. Cardiorespiratory response and power output during submaximal exercise in normobaric versus hypobaric hypoxia: a pilot study using a specific chamber that controls environmental factors. High Alt Med Biol. 22: 201–208, 2021.
Background:
Many previous studies have examined hypoxia-induced physiological responses using various conditions, e.g., artificially reduced atmospheric oxygen concentration [normobaric hypoxia (NH) condition] or low barometric pressure at a mountain [hypobaric hypoxia (HH) condition]. However, when comparing the results from these previous studies conducted in artificial NH and HH including real high altitude, we must consider the possibility that environmental factors, such as temperature, humidity, and fraction of inspired carbon dioxide, might affect the physiological responses. Therefore, we examined cardiorespiratory responses and exercise performances during low- to high-intensity exercise at a fixed heart rate (HR) in both NH and HH using a specific chamber where atmospheric oxygen concentration and barometric pressure as well as the abovementioned environmental factors were precisely controlled.
Methods:
Ten well-trained university students (eight males and two females) performed the exercise test consisting of two 20-minute submaximal pedaling at the intensity corresponding to 50% (low) and 70% (high) of their HR reserve, under three conditions [NH (fraction of inspired oxygen, 0.135; barometric pressure, 754 mmHg), HH (fraction of inspired oxygen, 0.209; barometric pressure, 504 mmHg), and normobaric normoxia (NN; fraction of inspired oxygen, 0.209; barometric pressure, 754 mmHg)]. Peripheral oxygen saturation (SpO2) to estimate arterial oxygen saturation and partial pressure of end-tidal carbon dioxide (PETCO2) were monitored throughout the experiment.
Results:
SpO2, PETCO2, and power output at fixed HRs (i.e., pedaling efficiency) in NH and HH were all significantly lower than those in NN. Moreover, high-intensity exercise in HH induced greater decreases in SpO2 and power output than did high-intensity exercise in NH (NH vs. HH; SpO2, 78.2% ± 5.0% vs. 75.1% ± 7.1%; power output, 120.7 ± 24.9 W vs. 112.4 ± 23.2 W, both p < 0.05). However, high-intensity exercise in HH induced greater increases in PETCO2 than did high-intensity exercise in NH (NH vs. HH; 54.2 ± 5.9 mmHg vs. 57.2 ± 3.4 mmHg, p < 0.01).
Conclusions:
These results suggest that physiological responses and power output at a fixed HR during hypoxic exposure might depend on the method used to generate the hypoxic condition.
Introduction
Exercise trainings in real high altitudes or in artificial simulated hypoxic chambers (hypoxic training) are well known as one of the special training strategies for improving various exercise performances, including endurance and repeated exercise performances (Bonetti and Hopkins, 2009; Millet and Brocherie, 2020). These underlying mechanisms remain unknown, the partial oxygen pressure (PO2) is considered to be one of candidates to induce various physiological adaptations by high-altitude and hypoxic exercise training.
PO2 in inspired gas (PIO2), which reflects oxygen availability in the body, is calculated as the atmospheric oxygen (O2) concentration multiplied by the barometric pressure. At a real high altitude, it decreases owing to reduction in the barometric pressure without changes in the atmospheric O2 concentration; this environment is generally called hypobaric hypoxia (HH). In contrast, most of artificial hypoxic conditions are called normobaric hypoxia (NH), wherein PO2 is decreased owing to the reduction in the O2 concentration in the atmosphere without alterations in the barometric pressure.
Although the classical hypothesis of Bert (1943) demonstrated that physiological responses and their associated adaptations might be similar between NH and HH conditions with the same PIO2, the recent growing evidence suggests that physiological responses during exercise differ between HH and NH (Viscor et al., 2018). Indeed, Faiss et al. (2013) reported that the exercise-induced increase in ventilation throughout 24 hours of hypoxic exposure was lesser in HH than in NH. Moreover, pedaling exercise performances under both NH and HH were lower than that under normobaric normoxia (NN), whereas the magnitude of performance decrements was greater in HH than in NH (Beidleman et al., 2014).
However, the previous studies investigating the differences between NH and HH did not consider the possibility that the potential variables, such as temperature, humidity, airflow, solar radiation, and landscape in high-altitude conditions far differ from those under sea-level NH conditions (Coppel et al., 2015), might have a significant impact on physiological responses between NH and HH. Moreover, exercise in a closed chamber might increase the partial pressure of carbon dioxide (PCO2) (Bowtell et al., 2014), leading to increased fraction of inspired carbon dioxide (FICO2), which consequently affects physiological responses and exercise performances (Fan and Kayser, 2013).
Thus, the purpose of this study was to investigate whether cardiorespiratory responses and exercise performances differ between NH and HH wherein environmental factors (temperature, humidity, airflow, solar radiation, landscape, and FICO2) are strictly controlled. To test this, we assessed physiological responses and exercise performances [i.e., power output at a fixed heart rate (HR)] during acute low- and high-intensity pedaling exercises using a specific chamber that can generate both NH and HH and control the abovementioned environmental factors.
Materials and Methods
Participants
Ten well-trained university students (age: 20.0 ± 0.7 years, height: 166.2 ± 7.1 cm, body weight: 73.6 ± 18.7 kg, eight males and two females) who were able to perform high-intensity pedaling exercises were recruited in this study. None of the participants had any history of cardiovascular, cerebrovascular, or respiratory diseases. They had not smoked, taken medications, and been to altitudes of more than 2,000 m for 3 months before the study. Each participant was informed of the purpose, experimental procedures, and possible risks involved in this study. Written informed consent was obtained before the start of the experiments. All experimental procedures of this study were approved by the Human Research Ethics Committee of the University of Yamanashi in Japan (approval number, H30-001) and performed in accordance with the guidelines of the Declaration of Helsinki.
Experimental environment
The participants were required to complete three exercise tests under NH, HH, and NN. All exercise tests were performed in the same custom-made specific chamber (width: 1,500 mm × length: 3,000 mm × height: 1,900 mm) (Fig. 1A). This chamber had a durability against decompression until 504 mmHg. Inhaling gas from a preset cylinder was supplied to the participants through a scuba-diving mask (GULL MANTIS FULLFACE GM-1582, Tokyo, Japan) with a regulator (Cayman1180400259; Johnson Outdoor, Inc., Racine, WI); conversely, exhaling gas was discharged inside the chamber through the mask attached to a low-resistance one-way valve with a flow meter. Although the PCO2 in the chamber increased over time in this experimental system, the PICO2 did not increase, as the inspired gas was supplied from only the cylinder filled with the experimental gas.
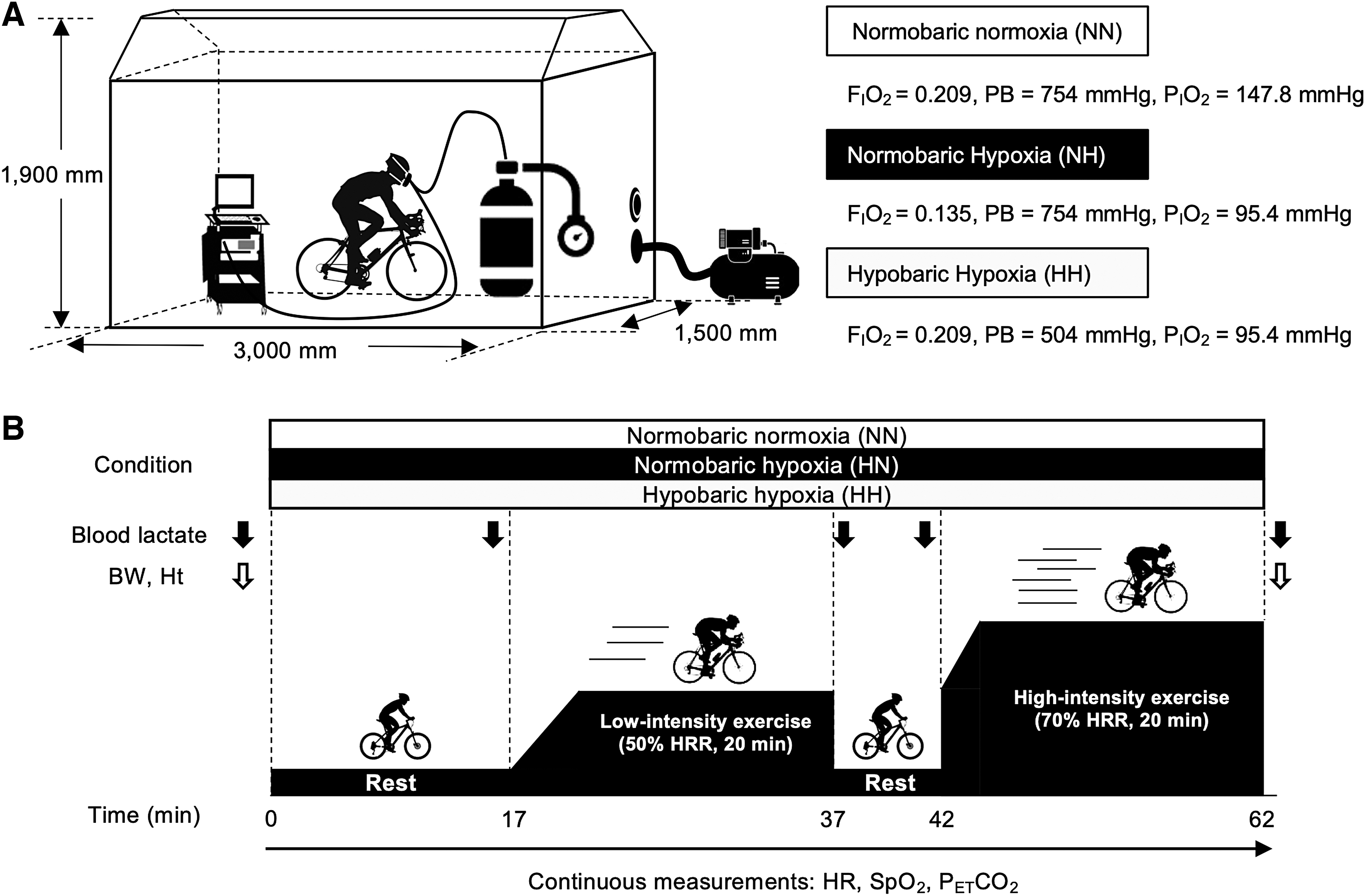
Overview of the experiment.
The composition of the experimental gas was 20.9% O2, 0.06% carbon dioxide (CO2), and 79.04% nitrogen (N2) in NN and HH and 13.5% O2, 0.06% CO2, and 86.44% N2 in NH. The barometric pressure in NN and NH was set at 754 mmHg (corresponding to ∼65 m above sea level, the height of laboratory), whereas that in HH was set at 504 mmHg (corresponding to ∼3,500 m above sea level). To generate HH, we conducted decompression by extracting the air inside the chamber using an air compressor (Rocking Piston Type Dry Pumps DOP-80SP; ULVAC Co. Ltd., Kanagawa, Japan).
Because of a methodological limitation in decompression from the chamber, decompression had to start 2.5 minutes after the participants entered the chamber. Our preliminary test revealed that a 14.5-minute decompression was needed to reach the target barometric pressure, that is, 504 mmHg; in other words, HH was completely prepared 17 minutes after the participants entered the chamber. Through these operations, we set the PIO2 in NN at 148 mmHg and that in the two hypoxic conditions (NH and HH) at 95.4 mmHg in the different generation methods. Moreover, the temperature and relative humidity in the chamber were controlled at 21°C and 55%, respectively, using a room air conditioner (CU-225CF; Panasonic Co. Ltd., Osaka, Japan) throughout the experiments.
Experimental procedure
Figure 1B shows an overview of the experimental procedure. On the experimental days, after a 5-minute warm-up (dynamic and static stretches) outside the chamber, the participants wore a respiratory mask and rested for 17 minutes on a cycle ergometer (AEROBIKE 75XLII; CONAMI Sports Life Co. Ltd., Tokyo, Japan) in the chamber. Subsequently, they started to pedal on the ergometer, to which ear sensors were connected to continuously measure their HR during the exercise.
For the first 5 minutes, the exercise intensity was gradually increased by 30 W/min and then further increased by 20 W/min in a step-like manner until the participants' 50% of heart rate reserve (HRR) was predetermined under NN by Karvonen formula (Robergs and Landwehr 2002; She et al., 2015). The average time to achieve 50% of HRR was ∼5 minutes. Once the HR reached 50% of HRR, the exercise intensity (i.e., load of pedaling) was automatically adjusted to maintain the target HR. Hence, this cycling ergometer could match the individual relative exercise intensity among the three different conditions based on the changes in the HR during pedaling. This method was already applied in a previous study that examined physiological responses during exercise under normoxic and hypoxic conditions (Komiyama et al., 2015). During the low-intensity exercise at 50% of HRR, the pedal cadence was kept at 70 rpm.
After a 5-minute rest interval, the participants restarted the high-intensity pedaling exercise at 70% of HRR (based on the HRR in NN). For the first 5 minutes, they were instructed to keep a pedal cadence of 95 rpm until their HR reached 70% of HRR and to decrease it gradually to 70 rpm thereafter. This manipulation enabled the participants to reach 70% of HRR within 5 minutes (the average time to achieve 70% of HRR was ∼2 minutes and 40 seconds) and to avoid unstable pedaling at 70 rpm during the subsequent constant exercise. Similar to that during the low-intensity exercise, the pedaling load during the high-intensity exercise was continuously modulated to maintain 70% of HRR. Each experiment was conducted on the same time of the day with an interval of more than 1 week between trials to eliminate the effects of diurnal variation and carry over. The order of the NN, NH, and HH trials was single blinded and counterbalanced across the participants.
Physiological measurements
Throughout the experiments, respiratory gas was continuously collected, and partial pressure of end-tidal carbon dioxide (PETCO2) was measured using an automatic gas analyzer (CO2SMO; NOVAMETRIX Co. Ltd., Wallingford, CT). Moreover, peripheral oxygen saturation (SpO2) to estimate arterial O2 saturation was also monitored, with the sensor of CO2SMO placed on the left index finger.
Capillary blood samples (0.5 μL) were obtained from the finger for measuring the blood lactate concentration and hematocrit level. The blood lactate concentration was measured using an automated lactate analyzer (Lactate Pro 2; Arkray, Tokyo, Japan) before entrance to the chamber (Pre) and immediately after the two exercises (low- and high-intensity exercises; Post Ex50%HRR and Post Ex70%HRR, respectively). In addition, we assessed the hematocrit level using a hematocrit reader (Kubota Corporation, Tokyo, Japan) as well as the body weight before and after the two exercises. Changes in the hematocrit level and body weight before and after the exercises were evaluated as an index of dehydration.
Power output
Exercise performance in our experiment was estimated by the power output during pedaling. As the cycle ergometer used in this study could memorize the participants' power outputs every minute during pedaling, we could evaluate the power output at the fixed HRs under each experimental condition as an index of exercise performance in this study.
Statistical analyses
For comparison of the cardiorespiratory variables (PETCO2 and SpO2) and power outputs during exercise, the averaged values during the second half (between 10 and 20 minutes) of the low- and high-intensity exercises were used because the physiological responses to the graded phase in relation to the exercise intensity were generally not stable. In terms of the body weight and hematocrit level, the relative changes from the values at the resting state before entrance to the chamber were assessed.
Data were presented as mean ± standard deviations. Statistical comparisons of the cardiorespiratory variables, power output, and index of dehydration were conducted using one-way analysis of variance (ANOVA) with Bonferroni post hoc analysis. Two-way repeated measures ANOVA was applied to assess the main effects and the interaction (trial × time) for comparison of the blood lactate concentrations. All statistical analyses were performed using JMP (ver.11.0; SAS Institute, Inc., Cary, NC). p-Values <0.05 were considered significant for all tests.
Results
Cardiorespiratory responses
Figure 2 shows the alterations in the SpO2 during the experiments. Although it took 17 minutes since the participants had entered the chamber to generate HH, the SpO2 immediately before the start of the exercise at 50% of HRR did not differ between NH and HH (Fig. 2A). The average SpO2 during the second half of the low-intensity exercise (50% of HRR) in NH and HH was significantly lower than that in NN (all p < 0.01; NH vs. NN, effect size: d = 3.9225, 1-β = 1.000; HH vs. NN, effect size: d = 3.0862, 1-β = 1.000; Fig. 2B). Moreover, the SpO2 in HH was significantly lower than that in NH (p < 0.01, effect size: d = 0.9573, 1-β = 0.9820, Fig. 2B). Similar with the SpO2 during the low-intensity exercise, the average SpO2 during the second half of the high-intensity exercise (70% of HRR) was also significantly lower in NH and HH than in NN (all p < 0.01; NH vs. NN, effect size: d = 3.4196, 1-β = 1.000; HH vs. NN, effect size: d = 4.1484, 1-β = 1.000; Fig. 2C). Additionally, the SpO2 during the high-intensity exercise in HH was significantly lower than that in NH (p < 0.01, effect size: d = 0.6698, 1-β = 0.8110, Fig. 2C).

Effect of NH and HH on the SpO2 during the low- and high-intensity exercises.
Figure 3 demonstrates the time course changes in the PETCO2 during the low- and high-intensity exercises in the three different conditions. The average PETCO2 during the second half of the low-intensity exercise in NH was significantly lower than that in NN (p < 0.01, effect size: d = 0.7462, 1-β = 0.9418, Fig. 3B), and that in HH was significantly higher than that in NH (p < 0.01, effect size: d = 1.3674, 1-β = 0.9999, Fig. 3B). Conversely, the average PETCO2 during the second half of the high-intensity exercise was significantly lower in NH and HH than that in NN (all p < 0.01; NH vs. NN, effect size: d = 0.8562, 1-β = 0.9495; HH vs. NN, effect size: d = 0.4486, 1-β = 0.6137; Fig. 3C). Furthermore, the PETCO2 in HH was significantly higher than that in NH (p < 0.01, effect size: d = 0.7170, 1-β = 0.9257, Fig. 3C).
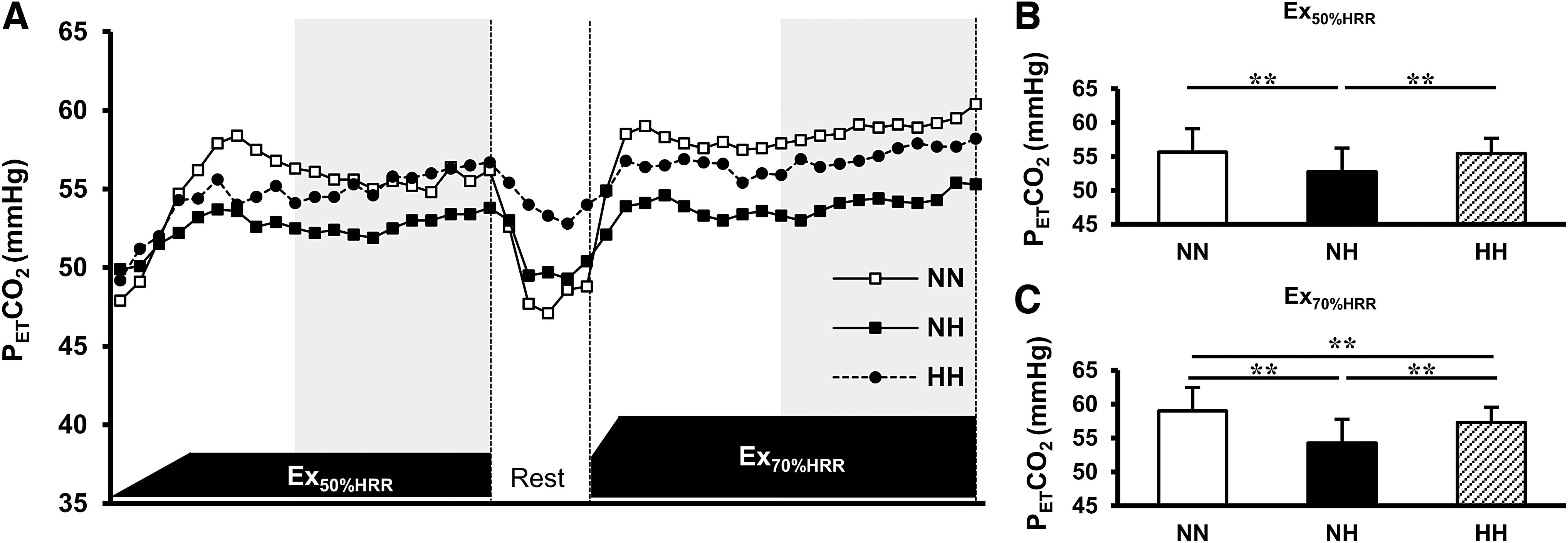
Effect of NH and HH on the PETCO2 during the low- and high-intensity exercises.
Power output
Figure 4 illustrates the power output during the low-intensity pedaling exercise in NN, NH, and HH. The average power outputs during the second half of the low-intensity exercise in NH and HH were significantly lower than that in NN (all p < 0.01; NH vs. NN, effect size: d = 1.9286, 1-β = 1.000; HH vs. NN, effect size: d = 1.9298, 1-β = 1.000). Moreover, the power output in HH was marginally lower than that in NH (p = 0.07, effect size: d = 0.6331, 1-β = 0.7761).
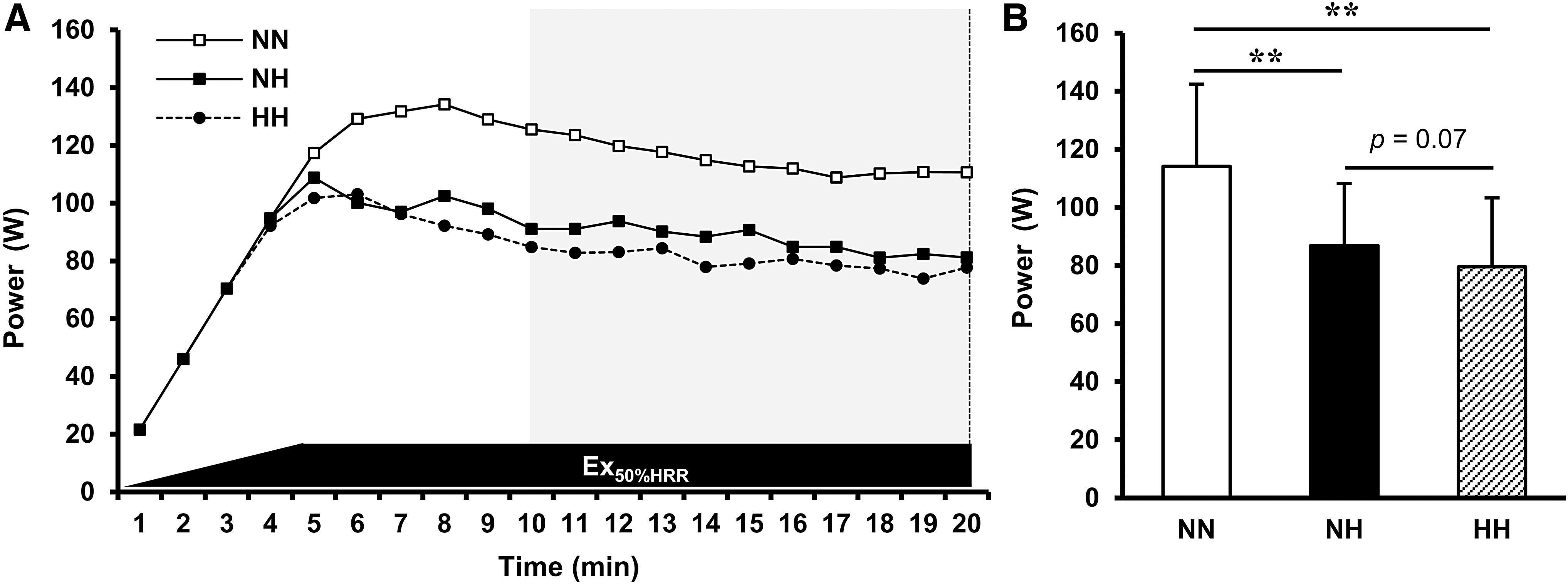
Effect of NH and HH on the power output during the low-intensity exercise.
The power outputs during the high-intensity pedaling exercise in NN, NH, and HH are described in Figure 5. The average power outputs during the second half of the high-intensity exercise in NH and HH were also significantly lower than that in NN (all p < 0.01; NH vs. NN, effect size: d = 2.8473, 1-β = 1.000; HH vs. NN, effect size: d = 3.1528, 1-β = 1.000). Additionally, the power output in HH was significantly lower than that in NH (p < 0.05, effect size: d = 1.0986, 1-β = 0.9965).
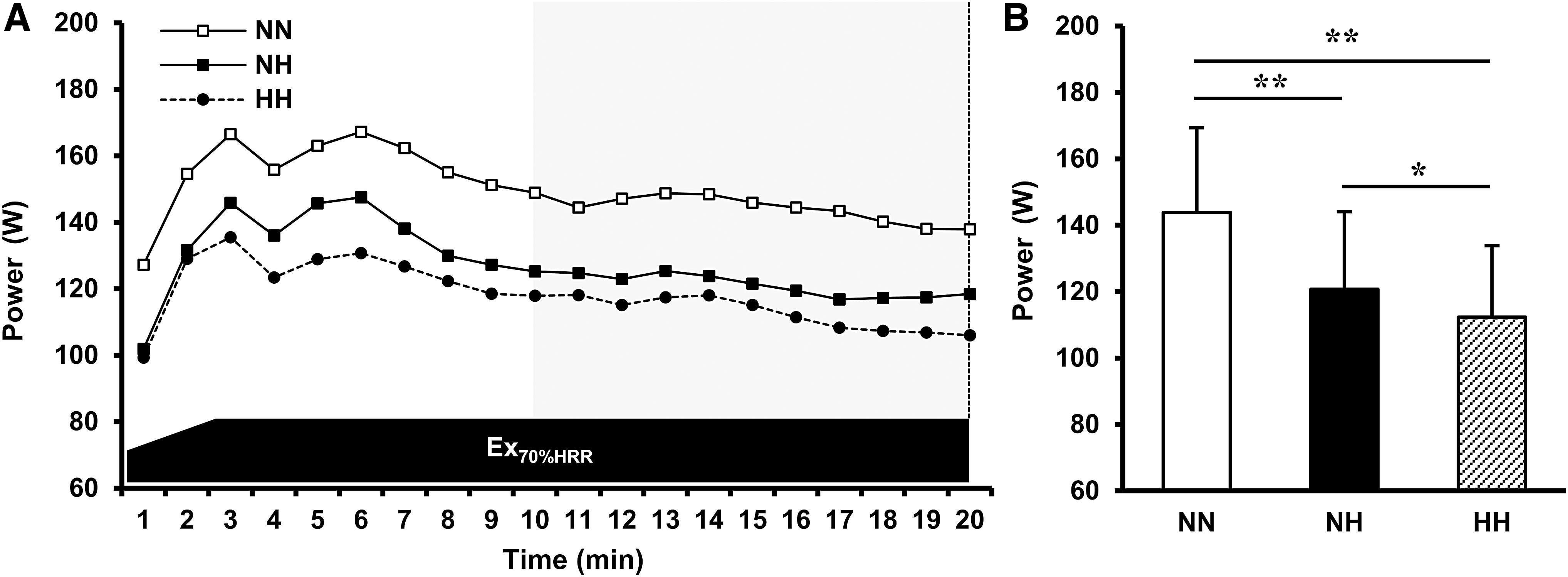
Effect of NH and HH on the power output during the high-intensity exercise.
Blood lactate concentration
Table 1 displays the changes in the blood lactate concentrations. Two-way ANOVA revealed a significant interaction between condition and time (p < 0.01, effect size: f = 0.4786, 1-β = 1.0000). The blood lactate concentrations increased as the exercise intensity also increased in all conditions, and those immediately after the high-intensity exercise in NH and HH were significantly higher than that in NN (all p < 0.05; NH vs. NN, effect size: d = 1.3688, 1-β = 0.9999; HH vs. NN, effect size: d = 1.2769, 1-β = 0.9997). However, no significant differences in the blood lactate concentrations were observed between NH and HH after the high-intensity exercise.
Blood Lactate Concentrations During the Low- (Ex50%HRR) and High-Intensity (Ex70%HRR) Exercises
Values are presented as mean ± standard deviations. Two-way ANOVA was performed.
p < 0.05 versus Pre, †p < 0.05 versus Post Ex50%HRR, §p < 0.05 versus NN at the same time period.
ANOVA, analysis of variance; HH, hypobaric hypoxia; HRR, heart rate reserve; NH, normobaric hypoxia; NN, normobaric normoxia.
Hydration status
Table 2 shows the changes in the body weight and hematocrit level after the exercises relative to those before the exercises. The relative changes in the body weight and hematocrit level were not significantly different among the three different conditions.
Changes in the Body Weight and Hematocrit Level After the Experiments Relative to the Baseline Values
Values are presented as mean ± standard deviations. One-way ANOVA was performed. No significant differences among the three experimental conditions were observed with regard to both changes in the body weight and hematocrit level.
Discussion
To the best of our knowledge, this is the first study to examine the physiological responses and power output at fixed submaximal HRs during low- and high-intensity exercises in NH and HH using a specific chamber wherein potential environmental factors (temperature, humidity, airflow, solar radiation, landscape, and FICO2) are strictly controlled. Our novel findings were that both the SpO2 and power output at fixed HRs during the high-intensity exercise in HH were significantly lower, and the PETCO2 was significantly higher than those in NH.
In this study, we simulated a hypoxic condition corresponding to an ∼3,500-m high altitude that athletes occasionally use for exercise training (Wilber, 2007). We found that the SpO2 throughout the exercise tests in NH and HH was significantly lower than that in NN (Fig. 2). Furthermore, the power outputs during the low- and high-intensity exercises in NH and HH also decreased in comparison with that in NN (Figs. 4 and 5). In contrast, the blood lactate concentrations immediately after the high-intensity exercise were significantly higher in NH and HH than in NN regardless of lower power outputs (Table 1). Katayama et al. (2010) reported that hypoxic exposure strongly shifted the dominant system for energy supply from oxidative to glycolytic during exercise and recovery in comparison with that during exercise at the same relative intensity under sea-level conditions. Based on the results from the previous study and this study, PIO2 reduction (NN, PIO2 = 147.8 mmHg; NH and HH, PIO2 = 95.4 mmHg) itself might be a powerful stimulus enough to impair O2 supply and exercise efficiency, irrespective of the different methods used to generate hypoxic conditions (NH and HH).
Interestingly, the decrease of the power output at a fixed HR during high-intensity exercise in HH was significantly greater, and marginally greater during low-intensity exercise, compared with NH (Figs. 4B and 5B). These findings are consistent with the results of previous studies that examined the differences in exercise performance under artificial NH and real high-altitude HH (Beidleman et al., 2014; Saugy et al., 2016). We speculate that the cardiorespiratory responses reflected by the alteration in the PETCO2 and SpO2 might, at least in part, explain the differences in the power output at a fixed HR between NH and HH. Generally, under the condition where the PIO2 is reduced, the respiratory rate increases to compensate for the decreased O2 supply to the peripheral tissues (Faiss et al., 2013). Along with this physiological reaction, blood CO2 is exhaled from the body through the lungs by increasing minute ventilation (VE); thereby, the decreased partial arterial pressure of CO2 might also reduce the PETCO2 (Townsend et al., 2016). However, the reduction in the PETCO2 in HH was significantly attenuated as compared with that in NH. Although the precise mechanism with respect to this phenomenon remains unknown, earlier studies have pointed out the effect of a low barometric pressure (Faiss et al., 2013; Coppel et al., 2015).
A previous systematic review has demonstrated the hypothesis that HH induces intravascular microbubble formation in the pulmonary circulation, increased alveolar dead space, altered fluid permeability, changes in chemosensitivity, and mismatch in ventilation and perfusion and might then lead to differences in cardiorespiratory responses compared with NH (Coppel et al., 2015). The reduction in VE owing to increases in the alveolar dead space might be associated with the increase in the PETCO2 and decrease in the SpO2 under HH when compared with those under NH (Faiss et al., 2013). From these noble previous findings, we speculate that the decrease in the SpO2 and attenuated decrease in the PETCO2 under HH might stem from the reduction in VE. Thus, the greater decrease in O2 supply to the active muscles in HH might induce further decrements in exercise efficiency compared with that in NH. However, we could not measure the respiratory rate, VE, and the alveolar dead space in this study. We must also acknowledge that acute hypobaria itself might decrease power output at a fixed HR accompanied with decreasing blood pressure. Therefore, further studies are required to confirm the hypothesis.
We also assessed the hydration status (i.e., changes in the body weight and hematocrit level) because this potential factor can affect exercise performances under hypoxic conditions (Castellani et al., 2010; Miyagawa et al., 2011). We found that this index did not differ among the three different conditions (Table 2), indicating that the hydration status might not have a significant influence on cardiorespiratory responses and exercise efficiency. However, a low barometric pressure could alter body fluid balance (e.g., increase in antidiuretic hormone and aldosterone levels) (Loeppky et al., 2005). Hence, the impact of body fluid balance on differences in exercise efficiency between NH and HH should be examined in more detail.
We employed unique technologies to control several potential environmental factors to examine the effect of the difference in the methods used for generating hypoxic conditions on physiological responses during exercise. However, some limitations still exist. First, the total exposure time for NH and HH was only 62 minutes in this study. A previous systematic review on physiological responses to NH and HH demonstrated that the SpO2 was altered between NH and HH during less than 60 minutes of hypoxic exposure and that differences in the SpO2 were damped with a much longer duration of hypoxic exposure (Coppel et al., 2015). That is, the physiological responses during hypoxic exposure might be dependent on the duration. Second, to generate a low O2 concentration, we set our experimental gas mixture in NH by increasing the density of N2. Although there are no reports concerning the effect of high concentrations of N2 on physiological responses, we should not ignore the possibility that they might affect cardiorespiratory responses and exercise efficiency. Third, all target HRs (50% and 70% of HRR) in this study were set on the basis of the values calculated from HRR under NN condition. However, previous reports suggested that hypoxia, irrespective of normobaria or hypobaria, decreases maximal HR compared with normoxia (Mourot 2018), and that resting HR in HH is significantly higher than that in NH (Savourey et al., 2003). Since Mourot and Millet (2018) also indicated that the maximal HR tended to differ between NH and HH in trained subjects, these potential confounders can raise the possibility that the relative intensities at fixed target HRs might not have been entirely equal between three different conditions. To examine whether substantial differences exist between HH and NH, we should also compare physiological responses and the muscular performance, along with measuring maximal HR under each hypoxic condition and then using these values to choose % HRR. Finally, as the sample size (n = 10) in this study was relatively small, future studies with larger populations are needed.
Conclusion
Despite the strict control for the environmental factors (temperature, humidity, airflow, solar radiation, landscape, and FICO2), we clearly observed reductions in the SpO2, PETCO2, and power output at a fixed HR in NH and HH. Moreover, the high-intensity exercise in HH induced greater decreases in the SpO2 and power output and an increase in the PETCO2 than did that in NH. Therefore, physiological responses during hypoxic exposure might depend not only on the PIO2 but also on the methodological difference in generating hypoxic conditions, that is, reduction in the atmospheric O2 concentration or barometric pressure.
Footnotes
Acknowledgments
Mr. Shiro Yokozawa and his employee in Cell Up Co. Ltd. (Gunma, Japan) designed and prepared the specific hypoxic chamber that we used in this study.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received to perform the experiment and publication for this study.