
Abstract
Bromer, Frederik Duch, Mikkel Bo Brent, Michael Pedersen, Jesper Skovhus Thomsen, Annemarie Brüel, and Casper Bindzus Foldager. The effect of normobaric intermittent hypoxia therapy on bone in normal and disuse osteopenic mice. High Alt Med Biol. 22: 225–234, 2021.
Background:
Systemic intermittent hypoxia therapy (IHT) has been shown to elicit beneficial effects on multiple physiological systems. However, only few studies have investigated the effect of long-term normobaric IHT on bone mass and mechanical and microstructural properties. The aim of the present study was to examine the effect of IHT on bone in both healthy and osteopenic mice.
Materials and Methods:
Thirty mice were stratified into four groups: Ctrl, Ctrl+IHT, Botox, and Botox+IHT. Osteopenia was induced by injecting Botox into the right hindlimb of the mice causing paralysis and disuse. IHT animals were placed in a normobaric hypoxia-chamber (10% oxygen) for 1 hour twice daily 5 days/week. Animals were sacrificed after 21 days, and DEXA, micro-computed tomography, and mechanical testing were performed on the femora.
Results:
As expected, Botox resulted in a significant reduction of bone mineral content (−23.4%), area bone mineral density (−19.1%), femoral neck strength (Fmax: −54.7%), bone volume fraction (bone volume/tissue volume: −41.8%), and trabecular thickness (−32.4%). IHT had no measurable effect on the bone properties in either healthy or osteopenic mice.
Conclusion:
The study confirmed that Botox led to loss of bone mass, deterioration of trabecular microstructure, and loss of bone strength. These changes were not influenced by IHT. Notably, IHT had no detrimental effect on bone in either healthy or osteopenic mice. This indicates that IHT of ailments outside of the skeletal system may be administered without causing harm to the bone.
Introduction
Systemic intermittent hypoxia therapy (IHT) is a potent stimulant of a multitude of different physiological systems. The reduction of available oxygen in the air provokes a wide range of systemic changes, from regulation of breathing frequency to cellular adaption by decreased energy consumption and adjustment of gene expression (Michiels, 2004). One possible pathway for the adaptive cellular response is hypoxia-inducible factor-1 (HIF-1) (Semenza, 2009). HIF-1 is upregulated when the cell senses low cellular oxygen levels (Wang et al., 1995; Semenza, 2007), which leads to increased transcription of a vast number of genes, including genes involved in erythropoiesis, angiogenesis, and autophagy (Wenger and Gassmann, 1997; Okuyama et al., 2006; Semenza, 2007; Kaelin and Ratcliffe, 2008). However, a large hypoxic stimulus is needed to induce HIF-1 pathways for therapeutic purposes. Another response to hypoxia is increased migration of stem cells from bone marrow to peripheral blood and altered mitochondrial regulation of enzymes related to metabolism (Hung et al., 2007; Gharib et al., 2010; Bergholt et al., 2019).
Studies investigating the effects of hypoxia on different physiological systems have described both beneficial and disadvantageous effects of IHT (Almendros et al., 2014; Camacho-Cardenosa et al., 2019). Therefore, when applying IHT by brief exposure to reduced oxygen levels alternating with normal oxygen levels, it is important that the short periods with reduced oxygen have an oxygen level low enough to cause advantageous cellular effects, but high enough to avoid unwanted effects (Navarrete-Opazo and Mitchell, 2014). Recent data seem to indicate that moderate hypoxia levels (9%–16% inspired O2) for short periods of time lead to the most beneficial outcome (Navarrete-Opazo and Mitchell, 2014). For the purpose of this study, IHT is defined as continuous periods of short-term exposure (minutes to hours) to air with an oxygen concentration lower than normal interspersed by longer periods of exposure to air with normal oxygen content. This is in contrast to chronic hypoxia, where the exposure to low oxygen concentrations is much longer (days to months). The intermittent hypoxia periods of altered oxygen concentrations in our IHT protocol were thus much longer than the short cycles of hypoxia seen in sleep apnea.
At present, only few studies have investigated the effect of IHT on bone tissue. One study showed that selective deletion of the von Hippel-Lindau gene, which normally downregulates expression of HIF-1, significantly increased trabecular bone volume fraction and bone vascularity in mice (Wang et al., 2007). Another study reported that exposing rats to a hypobaric IHT consisting of a barometric pressure of 430 mmHg (corresponding to an effective oxygen-level of 11.9%) for 5 hours/day, 5 days/week for 5 weeks increased area bone mineral density (aBMD) compared to control animals (Guner et al., 2013). In contrast, humans exposed to long-term hypobaric hypoxia by living at high altitude have been linked with osteopenia and osteoporosis (Tanaka et al., 1992; Basu et al., 2014). However, the underlying effects of IHT on bone mass, microstructure, and strength in animal models remain unknown.
To investigate the effects of IHT on bone in detail, it is important to consider not only healthy individuals but also individuals suffering from low bone mass. Osteoporosis is one of the most common bone diseases in humans (US Department of Health and Human Services, 2004). It is characterized by low aBMD and bone strength as well as deterioration of the trabecular microstructure, leading to increased risk of fractures (Sozen et al., 2017). Osteoporosis can be caused by aging, loss of oestrogen after the menopause, or by external factors such as disuse (Raisz, 2005; Atmaca et al., 2013; Lodberg et al., 2015). During disuse, the load applied to the skeleton is reduced, resulting in a rapid and substantial loss of bone mass (Krolner and Toft, 1983), eventually leading to osteopenia and osteoporosis (Minaire et al., 1974; Bloomfield, 2010). Multiple models of disuse osteopenia have been established in rodents (Komori, 2015; Lodberg et al., 2015). One method to induce disuse osteopenia is to inject botulinum toxin A (Botox) into the skeletal muscles of the hindlimb of the animal (Chappard et al., 2001; Vegger et al., 2014; Brent et al., 2021). This inhibits the release of acetylcholine from the neuromuscular junction leading to muscle paralysis and disuse (Rossetto et al., 2014). The result is a rapid loss of muscle mass of the affected hindlimb, followed by decreased bone mass and bone strength (Thomsen et al., 2012; Brent et al., 2018).
In recent years, the therapeutic potential of numerous different hypoxia protocols has been explored on various physiological systems. Currently, we investigate protocols applying normobaric oxygen levels of 10% for varying durations between 30 minutes and an hour, and initial results show some therapeutic effects. Due to the concerns of the association between chronic hypoxia and reduced aBMD, we wanted to investigate whether our IHT had any negative effects on bone parameters before further studies on the therapeutic effects. Furthermore, some recent studies have shown that hypoxia therapy may elicit positive effects on bone (Guner et al., 2013; Durand et al., 2014; Qiao et al., 2019). Consequently, we also investigated the bone formative potential of our IHT protocol on disuse osteopenic animals.
Therefore, the aim of the present study was to investigate the effects of normobaric IHT on the skeletal properties in both healthy and disuse osteopenic mice.
Materials and Methods
Animals and design
Thirty 14-week-old C57BL/6NRj (Janvier Labs, Le Genest-Saint-Isle, France) female mice were stratified by weight into four groups: Ctrl (n = 7), Botox (n = 7), Ctrl+IHT (n = 8), and Botox+IHT (n = 8). The animals were housed at 22°C with 50% humidity and a 12/12 hours day/night cycle with free access to water and food (Altromin 1324, Brogaarden, Lynge, Denmark).
At initiation of the study, the mice were anesthetized by inhalation with 4% isoflurane (IsoFlo Vet, Orion Pharma Animal Health, Copenhagen, Denmark) and had their right leg shaved and disinfected with 70% ethanol. Subsequently, Botox (2U/100 g BW; Allergan, Irvine, CA) was administered into the right quadriceps muscle (10 μl) and calf muscles (10 μl) of the mice in the Botox and Botox+IHT groups. The mice in the Ctrl and Ctrl+IHT groups were injected with a similar volume of saline.
IHT started on the day of the Botox injections and was performed in a normobaric hypoxic chamber developed to replace atmospheric oxygen by reducing oxygen through a nitrogen membrane unit (Fig. 1). During the experiment, the oxygen concentration in the hypoxia chamber was 10%. The animal cages were placed in the precalibrated hypoxic chamber for 1 hour at 8 am and 6 pm for 5 days/week. This exposed the mice to 1 hour of a constant oxygen-level of 10% twice a day interspersed with normal oxygen levels. The oxygen level was controlled by continuous oxygen measurements during the treatment period and by insertion of an external oxygen sensor (SmartSensor model AR8100; Graigar, Shenzhen, China) before all hypoxia-exposure periods. Temporary opening or closing of the small doors to the hypoxic chamber did not affect the overall oxygen level in the chamber. The adaptive response of mice to short-term exposure to hypoxia using this device has previously been presented (Bergholt et al., 2019).

The hypobaric hypoxia chamber.
To confirm Botox-induced paralysis of the hindlimb, the gait ability of the mice was assessed on day 2 using the score of Warner et al. (2006). In brief, the gait ability score assesses the ability of the mice to use their leg in five different ways, resulting in a score from 0 (completely disabled) to 10 (normal). The body weight of the mice was measured on day −7, 0, 3, 7, 10, 14, 17, and 21.
At the end of the study, the mice were anesthetized with 4% isoflurane and sacrificed by removal of the heart. The rectus femoris muscle was isolated and weighed. Tibiae and femora were isolated, and any remaining soft tissue was removed. The length of femur was measured with an electronic calliper before the bones were frozen at −20°C in Ringer's solution.
No animals died prematurely. The protocol was approved by The Danish Animal Experiments Inspectorate (Permit: 2018-15-0201-01436).
Dual-energy x-ray absorptiometry
The femora were placed in a pDEXA scanner (Sabre XL; Nordland Stratec, Pfortzheim, Germany) and the entire femur was scanned using a pixel size of 0.1 × 0.1 mm2 at a velocity of 3 mm/s. Bone mineral content (BMC) and aBMD were determined using the software provided with the scanner. Quality assurance was performed by scans of the two solid-state phantoms provided with the scanner. The coefficient of variation of mice femoral aBMD is 2.7% in our laboratory.
Micro-computed tomography
The right distal femoral metaphysis and femoral mid-diaphysis were scanned in a desktop micro-computed tomography (μCT) scanner (Scanco μCT 35; Scanco Medical AG, Brüttiselen, Switzerland). The scans were performed in high-resolution mode (1,000 projections/180°) with an isotropic voxel size of 3.5 μm, using an X-ray tube voltage of 55 kVp, a current of 145 μA, and an integration time of 800 ms. Beam-hardening effects were reduced using a 0.5 mm aluminum filter.
For analysis of the distal femoral metaphysis, a 1,000-μm high volume of interest (VOI) was used, starting 200 μm proximal to the most proximal part of the growth zone. The VOI was semi-interactively demarcated using the standard software supplied with the scanner (μCT Evaluation version 6.6) to include trabecular bone only.
For analysis of cortical bone, an 816-μm-high VOI was centered on the exact midpoint of the femoral mid-diaphysis. Demarcation of the periosteal surface of the bone was performed using the contour tool of the standard software supplied with the scanner.
Data were low-pass filtered using a Gaussian filter (σ = 0.8 and support = 1) and segmented with a fixed threshold filter. The threshold was determined with software supplied with the scanner (IPL, version 5.11; Scanco Medical AG, Brüttisellen, Switzerland) for each sample, and the median threshold was used for the evaluation of all samples. Thresholds for the distal metaphysis and mid-diaphysis were 525 and 574 mg HA/cm3, respectively. Quality assurance was performed by weekly (density) and monthly (geometry) scans of the solid-state calibration phantom provided with the scanner. The phantom contains five inserts of varying equivalent hydroxyapatite densities allowing the software supplied with the scanner to determine the density of the bone samples, following the scanner manufactures' protocol.
Mechanical testing
The femora were placed in a testing jig for three-point bending test with a fixed distance between the two supporting rods of 7.1 mm. Load was applied at a constant deflection rate of 2 mm/min with a third rod at the upper midpoint of the femur in a materials-testing machine (5566; Instron, High Wycombe, United Kingdom). During the mechanical testing, load-deflection data were recorded using Merlin (version 3.21; Instron) and subsequently analyzed using in-house developed software.
The proximal femur (the proximal half of the femur obtained after the three-point bending test) was mounted in a standard fixation device and placed in the material testing machine (Mosekilde et al., 1999). Load was applied to the top of the femoral head at a constant rate of 2 mm/min until fracture.
Statistical analysis
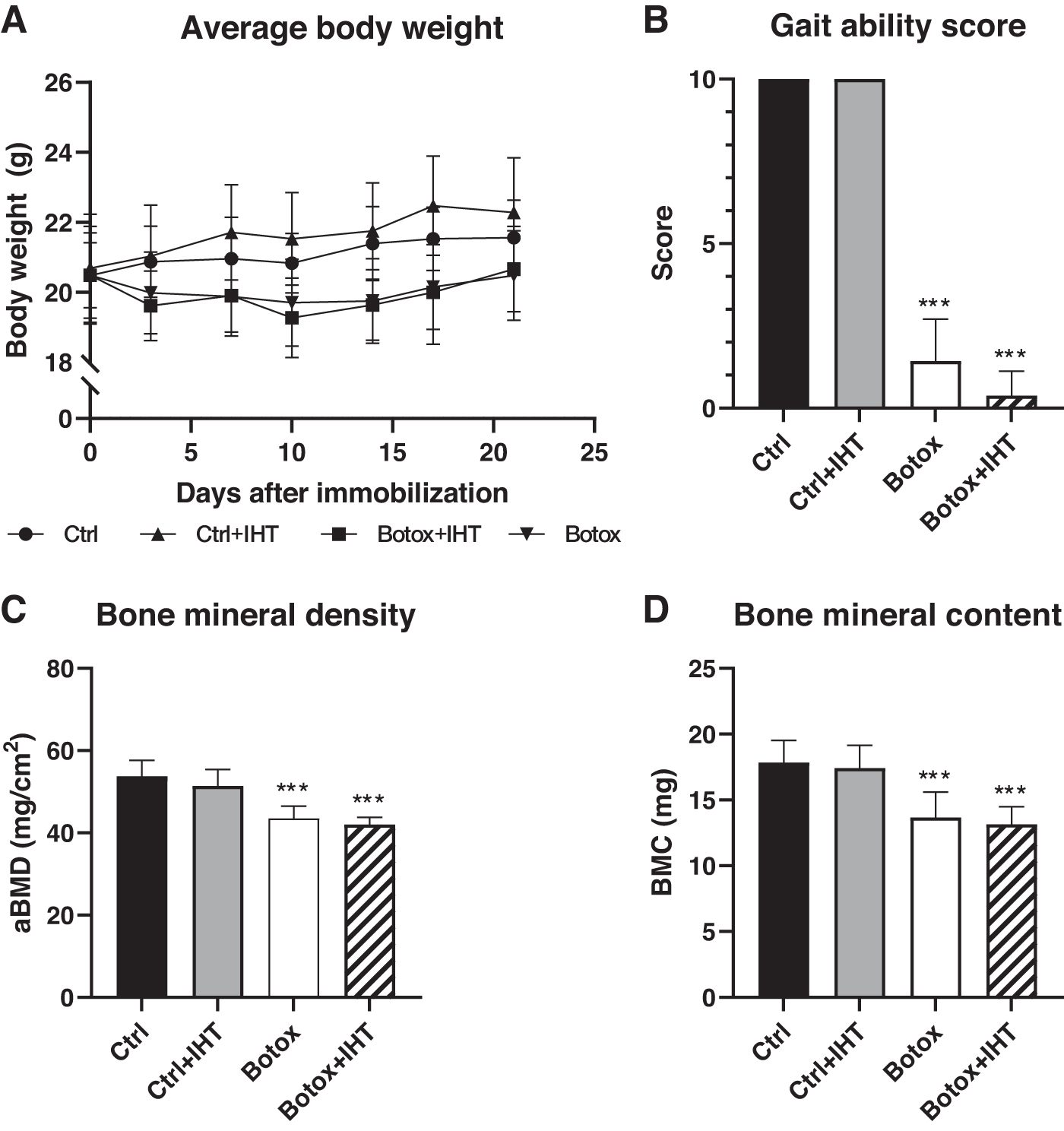
Data were analyzed with a parametric two-way analysis of variance (ANOVA; (Categorial variables: Ctrl vs. Botox and Normoxia vs. Hypoxia) performed on all groups, followed by a post hoc Holm-Sidak's multiple comparisons test. To analyze changes in body weight, a paired t-test was performed for initial and final body weight for each group. Results were considered statistically significant if p < 0.05. Data are presented as the relative difference between groups in text and as mean ± standard deviation in Tables 1, 2 and Figures 2, 4, and 6.

Distal femoral metaphysis bone parameters in control mice and mice with Botox-induced osteopenia treated with IHT.
Number of Animals, Initial and Final Body Weight, Femoral Bone Length, and Rectus Femoris Muscle Mass in Ambulating Mice and Mice with Botox-Induced Osteopenia Treated with Intermittent Hypoxia Therapy
Mean (SD).
p < 0.01 versus group initial BW.
p < 0.01 versus Ctrl.
p < 0.001 versus Ctrl.
IHT, intermittent hypoxia therapy; BW, body weight; RF, rectus femoris.
Properties of Trabecular and Cortical Micro-Structure at the Femoral Distal Metaphysis and the Femoral Mid-Diaphysis in Ambulating Mice and Mice with Botox-Induced Osteopenia Treated with Intermittent Hypoxia Therapy
Mean (SD).
p < 0.05 versus Ctrl.
p < 0.01 versus Ctrl.
p < 0.001 versus Ctrl.
BA, bone area; BV/TV, bone volume/tissue volume; CD, connectivity density; Ct.Th, cortical thickness; MA, marrow area; SMI, structure model index; TA, tissue area; Tb.N, trabecular number; Tb.Sp, trabecular spacing; Tb.Th, trabecular thickness; TMD, tissue mineral density; vBMD, volumetric bone mineral density.
Results
Body weight, gait ability score, and femoral length
The body weight did not differ between the groups at the study start (Fig. 2A). In both Ctrl and Ctrl+IHT mice, a significant increase in body weight was seen during the study. In contrast, no differences were found between initial and final body weight for Botox and Botox+IHT mice (Fig. 2A). A two-way ANOVA showed a significantly lower final body weight for disuse (Botox-injected) animals compared to control animals, however, a post hoc analysis did not reveal a significant difference between specific groups. IHT had no detrimental effect on the body weight in either Ctrl or Botox animals (Table 1).
Two days after the Botox injections, the gait ability score showed a significant loss of function of the right hindlimb of the Botox animals compared to their respective control animals (Fig. 2B). IHT did not influence the gait ability of the animals.
Botox did not affect femur length compared to respective Ctrl groups. Furthermore, IHT showed no detrimental effect on femoral length in either Ctrl or Botox mice, suggesting that IHT did not inhibit growth during the study (Table 1).
Muscle
Botox caused a significant loss of right rectus femoris muscle mass in Botox-injected mice compared to their respective control animals (Botox vs. Ctrl: −42% and Botox+IHT vs. Ctrl+IHT: −40%). IHT treatment did not alter rectus femoris muscle mass in either control or disuse osteopenic mice (Table 1).
Furthermore, Botox caused a significant reduction of the rectus femoris muscle mass in the uninjected left hindlimb compared to control animals (Botox vs. Ctrl: −14% and Botox+IHT vs. Ctrl+IHT: −16%).
Dual-energy x-ray absorptiometry
Animals injected with Botox showed a significant reduction in femoral aBMD compared to their respective control mice (Botox vs. Ctrl: −19% and Botox+IHT vs. Ctrl+IHT: −18%) (Fig. 2C). A similar significant decrease was found for femoral BMC for both Botox compared to Ctrl (−23%) and Botox+IHT compared to Ctrl+IHT (−24%) (Fig. 2D). Likewise, IHT did not affect aBMD or BMC for either Ctrl or Botox mice (Fig. 2C, D).
Micro-computed tomography
Distal femoral metaphysis
Disuse significantly decreased the density of the trabecular bone. Thus, trabecular bone volume fraction (bone volume/tissue volume [BV/TV]) was lost in both Botox (−42%) and Botox+IHT (−46%) animals compared to their respective control groups (Figs. 3 and 4). Similarly, volumetric bone mineral density (vBMD: Botox: −43% and Botox+IHT: −44%) and tissue mineral density (TMD: Botox: −3% and Botox+IHT: −4%) were significantly lower in disuse osteopenic mice than in control mice (Fig. 4). In addition, Botox significantly reduced trabecular thickness (Tb.Th) compared to the respective control groups (Tb.Th: Botox: −32% and Botox+IHT: −34%) (Fig. 4). A two-way ANOVA showed a significant increase in structure model index in disuse (Botox-injected) animals compared to control animals indicating transformation toward a more rod-like trabecular network, however, a post hoc analysis did not reveal a significant difference between specific groups. In contrast, connectivity density, trabecular number, and trabecular separation did not differ between disuse osteopenic and control animals (Table 2).

Three-dimensional reconstructed images of the proximal half of the femoral distal metaphysis from control mice and mice with Botox-induced osteopenia treated with IHT. The images presented are from the animal in each group with BV/TV closest to the group average.
IHT had no significant impact on the bone microstructure at the distal femoral metaphysis for either control or disuse osteopenic animals (Fig. 4 and Table 2).
Femoral mid-diaphysis
Botox resulted in a significant decrease in bone area at the femoral mid-diaphysis for both Botox (−9%) and Botox-IHT mice (−11%) compared to their respective controls (Fig. 5 and Table 2). In contrast, tissue area, marrow area, cortical thickness, and TMD did not differ between disuse osteopenic and control mice (Table 2).
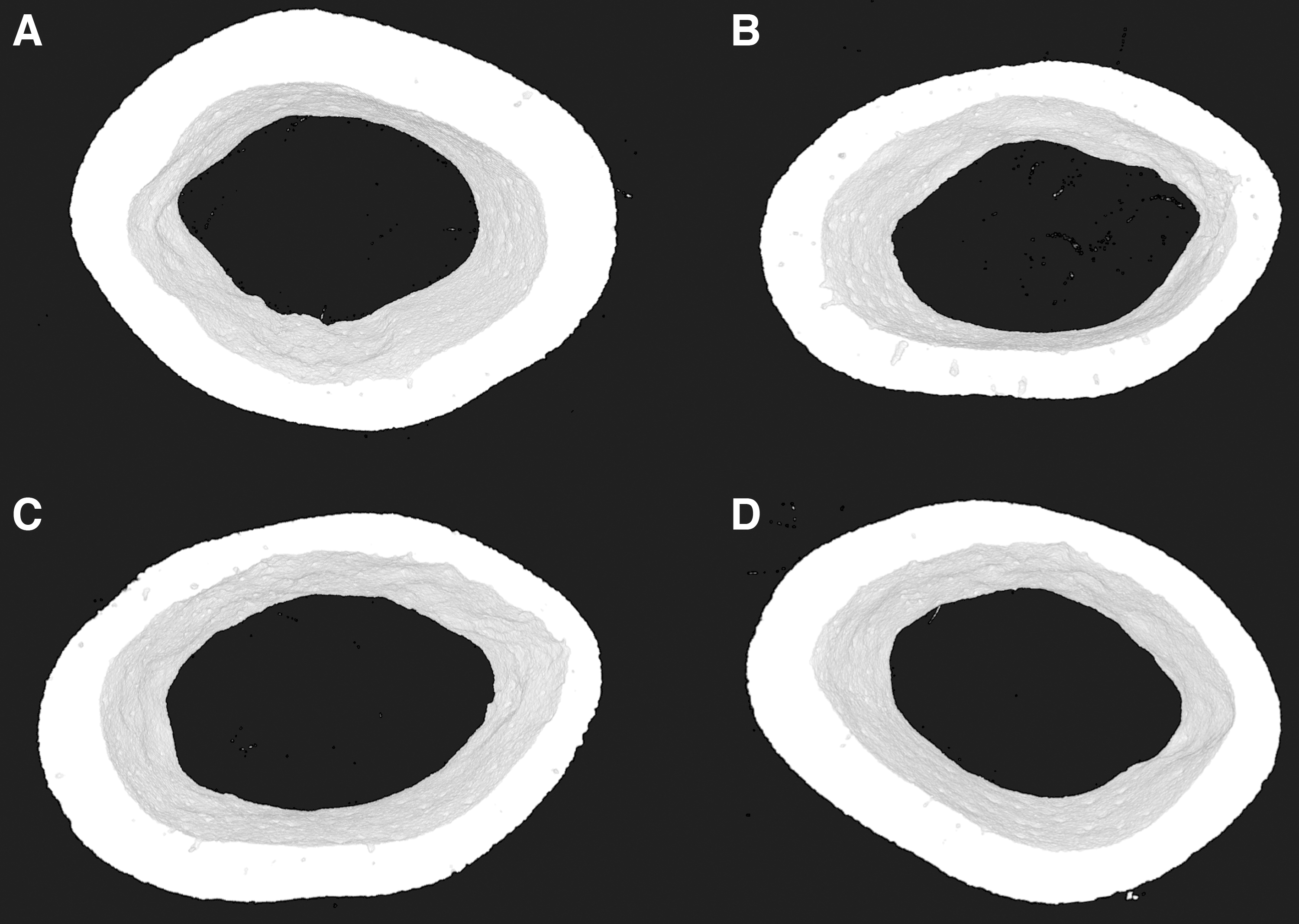
Three-dimensional reconstructed images of the femoral mid-diaphysis from control mice and mice with Botox-induced osteopenia treated with IHT. The images presented are from the animal in each group with average cross-sectional BA closest to the group average.
IHT had no significant impact on the femoral mid-diaphysis for either control or disuse osteopenic animals (Table 2).
Mechanical properties
Disuse significantly reduced femoral mid-diaphyseal bone strength compared to control animals in a two-way ANOVA analysis, but no difference between specific groups was found in the post hoc analysis (Fig. 6A). No detrimental effect of IHT on femoral mid-diaphyseal bone strength was found in any animals.

Bone strength (Fmax) of
In addition, disuse significantly decreased femoral neck strength in Botox (−55%) and Botox+IHT (−61%) mice compared to their respective controls (Fig. 6B). No detrimental effect of IHT on femoral neck strength was found in any animals.
Discussion
The present study found no effect of prolonged exposure to IHT on bone mass, trabecular microstructure, or bone strength in either control or disuse osteopenic mice.
Disuse resulted in a substantial loss of cortical and trabecular bone mass, Tb.Th, and femoral neck strength. Botox as a model of osteopenia has been tested extensively, and the findings of the present study are supported by those reported in previous studies (Warner et al., 2006; Grimston et al., 2007; Bloomfield, 2010; Brüel et al., 2013; Bettis et al., 2018; Brent et al., 2018).
In addition to the effect on the skeleton, Botox also caused large and significant muscle atrophy, which is in accordance with results from our laboratory and others (Warner et al., 2006; Lodberg et al., 2015; Bettis et al., 2018; Brent et al., 2018). The muscle atrophy was not only seen in the Botox-injected hindlimb but also in the contralateral hindlimb. In a previous study, we have shown that an additional Botox-injection 4 weeks after the primary injection caused a renewed paralysis, verified by decreased gait ability score, but that the additional Botox injection did not further affect either bodyweight or muscle mass of an uninjected muscle (Vegger et al., 2015). This suggests that the loss of body weight and muscle mass of the contralateral uninjected limb is not a direct effect of the Botox-injection, but may instead be due to the general stress inflicted on the animals by the paralysis. A similar reduction of body weight is seen in other models of disuse lending further credibility to the notion that the reduction in muscle mass and body weight is stress-induced (Mosekilde et al., 2000; Iwamoto et al., 2002; Knox et al., 2004). The Botox-induced muscle atrophy was not affected by treatment with IHT. Similarly, muscle mass in control animals was unaffected by IHT.
Our findings suggest that prolonged exposure to IHT has no negative effect on bone mass or mechanical and structural properties in control animals. Importantly, our findings indicate that application of IHT to treat diseases outside of the musculoskeletal system will not negatively affect bone mass, trabecular microstructure, or bone strength. Moreover, we established that IHT did not further exacerbate the harmful effects of Botox on bone mass and mechanical and structural properties. It can be argued that a potential positive effect of IHT on bone in Botox animals may be present, but is lost in the strong response to Botox. However, since we found a similar outcome in control animals, this is not likely the case.
The literature on the effect of IHT on bone is still sparse, and the few studies that exist show conflicting results. Guner et al. (2013) showed that IHT increased bone mineral density (BMD) in the lumbar spine in rats. They induced hypoxia by exposing the rats to a barometric pressure of 430 mmHg (corresponding to 4,500 m of altitude or a FiO2 of 11.9%) for 5 hours daily, 5 days/week for 5 weeks. In contrast, our finding that IHT did not alter the bone properties in control mice is supported by the findings in rats of Wang et al. (2016). They found no effects of a hypobaric IHT protocol, where they exposed rats to a barometric pressure of 404 mmHg (corresponding to 5,000 m of altitude or a FiO2 of 11.2%) for 4 hours daily for 14 consecutive days, on the bone properties in the femoral metaphysis in healthy animals, including BV/TV, BMC, BMD, and Tb.N (Wang et al., 2016). These findings are further supported by those of Durand et al., (2019) who demonstrated that IHT did not affect the loss of BV/TV in mice subjected to disuse by hindlimb suspension for 7 days. They placed their mice under hypobaric hypoxia conditions (375 mmHg corresponding to 5,575 m of altitude or a FiO2 of 10%) for four consecutive days starting at day 3 of hindlimb suspension (Durand et al., 2019).
In humans, Campo et al. (2015) found that 60 minutes of hypoxia training in athletes twice weekly did not alter BMD of the extremities, vertebral column, or total body. Furthermore, a study on hypoxia and bed rest in healthy young human males found that 21 days of hypobaric hypoxia (pressure corresponding to an altitude of 4,000 m or a FiO2 of 12.7%) did not further aggravate the loss of BMD due to bed rest (Rittweger et al., 2016).
The study by Wang et al., 2016 also investigated the effects of intermittent hypobaric hypoxia on estrogen deficient rats. The rats were ovariectomized (OVX) 4 weeks before initiation of the hypoxia therapy. Estrogen has an inhibitory effect on the bone-resorbing osteoclasts, and removal of the estrogen-producing ovaries leads to osteopenia and is a widely used model of postmenopausal osteopenia in rodents (Wronski et al., 1986; Arjmandi et al., 1996; Okazaki et al., 2002). They reported a detrimental effect of intermittent hypobaric hypoxia on bone in OVX rats, which resulted in a significant reduction of BMD, BMC, BV/TV, and Tb.N (Wang et al., 2016). These findings are in contrast to the findings of our study, where IHT did not lead to a further deterioration of the skeleton in disuse osteopenic mice. A reason for this difference may be that different models have been used to induce osteopenia. Thus, it is possible that IHT potentiates the effects of estrogen deficiency on the bone properties, while this may not be the case for disuse. This view may be further supported by the fact that two other studies of hypoxia found no exacerbation of immobilization-induced bone loss by chronic hypoxia (Rittweger et al., 2016; Durand et al., 2019). Another possible explanation for the different outcome is that Wang et al., 2016 studied rats, while we investigated mice in the present study. Finally, Wang et al., 2016 examined the effects of hypobaric hypoxia, while normobaric hypoxia was used in the present study. Further research in this area is needed especially concerning the effect of IHT in combination with estrogen deficiency.
As mentioned previously, two studies report that IHT improves bone fracture healing in mice (Durand et al., 2014) and rats (Qiao et al., 2019). One possible explanation for the different outcome of their and our study may be the essential role which vascularization plays in fracture healing (Bahney et al., 2015; Einhorn and Gerstenfeld, 2015). Hypoxia drastically increases vascularization through the HIF-1 pathway (Wan et al., 2010). Vascularization is imperative for fracture healing (Drager et al., 2015), but is not as relevant in disuse osteopenia, which is characterized by increased osteoclastic bone resorption and decreased osteoblastic bone formation.
Limitations
When developing an IHT protocol, important factors to consider are oxygen level, interval length, and interval frequency (Navarrete-Opazo and Mitchell, 2014). Thus, a limitation of our study is that we can only relate the bone response to our specific IHT regimen. Therefore, it cannot be precluded that other treatment regimens may affect bone differently. We know from previous studies that reduction of available oxygen to 10% is sufficient to cause an adaptive response in pathways for angiogenesis, metabolism, and inflammation in mice (Bergholt et al., 2019). However, in the present study, no control of the animal's response to hypoxia was performed.
Furthermore, research in obstructive sleep apnea (OSA), which can be seen as drastic IHT regimens with very low oxygen levels for short intervals, has linked OSA to decreased BMD in humans, but no conclusive data have been presented thus far (Swanson et al., 2015; Eimar et al., 2017). A study by Torres et al. (2013) simulating OAS reported no differences in BMD of an IHT regimen exposing mice to 6 hours/day of alternating 20 seconds of 5% oxygen and 40 seconds of 21% oxygen for 32 days. Based on these results, we surmise that while another IHT regimen may elicit either a negative or positive response on bone mechanical and structural properties, it will likely have to be much more aggressive than the one applied in the present study.
Conclusion
The findings of the present study indicate that the applied IHT regimen does not negatively affect bone mass, bone microstructure, or bone strength.
This suggests that treatment with IHT protocols similar to the one used in the present study, i.e., long-periods of moderate intermittent hypoxia as opposed to chronic hypoxia or short-term cyclic hypoxia, may cause no major adverse effect on bone in skeletally healthy individuals.
Footnotes
Author Contributions
Study design: F.D.B., A.B., C.B.F., J.S.T., and M.P. Study conduct: F.D.B., A.B., and C.B.F. Data collection, data analysis, and interpretation: F.D.B., A.B., J.S.T., and M.B.B. Article draft: F.D.B. Figures and graphical design: F.D.B. Article revision: all authors. Approval of final article: all authors.
Acknowledgments
The authors are thankful for the excellent technical assistance provided by Jytte Utoft and the animal care takes at Department of Clinical Medicine, Aarhus University. The μCT scanner was donated by the VELUX Foundation.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by Aarhus University Research Foundation NOVA grant (AUFF-E-2015-FLS-9-7).