
Abstract
Baranauskas, Marissa N., Joseph Powell, Alyce D. Fly, Bruce J. Martin, Timothy D. Mickleborough, Hunter L. Paris, and Robert F. Chapman. Influence of zinc on the acute changes in erythropoietin and proinflammatory cytokines with hypoxia. High Alt Med Biol. 22: 148–156, 2021.
Background:
Considerable, unexplained, interindividual variability characterizes the erythropoietin (EPO) response to hypoxia, which can impact hematological acclimatization for individuals sojourning to altitude. Zinc supplementation has the potential to alter EPO by attenuating increases in inflammation and oxidative stress. Yet, the application of such an intervention has not been evaluated in humans. In this proof-of-concept study, we aimed to evaluate the EPO and inflammatory responses to acute hypoxia in human participants following chronic zinc supplementation.
Methods:
Nine physically active participants (men n = 5, women n = 4, age 28 ± 4 years, height 176 ± 11 cm, mass 77 ± 21 kg) were exposed to 12 hours of normobaric hypoxia simulating an altitude of 3,000 m (FiO2 = 0.14) before and after 8 weeks of supplementation with 40 mg/day of elemental zinc from picolinate. Blood samples for subsequent analysis of serum zinc, EPO, superoxide dismutase (extracellular superoxide dismutase [EC-SOD]), C-reactive protein (CRP), and proinflammatory cytokines were obtained pre- and postsupplementation and exposure to hypoxia.
Results:
After zinc supplementation, EPO increased by 64.9 ± 36.0% (mean ± standard deviation) following 12 hours of hypoxia, but this response was not different from presupplementation (70.8 ± 46.1%). Considerable interindividual (range: −1% to +208%) variability was apparent in the acute EPO response. While most markers of inflammation did not change with hypoxia, interleukin-6 concentrations increased from 1.17 ± 0.05 to 1.97 ± 0.32 pg/ml during the final 6 hours. The acute EPO response at 12 hours was not related to changes in serum zinc, EC-SOD, CRP, or proinflammatory cytokines.
Conclusions:
Zinc supplementation does not influence the acute EPO or inflammatory response with short-term exposure to moderate levels of normobaric hypoxia (3,000 m) in apparently healthy young adults.
Introduction
Altitude training practices, defined as the process of acclimating for ≥12 hours/day at terrestrial or simulated altitudes of 2,500–3,000 m for ≥18 days (Levine and Stray-Gundersen, 1997; Brugniaux et al., 2006; Ploszczyca et al., 2018), have been adopted by endurance athletes, mountaineers, and military personnel for the purpose of eliciting physiological adaptations beneficial to physical performance at altitude and sea level (Wilber, 2007; Heil and Keenan, 2014). Within the first several hours of exposure to hypoxia, erythropoietin (EPO) activity acutely increases (Eckardt et al., 1989; Ge et al., 2002; Friedmann et al., 2005; Mounier et al., 2006) in response to reductions in arterial oxygen content (Montero and Lundby, 2019). Due to its rapid and marked response, EPO may have potential as a useful biomarker in predicting how an individual may respond to altitude training (i.e., improved physical performance) (Chapman et al., 1998).
However, the apparent relationships between the acute EPO response and subsequent hematologic adaptations (i.e., increased hemoglobin mass) following altitude training are likely precluded by interindividual variability in both responses (Friedmann et al., 2005). Large interindividual variability in the acute EPO response is consistently observed in younger healthy subjects exposed to moderate levels of hypoxia (≈2,500–3,000 m), with as much as a ≈400% difference noted between individuals (Chapman et al., 1998, 2010; Jedlickova et al., 2003; Friedmann et al., 2005; Gonzalez et al., 2006; Mounier et al., 2006). While this variability remains largely unexplained (Ge et al., 2002; Jedlickova et al., 2003; Gonzalez et al., 2006; Chapman et al., 2010; Hennis et al., 2010), several notable theories postulate that the hypoxic EPO response is dependent on oxidative and inflammatory signaling (Chandel et al., 1998; Morceau et al., 2009).
Notably, the antioxidant enzyme, extracellular superoxide dismutase (EC-SOD), has been identified as necessary for full EPO expression in response to hypoxia in mice (Suliman et al., 2004), while the proinflammatory cytokines interleukin-1alpha (IL-1α), IL-1 beta (IL-1β), and tumor necrosis factor-alpha (TNF-α) reportedly suppress hypoxia-induced erythropoiesis in vitro (Faquin et al., 1992; Fandrey et al., 1994; La Ferla et al., 2002; Krajewski et al., 2007). In human subjects, these relationships have been far less explored. Turner et al. (2017) found no relationship between the change in EPO in response to 2 hours of normobaric hypoxia simulating 3,600–4,800 m and IL-6 and TNF-α levels. However, proinflammatory cytokines notably increase after longer exposures (i.e., ≥12 hours) to hypoxia (Klausen et al., 1997; Hartmann et al., 2000; Mazzeo et al., 2001; Song et al., 2016; Lundeberg et al., 2018; Wang et al., 2018).
Zinc has a direct regulatory role in inflammation and oxidative stress (Jarosz et al., 2017). As a cofactor of the enzyme EC-SOD (Jarosz et al., 2017), it has been identified as a mediator of erythropoiesis in animal models (Chen et al., 2017, 2018). Evidence from human trials suggests that chronic zinc supplementation has the potential to influence the EPO response to hypoxia by reducing proinflammatory cytokines (Foster and Samman, 2012) and increasing EC-SOD activity (Kara et al., 2010). Therefore, the aim of our initial proof-of-concept investigation was to evaluate the influence of zinc supplementation on the acute erythropoietic and inflammatory responses of apparently healthy human participants exposed to a level of normobaric hypoxia (3,000 m; FiO2 = 0.14), typically used by athletes who train at altitude.
We hypothesized that 8 weeks of zinc supplementation (40 mg/day of zinc from picolinate) would increase serum zinc, reduce proinflammatory cytokine concentrations, and positively influence the erythropoietic response to hypoxia with minimal side effects associated with supplementation.
Methods
Participants
Ten physically active men (n = 6) and women (n = 4) volunteered for participation. Participants were recruited from the Indiana University Bloomington campus and were enrolled after providing written informed consent approved by the Indiana University Institutional Review Board. Individuals qualified if they reportedly engaged in ≥120 minutes of physical activity per week, had no history of renal disease, diabetes, and disorders related to hypoalbuminemia that would impair zinc metabolism (e.g., celiac disease, inflammatory bowel disease, cirrhosis, hepatitis).
All women were eumenorrheic, meaning they had consistent menstrual cycles lasting between 24 and 35 days in length for the duration of the study protocol. Women were tested during the same phase of their menstrual cycle for each experimental visit. A combined sample of men and women was selected given the evidence against sex-based differences in sea level EPO levels (Grote Beverborg et al., 2015) and the relative change in EPO with exposure to acute altitude (Chapman et al., 2010).
Participants were excluded if they were currently taking, or if they had taken any form of zinc supplement within 6 weeks before enrollment, were currently taking supplements that contained ≥10 mg of elemental iron, were born at an altitude ≥1,500 m, had previously lived at an altitude of ≥1,500 m for ≥7 days within 90 days before enrollment, or were planning to travel to an altitude of ≥1,500 m for ≥7 days during study enrollment.
Experimental design
Participants reported to the laboratory on six occasions after fasting for ≥8 hours. Three visits consisted of a continuous 12-hour stay in a normobaric hypoxic tent (FiO2 = 0.14) that simulated an altitude of 3,000 m. The level of hypoxia and duration of exposure were selected because of its relevance to current altitude training recommendations (i.e., 2,500–3,000 m, ≥12 h/day). The first hypoxic tent visit (PRE) was conducted before beginning zinc supplementation and a second after 8 weeks of supplementation (week 8). A third hypoxic tent exposure followed a 4-week washout period (WASH) during which zinc supplementation was discontinued. Three additional visits were completed at weeks 2, 4, and 6 of supplementation to assess serum zinc concentrations, adverse event frequencies, and compliance.
Upon arrival to the laboratory, participants had their height and mass measured along with measures of urine-specific gravity using a digital pen refractometer (Atago U.S.A., Inc., Bellevue, WA). A euhydration cutoff of 1.013 (Perrier et al., 2017) was set before blood collection. They were asked to return the following day if they did not meet the criteria for euhydration and were then excluded if they failed to meet the criteria for two consecutive days. Blood samples were obtained before hypoxia, after 6 and 12 hours at PRE, week 8, and WASH for subsequent analysis of serum zinc, EPO, proinflammatory cytokines, and EC-SOD. Additional blood samples were taken at weeks 2, 4, and 6 to monitor changes in serum zinc with supplementation.
Participants were required to complete three 24-hour dietary recall entries within 7 days of beginning supplementation and within 7 days of ending supplementation to assess dietary zinc intake using the electronic Automated Self-Administered 24 hours (ASA24®) Dietary Assessment Tool (v2018; National Cancer Institute, Bethesda, MD). Physical activity habits were self-reported via questionnaire at the same time points as the dietary recall.
Normobaric hypoxic tent exposures
Participants spent 12 hours resting quietly in a normobaric hypoxic tent (CAT-430™ Walk-In Tent; Colorado Altitude Training Systems, Denver, CO) on three separate occasions. The dimensions of the tent were 2.3 m × 2.9 m × 1.8 m, which comfortably fit two to three individuals per session. Participants were permitted to eat and drink ad libitum while in the tent.
For the hypoxic stimulus, portable nitrogen generators (Colorado Altitude Training Systems) were used to control the FiO2 of the enclosure at 0.14, simulating an altitude of 3,000 m. FiO2 and FiCO2 were continuously monitored using portable gas analyzers (Fast response O2 analyzer #17625 and CO2 analyzer #17630; VacuMed, Ventura, CA) in addition to temperature and humidity using a wireless weather station (Vantage Vue; Davis Instruments, Hayward, CA), which were recorded every 60 minutes. While positive pressure airflow of gas from the generators promoted turnover of air within the tent and minimized any rise in CO2 levels, a CO2 scrubber system (Sofnolime® 797 CO2 Absorbent; Molecular Products, Inc., Boulder, CO) was also used.
Peripheral oxyhemoglobin saturation (SpO2) was measured every 60 minutes using a fingertip pulse oximeter (Nellcor® DS100A-1; Covidien Ltd., Dublin, Ireland).
Zinc supplementation intervention
Participants completed 8 weeks of oral zinc supplementation during which they were asked to consume two doses of zinc as 20 mg tablets, equivalent to 40 mg/day of elemental zinc from picolinate (Douglas Labs, Pittsburgh, PA), once per day for 8 weeks. Instructions were provided to take the dose at the same time of day, preferably at least 1 hour before or 2 hours after a meal. Compliance with supplementation was assessed at each 2-week visit by asking participants to self-report the number of missed doses and by the researchers counting any remaining tablets. A side effect questionnaire was also administered at each 2-week visit during the 8-week supplementation period to assess the frequency of adverse events experienced with supplementation.
Dietary recall
Dietary recall entries were completed on 2 weekdays and 1 weekend day within 7 days of PRE and 7 days of week 8. Each participant received a unique username and password to remotely login to the ASA24 website. They then received reminders to complete three recalls within the requested time period and were excluded if they completed fewer than two entries within the 7-day allowance. From dietary recall analysis provided by ASA24 software, averages were obtained for the amount of zinc (mg) consumed per day to assess differences in dietary zinc intake with supplementation.
Blood sampling
Two 6 ml blood samples were collected via the antecubital vein from fasted participants in a silica clot-activated serum vacutainer (Becton Dickinson, Franklin Lakes, NJ) before entering the hypoxic tent and after 6 and 12 hours of exposure to hypoxia. One 6 ml sample was collected at 2-week intervals across the supplementation intervention (2, 4, 6 weeks) between hypoxic tent exposure visits. Serum was separated following a standardized clotting time of 30 minutes at room temperature by centrifugation at 1,300 g for 15 minutes at 4°C. Samples were then aliquoted into polypropylene cryogenic vials and stored at −80°C for <3 months until analysis.
Hemoglobin concentration
Capillary blood samples were collected at each experimental visit before participants entered the hypoxic tent and at 2-week intervals across the supplementation intervention (2, 4, 6 weeks) in a plastic microcuvette from the index finger in triplicate, and hemoglobin concentration was assessed using the HemoCue Hb200+ digital analyzer (Brea, CA) within 30 seconds of sampling. An average of the three samples was recorded.
Measurement of EPO, zinc, superoxide dismutase, C-reactive protein, and proinflammatory cytokines
Serum concentrations of EPO, zinc, EC-SOD, IL-1α, IL-1β, IL-6, TNF-α, and C-reactive protein (CRP) were determined via enzyme-linked immunosorbent assays in accordance with the manufacturer's instructions. Detection was performed with the use of a spectrophotometer (PowerWave® XS; BioTek Instruments, Winooski, VT). All samples were run in duplicate.
The assay kits had lower detection limits of EPO (ab119522; Abcam, Cambridge, United Kingdom) 0.4 mIU/ml, EC-SOD (Arbor Assays, Ann Arbor, MI) 0.044 U/ml, IL-1α (RayBiotech, Peachtree Corners, GA) 0.5 pg/ml, IL-1β (Thermo Fisher Scientific, Waltham, MA) 0.05 pg/ml, IL-6 (Thermo Fisher Scientific) 0.03 pg/ml, TNF-α (Boster Biological Technology, Pleasanton, CA) <1 pg/ml, and CRP (Cayman Chemical, Ann Arbor, MI) 0.05 pg/ml. The range of detection of the zinc assay (5-Br-PAPS Method; JaICA, Shizuouka, Japan) was 4–1,000 μg/dl. The intra-assay coefficient of variation was ≤12%.
Sample size considerations
An a priori power analysis using G*Power (v.3.1.9.4; Universitat Dusseldorf, Germany) projected that a sample size of nine participants would be requisite with the parameters 1-β = 0.8 and α = 0.05. The estimate was based on an anticipated increase in serum zinc of 19% with 8 weeks of supplementation (dz = 1.13) (Prasad et al., 2004) and an increase of 70% in EPO following 12 hours of hypoxia at 2,200 m (dz = 1.40) (Gonzalez et al., 2006).
Statistical analysis
All data were analyzed with SPSS (v.24; IBM Corp., Armonk, NY) and all figures were generated using Prism software (v.8; GraphPad Software, San Diego, CA). Normal distribution was assessed with the Shapiro–Wilk test and sphericity using Mauchly's test. A Greenhouse–Geisser correction was used in the event sphericity was violated. An analysis of covariance with repeated measures was conducted using average dietary zinc intake as a covariate to determine changes in serum zinc across the intervention. Mixed-effects modeling for repeated measures with a Bonferroni correction was used to determine differences between conditions for zinc, EPO, EC-SOD, CRP, and proinflammatory blood markers. Correlations were calculated using Pearson's product moment correlation coefficients. Effect sizes are reported as partial eta-squared (ηp2).
All data are reported as mean ± standard deviation and a p-value of <0.05 was considered statistically significant.
Results
Participant characteristics
A total of 10 participants completed PRE. One participant was withdrawn following the first week of supplementation due to side effects associated with Zn supplementation, and two electively dropped out due to time constraints that interfered with participation. Details of participant characteristics taken before and after the zinc supplementation protocol are presented in Table 1.
Participant Characteristics Pre- and Postsupplementation
Values are mean ± SD.
No differences were observed between PRE and week 8, p > 0.05.
n, number of subjects; PRE, presupplementation; week 8, after 8 weeks of zinc supplementation; SD, standard deviation.
Ambient conditions of the hypoxic exposure
The week 8 hypoxic tent exposure visit was conducted after 56 ± 2 days of Zn supplementation, and WASH following 29 ± 1 days after cessation of supplementation. The ambient O2 concentration of the enclosure remained constant (p = 0.297) between PRE, week 8, and WASH. As such, SpO2 was unchanged (p = 0.174) across visits and ranged from 89% to 97% between participants. There was no difference in ambient temperature (p = 0.187) that ranged from 21°C to 24°C between visits. However, differences in humidity were noted (p < 0.001) between PRE (50%) and week 8 (41%; p = 0.01) and week 8 and WASH (49%; p = 0.005).
Differences in serum zinc and compliance across the supplementation intervention
At the time of PRE, average serum zinc concentration was 114.9 ± 22.9; range: 89.4–149.3 μg/dl. Participants demonstrated 97% compliance in taking the total possible 56 doses of zinc picolinate. No participant reported missing more than two doses within any 2-week period throughout 8 weeks of supplementation. With supplementation, 40% of participants self-reported experiencing at least one adverse event (e.g., nausea, gastrointestinal discomfort, loss of appetite, headache, and metallic taste) attributed to the zinc supplement.
After controlling for differences in dietary zinc intake, supplementation had a significant effect on serum zinc concentrations (p = 0.005, ηp2 = 0.40, 1-β = 0.90). Compared with PRE, serum levels of zinc were significantly greater at week 2 (148.2 ± 64.0 μg/dl; p = 0.003), week 6 (139.1 ± 29.7 μg/dl; p = 0.047), and week 8 (135.2 ± 29.7 μg/dl; p = 0.043). At WASH, serum zinc concentrations returned to PRE (114.4 ± 12.3; p = 0.418). The relative change in serum zinc concentrations from PRE to supplementation is displayed in Figure 1.
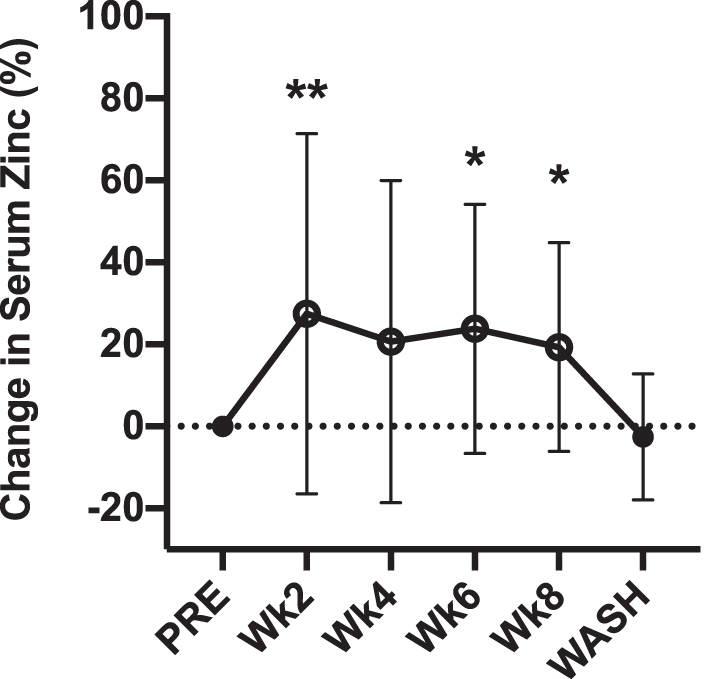
Percentage differences from presupplementation (PRE) in serum zinc concentration after weeks 2, 4, 6, 8 of supplementation with 40 mg/day of zinc from picolinate and following a 4-week washout. Open circles indicate time of supplementation. Values are mean ± SD. **p < 0.01. *p < 0.05. SD, standard deviation.
Differences in hemoglobin concentration across the supplementation intervention
Hemoglobin concentration at PRE was 14.5 ± 1.6 g/dl; range: 11.6–16.5 g/dl. The intervention had no effect on hemoglobin concentration (p = 0.143).
Differences in serum EPO with zinc supplementation and exposure to hypoxia
Serum EPO concentrations were unaltered by the zinc intervention (p = 0.932, ηp2 = 0.01, 1-β = 0.11). Time spent in hypoxia had a significant effect on EPO (p < 0.001, ηp2 = 0.60, 1-β = 1.0), with concentrations significantly increased from baseline (6.1 ± 0.6 mIU/ml) after 6 hours (9.1 ± 0.4 mIU/ml; p < 0.001) and 12 hours (10.6 ± 0.2 mIU/ml; p < 0.001). Figure 2 displays the mean erythropoietic responses to 12 hours of hypoxia at PRE, week 8, and WASH.

(
Differences in serum EC-SOD, CRP, and proinflammatory cytokines with supplementation and exposure to hypoxia
The zinc intervention had no effect on serum concentrations of IL-1α (p = 0.888), IL-1β (p = 0.171), IL-6 (p = 0.508), TNF-α (p = 0.940), or CRP (p = 0.580). There was additionally no difference in EC-SOD with supplementation (p = 0.615).
Time spent in hypoxia had no effect on IL-1α (p = 0.086), IL-1β (p = 0.579), TNF-α (p = 0.913), CRP (p = 0.912), or EC-SOD (p = 0.083). While serum concentrations of IL-6 did not change from 0 to 6 hours of hypoxia, levels were significantly elevated from baseline at 12 hours (p = 0.005, ηp2 = 0.25, 1-β = 0.81) due to an increase during the final 6 hours of exposure (p = 0.007). Figure 3 displays group mean data for EC-SOD, CRP, and proinflammatory cytokine measures taken at various time points of hypoxic exposure during the intervention.

Differences in serum concentration of
Relationships between the acute erythropoietic response to hypoxia, serum zinc, EC-SOD, CRP, and proinflammatory cytokines
The differences in EPO from baseline to 12 hours of hypoxia were not related to baseline concentrations of zinc, EC-SOD, CRP, or any proinflammatory cytokine (R2 ≤ 0.29, p ≥ 0.111) at PRE, week 8, or WASH. There was a significant relationship between EPO concentrations at 12 hours of hypoxia and baseline concentrations of IL-6 (R2 = 0.65, p = 0.008) and 12-hour concentrations of IL-6 (R2 = 0.78, p = 0.002). However, these relationships were not observed at week 8 (R2 ≤ 0.22, p ≥ 0.241).
Discussion
To our knowledge, this investigation is the first to assess the relationship between serum concentrations of EPO, zinc, and inflammatory cytokines with 12 hours of exposure to moderate hypoxia (FiO2 = 0.14; 3,000 m) in humans. Our working hypothesis anticipated that 8 weeks of zinc supplementation would augment the EPO response to hypoxia due to the positive influence of zinc on EC-SOD status (Kara et al., 2010) and its demonstrated potential to mediate erythropoiesis in animal models (Chen et al., 2017, 2018).
Although a significant increase in serum zinc concentration (19%) was observed following 8 weeks of supplementation, the EPO response to 12 hours of hypoxia remained unchanged with higher serum zinc levels. Furthermore, no relationships were observed between the acute EPO response and zinc, EC-SOD, CRP, or proinflammatory cytokines. Practically, our data do not support the application of zinc supplementation for the purpose of augmenting EPO in apparently healthy, younger adults acclimatizing to a moderate-level of hypoxia.
The influence of altered zinc bioavailability on erythropoiesis has been eloquently demonstrated in vitro (Lee et al., 2017) and in animal models (Konomi and Yokoi, 2005, 2018; Chen et al., 2017, 2018). A dose-dependent effect of zinc on erythropoiesis has been shown in rats (Chen et al., 2018) and low-oxygen dwelling fish (Chen et al., 2017). Furthermore, Konomi and Yokoi (2005) found moderate restrictions in dietary zinc intake to lower plasma EPO concentrations in rats presumably due to decreases in renal hypoxia-inducible factor gene expression (Konomi and Yokoi, 2018).
While mechanisms contributing to the synergistic effects of zinc bioavailability on EPO are unclear, preliminary evidence suggests involvement from TNF-α and IL-1β (Faquin et al., 1992; Fandrey et al., 1994; La Ferla et al., 2002; Krajewski et al., 2007) that is conceivably mediated by EC-SOD activity (Yu et al., 2008). Zinc supplementation reportedly suppresses gene expression of IL-1β and TNF-α (Prasad, 2014), which have been associated with blunted EPO transcription in response to hypoxia (Faquin et al., 1992; Fandrey et al., 1994; La Ferla et al., 2002; Krajewski et al., 2007). It is likely that activation of nuclear factor-kappaB by IL-1β and TNF-α partially mediates increases in EPO with exposure to hypoxia (Batmunkh et al., 2006), although the directional influence of this relationship is debated (Figueroa et al., 2002; La Ferla et al., 2002; Batmunkh et al., 2006).
The acute EPO and inflammatory response to 12 hours of hypoxia remained unchanged in our cohort of participants despite significant increases in serum zinc concentrations following supplementation with 40 mg/days of zinc. Furthermore, we observed no change in baseline levels of EC-SOD, CRP, or proinflammatory cytokines after 8 weeks of supplementation, which was unexpected. All participants had serum zinc levels within a normative range (>70 μg/dl) before supplementation (Trumbo et al., 2001). It is likely that the dose of zinc administered and/or magnitude of change in serum zinc, although statistically significant, may not have been enough to provoke changes in proinflammatory cytokines or EPO.
Higher doses of zinc (45–75 mg/day) have been associated with reductions in cytokine activity, namely IL-1 and TNF-α (Prasad et al., 2004, 2007; Bao et al., 2008, 2010; Foster and Samman, 2012). However, the potential risk of adverse health effects with doses higher than the one used in our study should be highlighted. Notably, 40% of our participants reported at least one side effect attributed to zinc supplementation. While most side effects were mild (e.g., metallic taste), one participant withdrew due to gastrointestinal discomfort that was described as severe enough to interfere with usual activities.
There are known side effects associated with zinc intakes exceeding the tolerable upper intake level of 40 mg/day, which include an impaired immune response and copper status (Trumbo et al., 2001). As a consequence of reduced copper bioavailability, EC-SOD activity has been shown to decrease with a total zinc intake of 60 mg/day for 10 weeks (Yadrick et al., 1989). When considering the additional intake of zinc from diet, the total daily zinc intake of our participants was 57 ± 7 mg (range: 47–68 mg), exceeding the tolerable upper intake limit. Thus, while there is a chance that higher doses of zinc may effectively reduce proinflammatory cytokine activity, a total dietary zinc intake exceeding the tolerable upper intake limit is unadvisable and therefore impractical for individuals traveling to altitude.
Although zinc supplementation did not influence proinflammatory cytokine activity with acute hypoxia, our data elucidate the cytokine response to levels of hypoxia equivalent to 3,000–4,800 m (Klausen et al., 1997; Hartmann et al., 2000; Mazzeo et al., 2001; Song et al., 2016; Turner et al., 2017; Wang et al., 2018). Turner et al. (2017) found that circulating IL-6 concentrations were unchanged at 2 hours of exposure to 3,600–4,800 m of normobaric hypoxia, while Julian et al. (2011) observed a positive trend for IL-6 (p < 0.10) at 9 hours of exposure to 4,875 m. Our data support these findings, as serum IL-6 concentrations increased at 12 hours but not at 6 hours of exposure. A delayed IL-6 response to hypoxia is supported by others who have reported increases at 24–96 hours (Klausen et al., 1997; Hartmann et al., 2000; Mazzeo et al., 2001; Song et al., 2016; Wang et al., 2018). Conversely, TNF-α, IL-1α, IL-1β, and CRP remained unchanged in our investigation.
Although speculative, it is possible that rising concentrations of IL-6 inhibited the release of TNF-α and IL-1. IL-6 exerts both pro- and anti-inflammatory effects, and administration via injection/infusion has been shown to inhibit TNF-α production in a dose-dependent manner (Mizuhara et al., 1994) and stimulate the release of the anti-inflammatory IL-1 receptor antagonist and IL-10 (Steensberg et al., 2003). The time-courses of IL-6 and IL-1 receptor antagonist release with exposure to 3,458 m also appear to coincide (Hartmann et al., 2000). An alternative explanation may be that our exposure time of 12 hours was too truncated to observe increases in other proinflammatory cytokines and acute-phase proteins. For example, others have shown TNF-α and IL-1β to increase at 72 hours of exposure to 3,200–3,860 m (Song et al., 2016; Wang et al., 2018). The peak in IL-6 at 30 hours also appears to precede the peak in CRP at 66 hours of exposure to 3,458 m (Hartmann et al., 2000).
Unlike IL-1α, IL-1β, and TNF-α (Faquin et al., 1992; Fandrey et al., 1994), IL-6 activity has been positively associated with EPO expression (Faquin et al., 1992; Fandrey et al., 1994; Klausen et al., 1997). In our participants, a positive association was observed between serum IL-6 concentrations at baseline and EPO concentrations following 12 hours of hypoxia. In vitro experiments have demonstrated dose-dependent stimulation of hypoxia-induced EPO with IL-6 (Faquin et al., 1992; Fandrey et al., 1994). In addition, a positive association has also been observed between IL-6 and EPO concentrations in human subjects after 96 hours of exposure to 4,350 m (Klausen et al., 1997). However, these findings should be interpreted with caution as IL-6 is known to upregulate hepcidin synthesis (Camaschella et al., 2020) and may therefore impair erythroid cell development with acclimatization to hypoxia.
The incorporation of antioxidant-rich foods is advocated by Stellingwerff et al. (2019) as a nutritional strategy to counteract oxidative stress with chronic altitude training practices. Specifically, in relation to the importance of zinc supplementation for altitude exposure, hypoxic stress prompts a redistribution of circulating zinc from plasma into organs and tissues (Rawal et al., 1999). Zinc supplementation (15 mg/day) has been shown to prevent against a ≈25% reduction in plasma zinc concentrations in response to 3,500 m of altitude observed in an unsupplemented control group (Rawal et al., 1999).
Despite the potential for high-dose antioxidant supplements to mitigate the effects of oxidative stress in individuals traveling to altitude, there have been concerns regarding whether such supplements may negatively impact adaptive hematological responses to altitude training (Stellingwerff et al., 2019). Notably, an increase in the total intake of antioxidant-rich foods did not influence changes in hemoglobin mass or maximal aerobic capacity following 3 weeks of altitude training in national team endurance athletes (Koivisto et al., 2018). Our data lend further support to the notion that antioxidant supplements (zinc) do not necessarily impair the acute EPO response to moderate altitude. However, these data should be interpreted with caution as further studies are warranted to comprehensively evaluate the downstream effects of zinc supplementation on changes in hemoglobin mass and aerobic performance following chronic exposure to altitude.
In addition to confirming the considerable interindividual variability in the acute EPO response to hypoxia (range: −1% to +208%) that is similar to others who have observed as much as a ≈400% difference between individuals (Chapman et al., 1998, 2010; Jedlickova et al., 2003; Friedmann et al., 2005; Gonzalez et al., 2006; Mounier et al., 2006), our data demonstrate a considerable degree of intraindividual variability in the response. Within the same individual, the relative change in EPO from baseline to 12 hours of hypoxia differed by an average of 49 ± 31% (range: 4%–90%) between consecutive exposures. A case study conducted by Garvican et al. (2007) assessed differences in the acute EPO response to repeated exposures of hypoxia in a male professional cyclist, finding a 49% difference (range: −11% to +38%) in the EPO response to four 8-hour exposures of normobaric hypoxia simulating 2,800 m.
Practically, our data and Garvican et al. (2007) demonstrate inconsistency in the individual EPO response to simulated altitude. Currently, there is no method for identifying individuals who will respond favorably (i.e., greater increases in hemoglobin mass, improved physical performance) to an altitude training intervention. When considering the substantial amount of time and resources that are allocated by athletes to overcome the strategic burdens associated with altitude training practices, nutritional interventions aimed at maximizing hypoxic-driven erythropoiesis are warranted. Additional research is needed to fully elucidate the mechanisms contributing to the unexplained variability in the acute EPO response to hypoxia.
Limitations and future directions
The aim of our investigation was to establish proof-of-concept for zinc supplementation as a strategy to augment the EPO response to a level of hypoxia typically used by athletes who train at altitude. While our sample size was relatively small, our study was sufficiently powered to detect changes in serum concentrations of EPO (1-β = 1.0) and IL-6 (1-β = 0.81) at 12 hours of exposure to hypoxia and in serum zinc concentrations following 8 weeks of supplementation (1-β = 0.90). Furthermore, a post hoc power analysis demonstrated that a sample size of >300 participants would be necessary to detect changes in the EPO response at 12 hours with zinc supplementation.
While our findings do not support chronic supplementation with 40 mg/day of zinc from picolinate as a practical approach to augment EPO in response to hypoxia, it is possible that a higher dose of zinc or alternate route of administration (e.g., intravenous infusion) may elicit positive effects on EPO in apparently healthy zinc-sufficient humans. However, the feasibility of such approaches should be evaluated with respect to the greater likelihood and severity of anticipated side effects with higher zinc doses and the practical considerations for individuals sojourning to altitude.
In perspective of the positive association that was found between IL-6 concentrations and EPO, supported by other accounts (Faquin et al., 1992; Fandrey et al., 1994; Klausen et al., 1997), and the negative influence of IL-6 on erythroid cell development via upregulated hepcidin, future studies of longer duration are needed to assess the effects of zinc supplementation on hematological parameters with altitude acclimatization. We recommend such studies include biomarkers of iron metabolism, including serum iron, ferritin, and soluble transferrin receptor concentrations. In addition, given the evidence for increased oxidative stress with hypobaric compared with normobaric hypoxia simulating 3,000 m (Faiss et al., 2013), a field study in which subjects sojourn to terrestrial altitude with and without zinc supplementation may yield different results from our present findings.
Conclusions
Our data indicate that the acute EPO and proinflammatory responses of apparently healthy zinc-sufficient individuals traveling to 3,000 m of altitude are unaffected by 8 weeks of supplementation with 40 mg/day of zinc from picolinate. The considerable interindividual variability observed in the EPO response to 12 hours of exposure to hypoxia was not explained by differences in plasma zinc, EC-SOD, CRP, or proinflammatory cytokine concentrations.
Footnotes
Authors' Contributions
M.N.B., J.P., A.D.F., B.J.M., T.D.M., and R.F.C. conceived the experimental design. M.N.B. and J.P. carried out the experiment. M.N.B. analyzed the results with R.F.C. and H.L.P. contributing to the interpretation of results. M.N.B. wrote the original article with revisions from J.P., A.D.F., B.J.M., T.D.M., H.L.P., and R.F.C. All authors have reviewed and approved the final version of the article before submission.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this work (supported by internal/faculty research accounts).