
Abstract
Pooja, Vandana Sharma, Manish Sharma, Rajeev Varshney, Bhuvnesh Kumar, and Niroj Kumar Sethy. Association between 17β-estradiol receptors and nitric oxide signaling augments high-altitude adaptation of Ladakhi highlanders. High Alt Med Biol. 22: 174–183, 2021.
Background:
Genomic studies have identified positive natural selection of plasma membrane estrogen receptor signaling pathway for Himalayan highlanders. We sought to investigate significance of this pathway for high-altitude adaptation by studying Ladakhi highlanders.
Materials and Methods:
We recruited 25 healthy Ladakhi males (age range: 19–37, height: 164 ± 6 cm, and weight 59 ± 4 kg) at Leh (altitude 3,520 m) and age matched sea level volunteers at Delhi (altitude 215 m), India. We evaluated circulatory levels of 17β-estradiol (E2) and testosterone (T) and levels of E2 biosynthesis pathway proteins. In addition, we analyzed mRNA levels of E2 pathway genes and their association with nitric oxide (NO) availability.
Results:
We observed higher circulatory E2 and lower testosterone (T) in Ladakhi highlanders compared to lowlanders. Studying E2 pathway genes, we identified higher transcript levels of E2 receptors ESR1 (2.02-fold) and ESR2 (3.87-fold) in Ladakhi highlanders. Higher NOS3 mRNA, plasma level of endothelial NO synthase (eNOS), p-eNOS Ser1177, NOx (nitrate and nitrite), and cGMP were observed for Ladakhi highlanders. In addition, we observed a positive correlation between E2 with plasma NOx (r = 0.52, p = 0.002) and cGMP (r = 0.72, p = 0.007) for Ladakhi highlanders.
Conclusion:
Our results demonstrate higher circulatory E2 and lower T levels in Ladakhi highlanders. Higher levels of E2 and its receptors (ESR1 and ESR2) are positively associated with observed higher levels of eNOS signaling pathway metabolites. These results highlight the functional importance of E2 and its receptors for Himalayan pattern of high-altitude adaptation.
Introduction
Decreasing partial pressure of oxygen with increasing altitude limits alveolar and arterial oxygen availability, a condition termed as hypobaric hypoxia. Additional high altitude-associated environmental factors like cold, low humidity, and ultraviolet radiations challenge human survival and propagation. Despite this, more than 140 million people permanently reside at altitude above 2,500 m across the globe (Moore et al., 1998). Humans have colonized the Himalayas for more than 25,000 years (Aldenderfer, 2003) and these inhabitants exhibit higher resting ventilation, hypoxic ventilatory response, and higher levels of nitric oxide (NO) in exhaled breath and blood, reduced hypoxic pulmonary vasoconstriction, and lower pulmonary artery pressure and percent of oxygen saturation of hemoglobin unlike other high-altitude populations (Beall, 2006, 2007, 2014; Bigham et al., 2013). The other distinctive feature of Himalayan pattern of adaptation is average 3.5 g/dl less hemoglobin than Andeans (Beall and Reichsman, 1984; Winslow et al., 1989; Beall et al., 1998) that prevents risk of chronic mountain sickness (CMS), which is more profound in the Andes (Gazal et al., 2019).
Research over decades has highlighted the role of steroid hormones for high-altitude adaptation. Andean males with higher levels of testosterone (T) suffer from CMS, a lack of adaptation to altitude characterized by excessive erythrocytosis (Gonzales et al., 2009, 2011) and increased risk of developing cardiovascular events (Corante et al., 2018). Lower estradiol (E2) levels in postmenopausal Peruvian women are related to low pulse oxygen saturation (Gonzales and Villena, 2000). In contrast, Andean women exhibit higher levels of E2 during pregnancy compared to European counterparts at same altitude, possibly due to higher aromatase (CYP19A1) activity that converts testosterone to estrogen (Charles et al., 2014; Moore, 2017). As mentioned earlier, Himalayan highlanders possess lower hemoglobin levels compared to Andeans, and thus prevalence of CMS is uncommon at Himalayas (Xing et al., 2008; MacInnis and Rupert, 2011; Negi et al., 2013; Sahota and Panwar, 2013). Indigenous Himalayan inhabitants, Ladakhi women, possess higher circulatory estrogen levels compared to age-matched sea level women (Pooja et al., 2018). It is important to note that estrogen prevents excessive erythropoiesis and CMS in females at high altitude (Azad et al., 2020). Recent whole-genome sequencing combined with selection statistics studies has identified positive natural selection of E2 receptors (ESR1) and plasma membrane E2 receptor signaling pathway for Himalayan highlanders (Gnecchi-Ruscone et al., 2018), and positively associated with higher NO availability at Himalayas (Pooja et al., 2020). These observations emphasize population-specific regulation of steroidogenic hormones, especially T and E2, which confer adaptive features to native highlanders, although their precise role remains to be elucidated.
The steroid hormone estrogens confer protection to the cardiovascular system by increasing arterial vasodilation, inhibiting cellular response to vascular injury, and preventing atherosclerosis (Farhat et al., 1996; Mendelsohn and Karas, 1999). In the cardiovascular system, estrogen upregulates endothelial NO synthase (eNOS) transcription and also acutely activates PI3K signaling, leading to phosphorylation, activation of eNOS-mediated NO signaling, and arterial vasodilation (Chambliss and Shaul, 2002; Murphy, 2011). Estrogen mediates these effects by activation of estrogen receptors (ERs), ERα and ERβ, encoded by two distinct genes ESR1 and ESR2, which can alter gene transcription in the nucleus or acutely activate kinase signaling in the cytosol. In the classical signaling pathway, binding of the E2 to the ERs results in dimerization, interaction with co-activators, and nuclear localization followed by dimer binding to estrogen response elements (EREs) in gene promoters, leading to transcriptional activation of target genes (Hewitt et al., 2016). In addition, E2 also mediates rapid non-nuclear signaling events through pathways that involve transmembrane ERs, such as G protein-coupled ER 1 (GPER, formerly known as GPR30). This membrane-initiated steroid signaling mediates rapid activation of cytoplasmic or nuclear pathways by altering level of secondary messengers like calcium and cAMP (Prossnitz and Barton, 2011; Arnal et al., 2017).
In this study, we investigated E2 biosynthesis in an indigenous high-altitude population of the Himalayas, namely Ladakhis. We further analyzed the expression of E2 pathway genes and identified upregulation of E2 receptors ESR1 and ESR2 in Ladakhis. Using statistical analysis combined with protein-level validation, we report positive association of E2 with eNOS signaling pathway metabolites (NOx and cGMP). These results will aid in better understanding of role of E2 in human adaptation to high altitude, specifically Himalayan pattern of high-altitude adaptation.
Materials and Methods
Volunteers
The study recruited 25 healthy male volunteers born and brought up at Leh, Ladakh (Ladakhi, 3,250 m, age range: 19–37, height: 164 ± 6 cm, and weight 59 ± 4 kg), and 25 age-matched healthy volunteers residing at Delhi (Lowlander, 215 m). Volunteers with smoking habit and receiving antibiotic treatment, and lowlanders with history of high-altitude travel (above 2,500 m) in last 1 year were excluded. Written consent from all the volunteers was obtained after explaining the objectives and experimental procedures of the study. The Institutional Ethics Committee approved the study (IEC/DIPAS/B-6/2/2015), which is in accordance with Helsinki declaration. Overnight fasting venous blood was collected from the volunteers in EDTA vacutainers and PAXgene blood RNA tubes (Qiagen, Hilden, Germany) in the morning hours. Plasma was separated by centrifugation at 1,500 g for 15 minutes at 4°C and used fresh or stored in aliquots with protease and phosphatase inhibitor at −80°C for further use.
Determination of 17β-estradiol levels
The plasma 17β-estradiol levels were measured using estradiol enzyme-linked immunosorbent assay (ELISA) kit (EIA-2693; DRG International, NJ) as suggested by manufacturers. Briefly, 25 μl of plasma was added to 100 μl of enzyme conjugate and incubated at room temperature for 90 minutes with constant shaking. The plate was then washed and 100 μl of substrate was added to each well and incubated for 30 minutes in dark. Stop solution was added to terminate the reaction and absorbance was recorded at OD450 nm.
Determination of testosterone levels
The plasma levels of testosterone were estimated using testosterone ELISA kit (E-EL-0155; Elabscience) according to manufacturer's instructions. Briefly, 50 μl of biotinylated detection antibody was added to 50 μl plasma and incubated at 37°C for 45 minutes. Subsequently after washing, 100 μl of HRP conjugate was added and the plate was incubated at 37°C for 30 minutes. Followed by washing, 90 μl substrate was added and incubated again at 37°C for 15 minutes. Finally, stop solution was added and the absorbance was recorded at OD450 nm.
Determination of free testosterone levels
The plasma levels of free testosterone were estimated using Human Free Testosterone ELISA Kit (CSB E05096h; Cusabio, Wuhan) according to manufacturer's instructions. Briefly, 50 μl each of plasma, HRP conjugate, and biotinylated detection antibody was added and incubated at 37°C for 60 minutes. After washing, 50 μl of substrate A and B was added and the plate was incubated at 37°C for 15 minutes in dark. Stop solution was added to terminate the reaction and absorbance was recorded at OD450 nm.
Determination of human sex hormone-binding globulin
The plasma levels of sex hormone-binding globulin (SHBG) were estimated using Human SHBG ELISA Kit (CSB E08232h; Cusabio) according to manufacturer's instructions. Plasma samples were incubated with 100 μl of biotinylated detection antibody at 37°C for 60 minutes. After washing, 100 μl of HRP-avidin was added and incubated again at 37°C for 60 minutes. The plate was subsequently washed and 90 μl of TMB substrate was added, followed by incubation for 15 minutes in dark. Stop solution was added and the absorbance was recorded at OD450 nm, and the concentrations were inferred with reference to a standard curve.
Determination of albumin levels
The plasma levels of albumin were evaluated using human albumin ELISA kit (E-EL-H6105; Elabscience) as per manufacturer's recommendations. Briefly, 100 μl of diluted plasma samples (1:100,000 v/v) was added to the precoated wells and incubated at 37°C for 90 minutes. The wells were decanted and 100 μl of biotinylated detection antibody was added to each well and incubated at 37°C for 60 minutes. After washing, 100 μl of HRP conjugate was added and again incubated at 37°C for 30 minutes. The plate was washed and incubated with 90 μl of substrate at 37°C for 15 minutes in dark. Stop solution was added to terminate the reaction and the absorbance was recorded at OD450 nm, and the concentrations were inferred with reference to a standard curve.
RNA isolation, cDNA synthesis, and polymerase chain reaction
Total RNA was extracted from PAXgene blood RNA tubes using PAXgene Blood RNA kit (Qiagen, Hilden, Germany) according to manufacturer's instructions. The concentration of the isolated RNA was estimated by recording OD260/280 ratio. The integrity of isolated RNA was also checked by agarose gel electrophoresis. Using high-capacity cDNA reverse transcription kit (Applied Biosystems, Foster City, CA), 500 ng of DNase I-treated RNA was reverse transcribed to cDNA following manufacturer's recommendations. The mRNA levels of NOS3 and GAPDH were evaluated by semiquantitative real time-polymerase chain reaction as mentioned earlier (Pooja et al., 2018). The polymerase chain reaction (PCR) products were resolved on agarose gels stained with ethidium bromide and images were captured on gel documentation system (BIO-RAD Molecular Imager® Gel Doc™ XR + Imaging System; Bio-Rad, Inc., Laboratories).
Human estrogen signaling PCR array
Expression analysis of estrogen pathway genes was evaluated using Human estrogen signaling TaqMan™ array (Cat no. 4414087, 92 target genes; ThermoFisher Scientific). This PCR array contains individual TaqMan assays for 92 genes associated with estrogen signaling. Three samples from each group (Lowlander and Ladakhi) were analyzed on StepOnePlus™ real-time PCR system using TaqMan Fast Advanced Master Mix (Cat no. 4444963; ThermoFisher Scientific). The results were normalized and expression values were obtained using comparative Cτ method (ΔΔCτ).
Plasma NOx measurement
The NOx level was determined in plasma samples using nitrate/nitrite Colorimetric Assay Kit (780001; Cayman Chemical, Ann Arbor, MI) according to manufacturer's recommendations. Briefly, plasma sample was diluted with assay buffer (1:1, v/v) for NOx determination. The absorbance was recorded at OD540 nm and the concentrations were inferred with reference to a standard curve.
Evaluation of cGMP levels
The plasma cGMP concentrations were determined using Cyclic GMP ELISA Kit (581021; Cayman Chemical) according to manufacturer's instructions. Briefly, 2 ml of ice-cold ethanol was added to 500 μl of plasma. The precipitate was centrifuged at 1,500 g for 10 minutes and supernatant was separated. The collected supernatant was vacuum dried and reconstituted for cGMP estimation. The absorbance was recorded at OD412 nm and the concentrations were determined with reference to a standard curve.
Western blotting
The protein and phosphoprotein levels of eNOS along with E2 biosynthetic pathway proteins cytochrome P450 family 11 subfamily A member 1 (CYP11A1/cholesterol side-chain cleavage enzyme, mitochondrial), cytochrome P450 family 17 subfamily A member 1 (CYP17A1/steroid 17-alpha-hydroxylase/17,20 lyase), and cytochrome P450 family 19 subfamily A member 1 (CYP19A1/aromatase) were estimated in plasma samples by Western blot analysis. Briefly, plasma proteins (30 μg) were separated on 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to nitrocellulose membrane. The membranes were blocked overnight using 5% nonfat dried milk (M7409; Merck) in PBST/TBST, and incubated anti-eNOS (21170; Signalway antibody), anti-phospho-eNOS Ser1177 (9570; Cell Signaling Technologies), anti-phospho-eNOS Thr495 (9574; Cell Signaling Technologies), anti-CYP11A1 (GTX56293; GeneTex), anti-CYP17A1 (GTX56294; GeneTex), anti-CYP19A1 (E-AB-31086; Elabscience), and anti-tubulin (T9026; Sigma-Aldrich) for 2 hours at room temperature. The membranes were washed thrice for 5 minutes each with 0.1% Tween20 in 1 × phosphate-buffered saline (PBS). Secondary antibodies were added to the blots and incubated for 1 hour at room temperature on constant shaking. The membranes were washed using 1 × PBS with 0.1% Tween20 and were finally developed using chemiluminescent substrate (34095; ThermoFischer Scientific). The images were captured using BioSpectrum 515 Imaging System (UVP, Upland, CA) and analyzed by Image J software.
Statistical analysis
Data were analyzed with GraphPad Prism version 7.0 (San Diego, CA) for Pearson correlation analysis and t tests. Results were plotted as mean ± SEM and p < 0.05 was considered statistically significant.
Results
Enhanced estradiol biosynthesis in Ladakhi highlanders
We estimated the levels of plasma E2, T, T/E2 ratio, free T/total T ratio, and SHBG and albumin levels in both the studied groups. The E2 levels were found to be 1.39-fold higher in Ladakhis (p < 0.01) than lowlanders (Fig. 1a). In corroboration, total T levels were found to be 0.66-fold lower in Ladakhis (p < 0.01) compared to lowlanders (Fig. 1b). Consequently, Ladakhis exhibited 0.46-fold lower T/E2 ratio (p < 0.001) compared to lowlanders (Fig. 1c). We observed 1.61-fold (p < 0.01) higher levels of free T (Fig. 1d) and 2.19-fold (p < 0.01) higher ratio of free T to total T (Fig. 1e) in Ladakhis compared to lowlanders. Interestingly, the SHBG levels was 3.48-fold (p < 0.001) higher in Ladakhis (Fig. 1f), whereas albumin levels were 0.58-fold (p < 0.001) lower compared to lowlanders (Fig. 1g). To ascertain E2 biosynthesis, we evaluated the levels of CYP11A1, CYP17A1, and CYP19A1. Ladakhi highlanders exhibited significantly higher levels of CYP11A1 (1.21-fold, p < 0.05), CYP17A1 (1.3-fold, p < 0.01), and CYP19A1 (1.16-fold, p < 0.05) compared to lowlanders (Fig. 1h, i).

Evaluation of 17β-estradiol (E2) biosynthesis in Ladakhi highlanders compared to lowlanders. Estimation of
Expressions of E2 pathway genes are higher in Ladakhis
PCR array-based transcriptional profiling of E2 pathway genes revealed higher transcript levels of ESR1 (2.02-fold, p < 0.05), ESR2 (3.87-fold, p < 0.001), hydroxysteroid 17-beta dehydrogenase 8 (HSD17B8; 1.52-fold), HSPB8 (4.25-fold, p < 0.001), peroxisome proliferative activated receptor gamma (PPARG) coactivator 1 alpha (PPARGC1A; 3.66-fold, p < 0.001), RNA binding fox-1 homolog 2 (RBM9; 3.69-fold, p < 0.001), steroid sulfatase (STS; 2.14-fold, p < 0.01), vasoactive intestinal peptide (VIP; 4.01-fold, p < 0.01), and androgen receptor (AR; 4.06-fold, p < 0.01), whereas lower transcript levels of DEAD-box helicase 54 (DDX54; 0.63-fold), scaffold attachment factor B (SAFB; 0.13-fold), and scaffold attachment factor B2 (SAFB2; 0.50-fold) were observed for Ladakhi highlanders compared to lowlanders (Fig. 2).
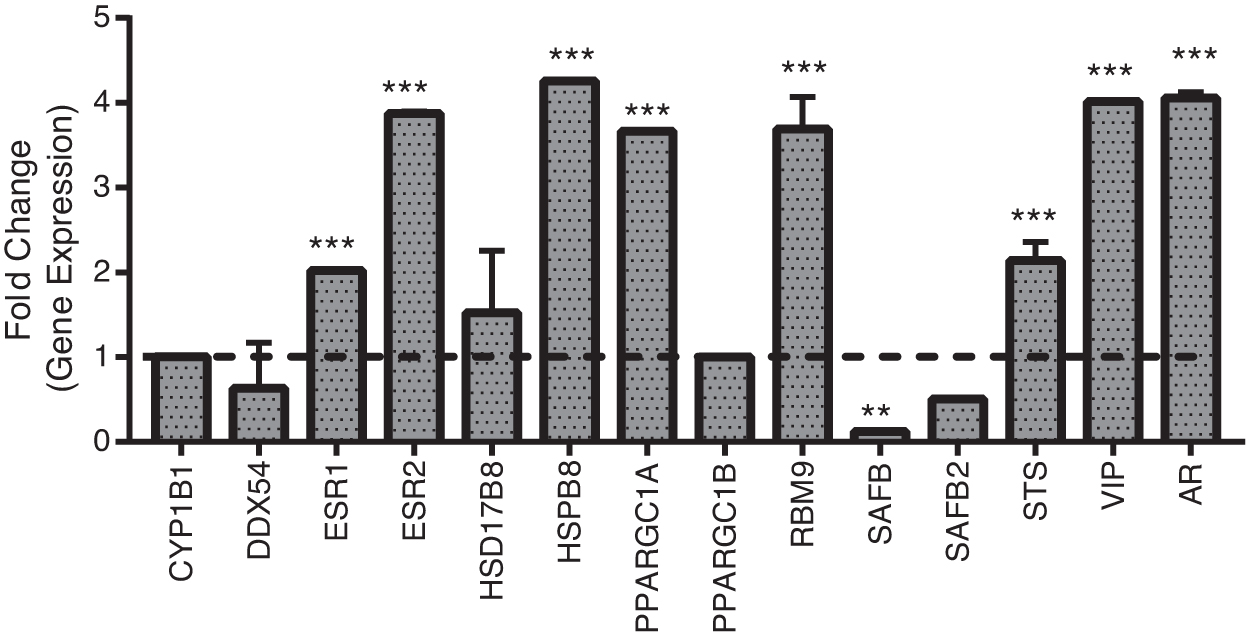
Real-time polymerase chain reaction-based transcriptional profiling of E2 pathway. The bar graph represents fold change in genes, which regulates estradiol levels in Ladakhis with respect to lowlanders. Results are presented as mean ± SEM of three independent experiments (n = 3 per group). **p < 0.01 and ***p < 0.001 compared to lowlanders. AR, androgen receptor; CYP1B1, cytochrome P450 family 1 subfamily B member 1, DDX54, DEAD-box helicase 54; ESR1, estrogen receptor 1; ESR2, estrogen receptor 2; HSD17B8, hydroxysteroid 17-beta dehydrogenase 8; PPARGC1A, PPARG coactivator 1 alpha; PPARGC1B, PPARG coactivator 1 beta; RBM9, RNA binding motif protein 9/RNA binding fox-1 homolog 2; SAFB, scaffold attachment factor B; SAFB2, scaffold attachment factor B2; STS, steroid sulfatase; VIP, vasoactive intestinal peptide.
Enhanced eNOS-mediated NO production in Ladakhi
To evaluate eNOS-mediated NO production and downstream signaling, we measured mRNA and protein levels of NOS3/eNOS in Ladakhi highlanders. Our PCR-based studies revealed 1.39-fold (p < 0.05) higher NOS3 mRNA in Ladakhis compared to lowlanders (Fig. 3a, b). We also observed 2.47-fold (p < 0.001) higher eNOS protein in Ladakhi highlanders. In corroboration, we observed 1.29-fold (p < 0.05) higher p-eNOS Ser1177 along with 0.78-fold (p < 0.05) lower levels of p-eNOS Thr495 in Ladakhis compared to lowlanders (Fig. 4a, b). Evaluating plasma NOx, we observed 2.64-fold (p < 0.001) higher levels in Ladakhis compared to lowlanders (Fig. 4c). We also observed 1.79-fold (p < 0.01) higher cGMP levels in Ladakhis compared to lowlanders (Fig. 4d).
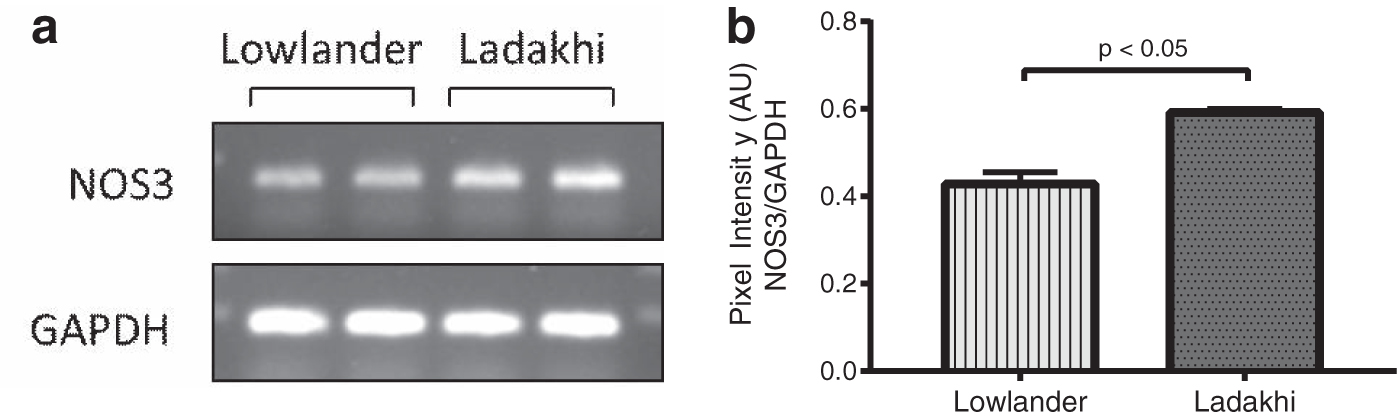
Enhanced NOS3 mRNA expression in Ladakhi highlanders.
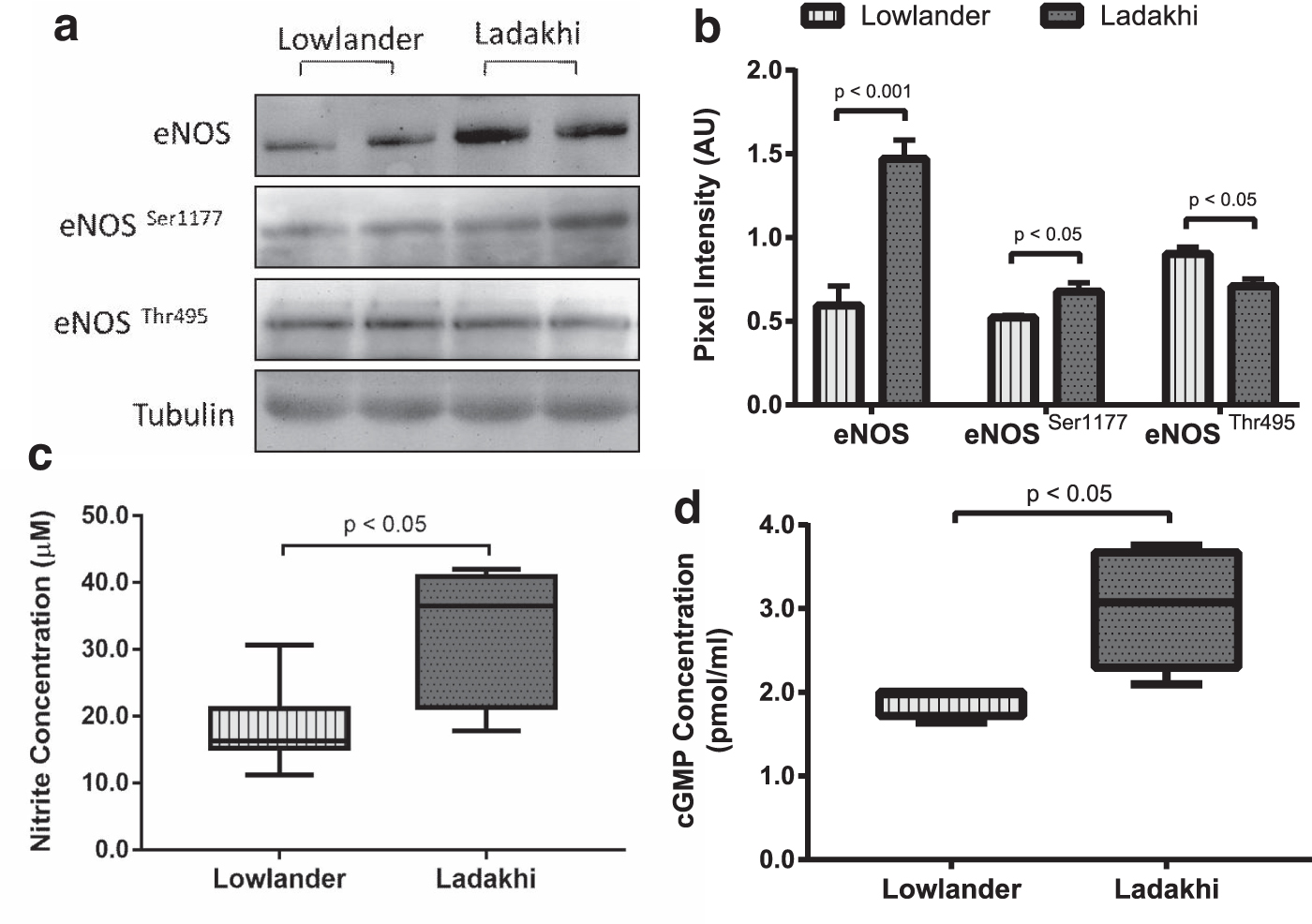
Ladakhis exhibit enhanced eNOS-NO-cGMP signaling.
Correlation between E2 and NO metabolites
Evaluating correlation between E2 and NO metabolites (NOx and cGMP), we observed moderate positive association between E2 and NOx (r = 0.52, p < 0.01). In addition, we also observed moderate positive correlation between E2 and cGMP (r = 0.72, p < 0.01) for Ladakhis (Table 1). In contrast, no such association was observed for lowlanders.
Pearson Correlation Analysis Between Plasma E2 and Nitric Oxide Metabolites (NOx and cGMP) for Lowlanders and Ladakhi
Data were analyzed with GraphPad Prism Version 7.0 (San Diego, CA). R indicates correlation coefficient and p represents level of significance.
Discussion
Humans inhabiting high-altitude regions across the world have undergone positive natural selection over the generations. The indigenous populations in the Himalayas, the Andean Altiplano, and the Semian Plateau have evolved adaptive physiological responses to limit negative effects of hypoxia. The Himalayas have the longest human inhabitation history and consequently had more time and opportunity for natural selection in response to hypoxic environment than any other high-altitude human population (Petousi and Robbins, 2014). The striking physiological features of Himalayan or Tibetan pattern of high-altitude adaptation include comparative lower hemoglobin levels and prevalence of CMS unlike Andeans. Other notable physiological adaptations include higher levels of the vasodilator NO in the lungs and blood of Tibetans, which is consistent with their lower pulmonary artery pressure (Beall, 2006, 2007, 2014).
Beyond these phenotypic observations, genetics-based investigations have identified positive natural selection of estrogen signaling pathway (Gnecchi-Ruscone et al., 2018) and underlying gene CYP17A1 involved in the synthesis of steroid hormones (Simonson et al., 2010; Wuren et al., 2014) for Himalayan highlanders. The steroid hormone estrogens are pivotal for development and maintenance of normal sexual and reproductive function, as well as have pleiotropic effects on the cardiovascular system (Mendelsohn and Karas, 1999). Estradiol, also known as 17β-estradiol or estrogen (E2), is the most abundant form of circulating estrogens and is synthesized from cholesterol along with estrone (E1) and estriol (E3) by series of reactions involving cytochrome P450 family (CYPs). The first and rate-limiting step in the synthesis of the steroid hormones is the conversion of cholesterol to pregnenolone at inner membrane of mitochondria. This is mediated by a cholesterol side-chain cleavage enzyme (CYP11A1/P450scc) located at mitochondria. Furthermore, pregnenolone and progesterone are converted to dehydroepiandrosterone and androstenedione, respectively, by action of CYP17A1 (steroid 17-alpha-hydroxylase/17,20 lyase). The last and rate limiting step in E2 biosynthesis is mediated by the enzyme aromatase (CYP19A1) that converts testosterone to E2 (Payne and Hales, 2004; Miller and Auchus, 2011). In addition to gonadal sites, E2 is also synthesized at adrenal glands, brain, adipose tissue, skin, pancreas, and blood vessels and acts locally in a paracrine and intracrine manner (Simpson et al., 2002).
Studying Ladakhi highlanders, we observed higher circulatory E2 levels as well as higher levels of E2 biosynthetic pathway proteins CYP11A1, CYP17A1, and CYP19A1 compared to lowlanders. Concomitantly, we also observed lower total testosterone levels and T/E2 ratio in Ladakhi highlanders, although free testosterone levels were higher compared to lowlanders. Under physiological conditions, ∼70% of testosterone is bound to SHBG with high affinity, 20%–30% is weakly bound to albumin, and 1%–2% remains free (Dunn et al., 1981; Westphal, 1986). Free testosterone along with albumin-bound testosterone can be aromatized by aromatase/CYP19A1 for E2 synthesis. It is noteworthy that the binding affinity of testosterone to SHBG is almost threefold higher than E2 and higher SHBG levels result in enhanced E2 biosynthesis (Burke and Anderson, 1972). Our present results provide multiple evidences for enhanced E2 availability for Ladakhi highlanders, including higher levels of key E2 biosynthetic pathway proteins, including aromatase, higher SHBG and lower albumin levels, higher free testosterone levels that serve as substrate for aromatase, and higher circulatory E2 levels. These observations also corroborate genomic studies reporting positive natural selection of steroid biosynthesis genes, estrogen pathway and ERs for Himalayan pattern of high-altitude adaptation (Simonson et al., 2010; Wuren et al., 2014; Gnecchi-Ruscone et al., 2018). Higher E2 levels with concomitant lower testosterone levels may explain lower hemoglobin level, higher vasodilation, and reduced CMS prevalence at high altitude compared to Andean populations. Recent studies of Azad et al. reporting E2-mediated prevention of excessive erythropoiesis and Monge's disease at high altitude (Azad et al., 2020) further supports our present observations.
E2 mediates its action through two predominant mechanisms, genomic and rapid nongenomic pathways. In the genomic pathway, E2 diffuses into the cell and binds the ERs (ERα/ESR1 and ERβ/ESR2), which form homodimers or heterodimers and then bind to EREs (defined as GGTCAnnnTGACC) in the promoter region of estrogen-responsive genes. Several coactivators (e.g., NCOA1, NCOA2, NCOA3, CREBBP, PPARBP, P68, and SRA) and corepressors (e.g., NCOR1 and NRIP) reportedly interact with ligand-bound ERα or ERβ and influence the ability of the receptor to activate or inhibit gene expression. Beside this, ERs can also trigger faster responses through plasma membrane receptors activating nongenomic signaling (Deschamps et al., 2010; Prossnitz and Barton, 2011; Arnal et al., 2017). Our gene expression studies identified activation of E2 receptor genes ESR1 and ESR2 in Ladakhi highlanders. Studies on animal models have reported hypoxia-induced upregulation of ESR1 (Wu et al., 2008) and ESR2 genes in the lung and pulmonary vasculature (Frump et al., 2018). A recent transcriptomic study using a rat model mimicking the human condition of cardiopulmonary adaptation to high altitude has identified activation of ESR1 in the lung and right ventricle (Krishnan et al., 2020). In humans, disruptive mutation of ESR1 abolishes flow-mediated endothelium-dependent vasodilation of brachial artery in humans (Sudhir et al., 1997b) and accelerates coronary arteriosclerosis (Sudhir et al., 1997a). These studies along with our present results corroborate the importance of E2-ER pathways in endothelium-dependent vasodilation and high-altitude adaptation. We also identified higher transcript level of several ER coactivators like PPARGC1A (Tcherepanova et al., 2000) and RBM9/RBFOX2. In contrast, we observed lower transcript level of corepressors like DDX54, SAFB, and SAFB2, further indicating activation of E2 signaling pathway. We have also identified higher mRNA levels of VIP, a downstream target of E2 (Lam et al., 1990) involved in vasodilation in humans (Kellogg et al., 2010). These cumulative results further support enhanced E2 availability and E2-ESR1-/ESR2-mediated signaling conferring vascular adaptive features to Ladakhi highlanders.
Epidemiological data, experimental and clinical evidence, have demonstrated that E2 regulates cardiovascular physiological function and features by increasing NO availability (Mendelsohn and Karas, 1999; Vitale et al., 2009; Pooja et al., 2020). E2 also plays a crucial role in the ability of resistance arteries to remodel in response to long-term increase in blood flow, which is necessary to optimize tissue perfusion (Tarhouni et al., 2014). ERα in the endothelium is involved in the beneficial vascular actions of E2, which increases basal NO production by both genomic and nongenomic pathway (Darblade et al., 2002; Arnal et al., 2010; Guivarc'h et al., 2018). Ligand-activated ERs bind to the ERE present in the eNOS promoter and activate gene transcription (MacRitchie et al., 1997). The nongenomic pathways include eNOS phosphorylation through the phosphatidylinositol-3 kinase/protein kinase B pathway, promoting NO release (Caulin-Glaser et al., 1997; Dimmeler et al., 1999; Haynes et al., 2000; Hisamoto et al., 2001; Simoncini et al., 2002). We observed 1.38-fold higher NOS3 mRNA levels in Ladakhis, indicating higher eNOS transcription. Similarly, we observed higher levels of eNOS and phospho-eNOS Ser1177, plasma biomarkers for NO production (NOx) and signaling (cGMP), in Ladakhi highlanders compared to lowlanders. It is important to note that we observed lower levels of phospho-eNOS Thr495 in Ladakhis, similar to our previous observations in Ladakhi women (Pooja et al., 2018). Phosphorylation of eNOS at Ser1177 and simultaneous dephosphorylation at Thr495 augment eNOS activity (Vanhoutte et al., 2016). All these studies, along with our present observations, suggest that increased availability of E2 and activation of E2 receptors stimulate eNOS-mediated NO production in Ladakhi natives. This observation was further supported by positive correlations between E2 and NO biomarkers (NOx and cGMP), supporting critical role of E2 in augmenting NO availability at Himalayas.
In vascular system, regulatory action of E2 has been implicated in the release of additional endothelial-derived molecules such as prostacyclin and angiotensin (Ang) 1–7, and decrease in endothelin-1 bioavailability and Ang II receptor type 1 (AT1) expression, thus reducing vasoconstriction and promoting vasodilation (Pérez-Cremades et al., 2018). Further studies, including diverse Himalayan populations, may precisely elucidate the specific role of E2 in regulating these vasodilator and vasoconstrictor molecules for high-altitude adaptation. Along with cardiovascular system, E2 is involved in many nonsexual physiological processes, including regulation of skeletal homeostasis, lipid and carbohydrate metabolism, electrolyte balance, and central nervous system function (including cognition and behavior) (Pérez-Cremades et al., 2018). Further studies are warranted to understand the effects of higher E2 levels on these physiological processes at high altitude. Emerging evidence suggests that epigenetic estrogen-regulation mechanisms are implicated in E2-mediated regulation of cardiovascular function (Post et al., 1999; Pérez-Cremades et al., 2018). Identification of these epigenetic mechanisms for E2 regulation in high-altitude populations may augment current understanding of high-altitude adaptation. It is important to note that relative balance between ERα and ERβ expression in target organs selectively regulate E2 signaling. These receptor subtypes can also have opposing gene expression regulatory effects, in addition to their redundant mediatory roles (Arnal et al., 2017). Although we have not delineated the specific role of each receptor in this study, their upregulation in Ladakhi highlanders suggests major roles of these receptors for high-altitude adaptation. We studied only young Ladakhi male volunteers for several reasons. First, E2 concentrations widely vary depending on gender and age that result in gender- and age-specific physiological functions, notably enhanced cardiovascular protection in premenopausal women (Vitale et al., 2009; Pérez-Cremades et al., 2018). Second, transgenic and knockout animal models of E2 pathway genes, including ESR1, ESR2, and aromatase, have also revealed gender-specific physiological functions (Rubanyi et al., 1997; Cooke and Nanjappa, 2017). Third, it is extremely difficult to recruit menstrual cycle phase-matched high-altitude native female volunteers. However, such information for both the genders will supplement the current understanding of high-altitude acclimatization.
Conclusion
This study reports that Ladakhi highlanders possess elevated E2 levels with concomitant lower testosterone levels and T/E2 ratio. Higher levels of E2 biosynthesis pathway proteins CYP11A1, CYP17A1, and CYP19A1 provide evidence for increased circulatory E2 availability. Our transcript profiling studies demonstrated activation of E2 receptors (ESR1 and ESR2) with concomitant lower levels of corepressors DDX54, SAFB, and SAFB2, further supporting augmented E2 biosynthesis. We also observed higher levels of NO metabolites and activation of eNOS-NO-cGMP pathway in Ladakhi highlanders. More importantly, we observed a moderate positive correlation of E2 with eNOS pathway metabolites providing molecular insights for higher vasodilation observed in the Himalayas, unlike Andean populations. These results expand our current understanding for Himalayan pattern of high-altitude adaptation and suggest that steroidogenic hormones play a major role for human adaptation to high altitude.
Footnotes
Author's Contributions
Pooja and N.K.S. conceived and planned the experiments. Pooja, V.S., M.S., and N.K.S. carried out the experiments at sea level and high altitude. N.K.S., R.V., and B.K. contributed to the interpretation of the results. N.K.S. took the lead in writing the article. All authors provided critical inputs and consent for the article.
Acknowledgment
The authors acknowledge Director, DIHAR, DRDO, Leh, for providing laboratory facilities to carry out experiments at high altitude. The authors also acknowledge The Indian Army for facilitating this study. Pooja is recipient of Senior Research Fellowship from University Grants Commission (UGC). V.S. is recipient of fellowship from DRDO, Delhi, India.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work is supported by DRDO, Ministry of Defence, Government of India (Grant number DIP-263).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.