
Abstract
Cai, Jing, Junyong Ruan, Xi Shao, Yuanjun Ding, Kangning Xie, Chi Tang, Zedong Yan, Erping Luo, and Da Jing. Oxygen enrichment mitigates high-altitude hypoxia-induced hippocampal neurodegeneration and memory dysfunction associated with attenuated tau phosphorylation. High Alt Med Biol. 22:274–284, 2021.
Background:
Brain is predominantly vulnerable to high-altitude hypoxia (HAH), resulting in neurodegeneration and cognitive impairment. The technology of oxygen enrichment has proven effective to decrease the heart rate and improve the arterial oxygen saturation by reducing the equivalent altitude. However, the efficacy of oxygen enrichment on HAH-induced cognitive impairments remains controversial based on the results of neuropsychological tests, and its role in HAH-induced hippocampal morphological and molecular changes remains unknown. Therefore, this study aims to systematically investigate the effects of oxygen enrichment on the memory dysfunction and hippocampal neurodegeneration caused by HAH.
Materials and Methods:
Fifty-one male Sprague-Dawley rats were equally assigned to three groups: normal control, HAH, and HAH with oxygen enrichment (HAHO). Rats in the HAH and HAHO groups were exposed to hypoxia for 3 days in a hypobaric hypoxia chamber at a simulated altitude of 6,000 m. Rats in the HAHO group were supplemented with oxygen-enriched air, with 12 hours/day in the hypobaric hypoxia chamber.
Results:
Our results showed that oxygen enrichment improved the locomotor activity of HAH-exposed rats. The Morris water maze test revealed that oxygen enrichment significantly ameliorated HAH-induced spatial memory deficits. Oxygen enrichment also improved morphological alterations of pyramidal cells and the ultrastructure of neurons in the hippocampal CA1 region in rats exposed to acute HAH. Tau hyperphosphorylation at Ser396, Ser262, Thr231, and Thr181 was also significantly attenuated by oxygen enrichment in HAH-exposed rats.
Conclusions:
Together, our study reveals that oxygen enrichment can ameliorate HAH-induced cognitive impairments associated with improved hippocampal morphology and molecular expression, and highlights that oxygen enrichment may become a promising alternative treatment against neurodegeneration for humans ascending to the plateau.
Introduction
Hypobaric hypoxia, a unique environmental condition at high altitudes, is associated with the decline in atmospheric pressure and consequently reduced partial pressure of oxygen (West, 2012). Millions of individuals (e.g., soldiers, miners, and travelers) ascend to the plateau for occupational or recreational pursuits with the advent of more convenient transportation to high altitudes (Moore, 2001; West, 2015). Migrants ascending to high altitudes exceeding 2,500 m suffer from acute mountain sickness with symptoms such as headache, insomnia, and fatigue (Hackett and Roach, 2001).
The brain requires up to 20% of the total oxygen consumption (Raichle and Gusnard, 2002) and thus is especially vulnerable to high-altitude hypoxia (HAH) conditions (Lieberman et al., 1994). Several field-based studies have shown that HAH jeopardizes human brain functions and causes progressive neuronal injuries even after people return to the sea level (Bahrke and Shukitt-Hale, 1993; Cavaletti and Tredici, 1993). Substantial studies have demonstrated that humans exhibit extended reaction time, shortened attention span, and impaired learning and spatial memory after acute exposure to HAH (Cavaletti and Tredici, 1993; Virués-Ortega et al., 2004; McMorris et al., 2017). Considering that HAH has profound detrimental impacts on the brain structure and cognitive performance of high-altitude populations, it is necessary to explore effective countermeasures to attenuate HAH-induced cognitive impairments.
Increasing the intake of oxygen using equipment such as bottled oxygen, hyperbaric oxygen chamber, and portable hyperbaric chamber is a common strategy to ameliorate HAH-induced deleterious effects. Bottled oxygen can provide high-concentration oxygen to ameliorate symptoms of high-altitude sickness (Peacock, 1998), whereas this technique is difficult to be widely applied at high altitudes due to the limited oxygen supply and potential oxygen toxicity (Deneke and Fanburg, 1980). Although hyperbaric oxygen chamber has proven to be effective to mitigate HAH with increased partial pressure of oxygen in arterial blood (Kasic et al., 1991; Zhu et al., 2015), the poor portability and possible side effects (such as middle ear injuries, pulmonary barotrauma, and hyperoxic myopia) limit its extensive application at high altitudes (Heyboer et al., 2017). It has been demonstrated that the portable hyperbaric chamber exerts beneficial effects on HAH-related sickness (Bärtsch, 1992). However, some patients may suffer from vomiting or claustrophobia due to the confined space of the portable hyperbaric chamber (Davis and Hackett, 2017).
Oxygen enrichment is increasingly recognized as an effective approach to reduce the equivalent altitude, which can raise the oxygen concentration to mimic higher oxygen partial pressure of a lower altitude (West, 2003, 2012). Several studies have demonstrated that every 1% increase in oxygen concentration results in an ∼300 m decrease in equivalent altitude (Luks et al., 1998; West, 2002, 2016). The technology of oxygen enrichment is able to offer long-run supply of oxygen-enriched air with rare side effects (Nagatomo et al., 2012). By utilizing the membrane separation technique, our team has developed novel oxygen enrichment equipment for various situations at high altitudes (e.g., the manpack version, vehicle-mounted version, and room-mounted version), which has proven effective to decrease the heart rate and improve the saturation of peripheral oxygen (Shen et al., 2013).
Several attempts have been made by investigators to study the effects of the oxygen enrichment technique on the cognitive impairments for humans at high altitudes, whereas the potential efficacy remains controversial (Luks et al., 1998; Gerard et al., 2000; Moraga et al., 2018). Moreover, these clinical studies only performed neuropsychological tests without systemic assessment of the potential changes of brain morphology and function (Gerard et al., 2000; McMorris et al., 2017; Moraga et al., 2018).
It is well known that the hippocampus is an important region of the brain, which plays a critical role in the memory and spatial navigation (Lisman et al., 2017; Ólafsdóttir et al., 2018; Rolls and Wirth, 2018). A growing body of evidence has shown that pyramidal neurons in the CA1 region of the hippocampus are highly sensitive to the deleterious effects of hypoxia (Kreisman et al., 2000; Gozal et al., 2002), and the hippocampal CA1 region is regarded as a potential important target for resisting HAH-induced cognitive impairments (Wang et al., 2002). However, the effects of oxygen enrichment on HAH-induced morphological and molecular changes in the hippocampus (especially in the CA1 region) and the corresponding neurological functions have never been investigated.
In this study, we aim to investigate the efficacy of oxygen enrichment on the hippocampal neurodegeneration at the tissue, cellular, and molecular levels. An acute HAH animal model established by exposure to the simulated hypoxia conditions at high altitude (6,000 m) was subjected to the treatment with oxygen enrichment. Locomotor activity and spatial working memory of rats were evaluated via the open field test and Morris water maze test, respectively. The morphology, ultrastructure, and expression of Tau proteins of neurons in the hippocampal CA1 region were examined via histological, transmission electron microscopy, and Western blotting assays. This study represents the first effort investigating the potential effects of oxygen enrichment on the neurodegenerative alterations in the hippocampal CA1 region and hippocampus-dependent memory functions in HAH animals.
Methods
Animals and experimental design
Fifty-one male Sprague-Dawley rats (7–8 weeks old; 192–230 g) were purchased from the Animal Center of the Fourth Military Medical University. All rats were housed under controlled temperature (23°C ± 1°C) and relative humidity (40%–60%) with a 12-hour light/dark cycle. The animals were allowed to get free access to tap water and food and acclimate to the environment for 1 week before the start of experiments. All animal protocols were approved by the Institutional Animal Care and Use Committee of the Fourth Military Medical University, and all animal procedures were strictly performed according to the guidelines published by the National Institutes of Health.
All rats were randomly divided into the following three groups (17 rats in each group): normal control (NC) group, HAH group, and HAH with oxygen enrichment (HAHO) group. Rats in the NC group were kept under normobaric normoxia in the city of Xi'an (400 m above sea level). Rats in the HAH group and the HAHO group were housed in the hypobaric hypoxia chamber equivalent to 6,000 m above sea level for 3 continuous days. Rats in the HAHO group were subjected to oxygen enrichment treatment for 12 hours per day using the novel oxygen enrichment device developed by our group. Oxygen-enriched air was provided from 8 a.m. to 8 p.m. (opposite circadian rhythm between human and rat) to simulate an oxygen enrichment treatment for highlanders during the rest and sleep time without influencing their daytime work.
A total of 21 rats (n = 7 in each group) were used to assess the locomotor activity and the morphology of neurons, and the other 30 rats (n = 10 in each group) were assessed for spatial working memory, the ultrastructure of neurons, and the expression of Tau protein in the hippocampal CA1 region, respectively.
Hypobaric hypoxia exposure and local oxygen enrichment
A commercial hypobaric hypoxia chamber (#LAT-SY01; Suzhou Liante Medical Equipment Co., Ltd, Suzhou, China) was utilized to simulate the hypobaric hypoxic conditions equivalent to an altitude of 6,000 m (barometric pressure, 47.2 kPa; partial oxygen pressure, 9.9 kPa). The air in the chamber was evacuated through vacuum pumps and the proportion of gas into or out of the chamber was precisely modulated by solenoid valves to allow air circulation. The chamber was equipped with a pointer pressure gauge and a control panel to monitor and maintain the barometric pressure in the chamber. In addition, high-precision sensors were used to monitor the real-time temperature and humidity in the chamber. The ascending and descending rates of the chamber were both set to 91.3 Pa/s.
A local oxygen-enriched environment was constructed in the hypobaric hypoxia chamber by assembling the portable oxygen enrichment device to an individually ventilated cage (IVC) for rats (Fig. 1A). Based on the membrane gas separation technique, the portable and convenient oxygen enrichment device was manufactured by our group (China Patent No. ZL201210309573.3 and ZL201310468909.5) and has been described precisely in our previous study (Shen et al., 2013).

Experimental setup and experimental protocol used in the current study.
In brief, the device mainly consisted of five parts: a vacuum pump, a DC ventilation fan, a filter unit, 16 oxygen enrichment membrane units, and a gas buffer. The vacuum pump was used to evacuate the internal air out to form a negative pressure environment and thus drive the external air passing through the oxygen-enriched membrane. A vacuum controller was used to accurately control the baric pressure gradient across the membrane. Each oxygen-enriched membrane unit contained two layers of membrane supported by a rectangular frame. Considering a higher permeation rate of oxygen than nitrogen, the oxygen-enriched membrane was coated with a selective layer of polymer solution to reach high oxygen permeability and consequently separate the gas. The gas buffer was utilized to stabilize air flow and reduce noise.
The oxygen concentration in the IVC was determined using an oxygen analyzer (OXYMAT 61; Siemens, Erlangen, Germany) under normobaric normoxic and simulated hypoxic conditions, respectively. As shown in Figure 1B, the 12-hour oxygen concentration curve (without oxygen enrichment) in the hypobaric chamber at a simulated altitude of 6,000 m was similar to that under normobaric normoxic conditions. The oxygen enrichment device has the ability to increase the oxygen concentration in the IVC to 30% within 20 minutes under normobaric normoxia, and maintained the oxygen concentration at 32.5% ± 1.1% (Fig. 1C).
The oxygen concentration of the oxygen-enriched air in the IVC was maintained in the range of 28.4%–30.5% at the simulated altitude from 2,500 to 6,000 m, and the temperature was maintained in the range of 21.6–22.7°C (Fig. 1D). Partial oxygen pressure of the air in the hypobaric chamber at the simulated altitude of 6,000 m with and without oxygen enrichment was 15.34 and 11.51 kPa, respectively.
Open field test
An activity recording device (7420; Ugo-Basile, Varese, Italy) equipped with a transparent plexiglass cage (40 × 40 × 31 cm) was utilized to evaluate the spontaneous locomotor activity of rats in the three groups. This device contained an electronic unit incorporating a counter and two vertical facing blocks generating/perceiving infrared rays. Infrared beam interruptions caused by the movements of animals in vertical and horizontal directions were automatically recorded. Before testing, rats were placed into the center of the device and allowed to move freely. After an initial 10-minute habituation in the device, the numbers of locomotor activity of the animals in the vertical and horizontal directions were collected for a period of 10 minutes.
Morris water maze test
The spatial working memory of rats in the three groups was assessed using the Morris water maze test. The water maze contained a polypropylene circular pool with 120 cm in diameter and 50 cm in depth. The pool was separated into four identical quadrants: northwest, northeast, southwest, and southeast. The pool was filled with water (maintained at 24°C) to a depth of 35 cm, and nontoxic black ink was added to make the water opaque. The escape platform (20 cm in diameter) was submerged into the center of northwest quadrant at 1 cm beneath the water surface. The pool was surrounded by numerous visible cues for rats to identify the location of the hidden submerged platform. An infrared digital camera was installed above the pool to record the swimming paths of rats.
The whole tests included the acquisition trials for 5 consecutive days and probe trial on the ninth day after the HAH exposure (Fig. 1E). In the acquisition trials, rats were trained for four times per day with an interval of 1 minute. During the trial, rats were released into the water choosing any of the four quadrants as the starting position, and trained to locate the platform for 60 seconds. If a rat failed to reach the platform within 60 seconds, it would be gently guided to the platform for adaptation and memory for 10 seconds. After the training, rats were dried with a towel and returned to the home cages. The swimming track and the escape latency (time to reach the hidden platform) were recorded.
After the acquisition trials, rats were exposed to HAH/HAHO for 3 days and the probe trial was then performed. During the probe trial, the hidden platform was retrieved from the pool, and rats were allowed to swim freely for 120 seconds. The frequency of platform crossings and the path traveled in the target quadrant were recorded for the assessment of spatial working memory.
Histological analysis
Rats were immediately anesthetized intraperitoneally using 3% sodium pentobarbital and transcardially perfused with normal saline through the ascending aorta. The skull and cranial nerves were rapidly removed, and the CA1 regions of hippocampus were dissected out carefully and fixed in 4% paraformaldehyde for 24 hours. Then, the brain samples were dehydrated in a graded series of ethanol, transparentized in xylene, and embedded in paraffin. Five-micrometer-thick sections were cut and then stained with hematoxylin and eosin (H&E). Samples were observed using an optical microscope (Olympus, Tokyo, Japan) for hippocampal morphology. Five fields at 400 × magnification in each section were randomly selected and quantified for the number of denatured pyramidal cells.
Transmission electron microscopy
After sacrifice, specimens from the hippocampal CA1 region were disserted and cut into tissue blocks with ∼1 mm3, and then immediately fixed in 2% paraformaldehyde and 3% glutaraldehyde at 4°C for 4 hours. The specimens were rinsed in 0.1 M phosphate-buffered saline three times and fixed in 1% osmium tetroxide at room temperature for 2 hours. Then, samples were dehydrated with a series of graded ethanol solutions, embedded in Embed-812 epoxy resin, and cut into 60–80 nm sections using Leica EM UC7 ultramicrotome (Leica Microsystems, Wetzlar, Germany). The sections were stained with uranyl acetate and lead citrate, and the ultrastructure of neurons was examined using a transmission electron microscope (HT7700; Hitachi, Tokyo, Japan).
Western blotting
The CA1 region of the hippocampus of rats was isolated and frozen at −80°C for Western blotting analysis. Samples were homogenized with Radio Immuno Precipitation Assay (RIPA) lysis buffer containing 1% enylmethylsulfonyl fluoride (PMSF) to extract the total protein. The homogenates were centrifuged at 12,000 rpm for 10 minutes at 4°C, and the protein concentration of the supernatant was quantified using the BCA assay kit (Thermo Fisher Scientific, Waltham, MA). The proteins were electrophoresed on 10% sodium dodecyl sulfate (SDS)-polyacrylamide gels and transferred to polyvinylidene difluoride (PVDF) membranes (Millipore Corporation, Bedford, MA).
The membranes were then blocked in 5% skim milk for 1 hour at room temperature, and then incubated with the rabbit anti-rat antibody Tau-5 and phospho-specific anti-tau antibodies Ser396, Ser262, Thr231, and Thr181 (Abcam, Cambridge, United Kingdom) overnight at 4°C. Then, the membranes were washed with TBST buffer (tris-buffered saline, 0.1% Tween 20) thrice for 10 minutes, and then incubated with the horseradish peroxidase-labeled goat anti-rabbit IgG secondary antibody (Abcam) for 1 hour. After washing three times with TBST for 10 minutes, the blots were visualized by an ECL chemiluminescent system (GE ImageQuant 350; GE Healthcare).
Statistical analysis
The SPSS 20.0 software (SPSS, Inc., Chicago, IL) was used to perform the statistical analysis. Normal distribution was examined using the Kolmogorov–Smirnov test, and homogeneity of variance was determined using the Levene's test. All data were proven to obey normal distribution and homoscedasticity. One-way analysis of variance with Bonferroni's post hoc test was used to compare the differences between every two groups. All data are presented as mean ± SD. p < 0.05 was considered to be statistically significant.
Results
Effect of oxygen enrichment on body mass and locomotor activity in HAH rats
As shown in Figure 2A, no significant difference in the body weight among the three groups was observed before oxygen enrichment treatment (p > 0.05). However, after a 3-day hypobaric hypoxia exposure, the body weight of rats in the HAH group (p < 0.001, −12.1%) was significantly lower than that in the NC group. The HAHO group exhibited a significantly higher body weight than the HAH group (p < 0.05, +6.9%) and a significantly lower body weight than the NC group (p < 0.05, −6.0%).
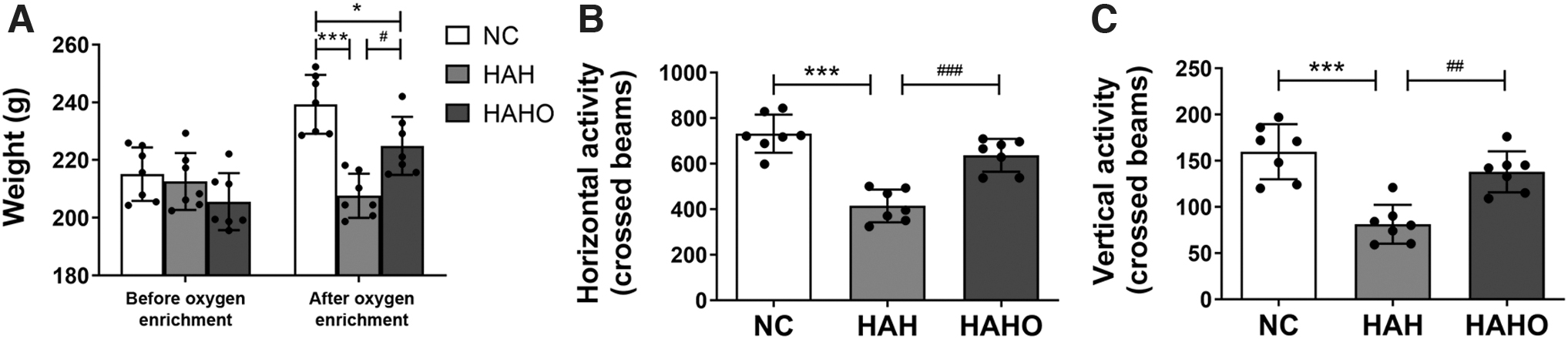
Effect of oxygen enrichment on body mass and locomotor activity in rats exposed to 3-day hypobaric hypoxia.
Rats in the HAH group showed a significant decrease in the horizontal activity (Fig. 2B, p < 0.001, −43.4%) and vertical activity (Fig. 2C, p < 0.001, −49.2%) compared with the NC group. In addition, the HAHO group exhibited a significantly increased horizontal activity (p < 0.001, +53.6%) and vertical activity (p < 0.01, +70.0%) compared with the HAH group. No significant difference in the horizontal or vertical activity was found between the HAHO group and the NC group (p > 0.05).
Effect of oxygen enrichment on spatial working memory in HAH rats
In the acquisition trials, a trend of decrease in the escape latency during the five successive training days was found in all the three groups (Fig. 3A), revealing that rats acquired spatial memory of the location of the hidden platform. Nevertheless, no significant difference was observed in the escape latency of rats among the three groups at each day (p > 0.05). As shown in Figure 3B and C, the numbers of platform crossings (p < 0.001, −62.2%) and target quadrant entries in the HAH group were significantly lower than those in the NC group. The HAHO group showed a significantly increased frequency that rats crossed the platform area (p < 0.001, +143.1%) and an increased number of target quadrant entries compared with the HAH group. No significant difference in the number of platform crossings was observed between the NC group and the HAHO group (p > 0.05).

Effect of oxygen enrichment on spatial working memory in rats exposed to 3-day hypobaric hypoxia.
Effect of oxygen enrichment on the morphology in the hippocampal CA1 region in HAH rats
According to the H&E staining results, pyramidal neurons in the hippocampal CA1 region were round shaped with clearly visible nucleus and intact membranes in the NC group (Fig. 4A). After hypobaric hypoxia exposure, neurons in the HAH group were pyknotic and irregularly shaped. The boundaries between the nucleus and cytoplasm were invisible, and the distribution of neurons was disorganized in the HAH group (Fig. 4B). Oxygen enrichment significantly mitigated the morphological changes of neurons, as evidenced by decreased nuclear pyknosis and improved structure of neurons in the HAHO group (Fig. 4C).
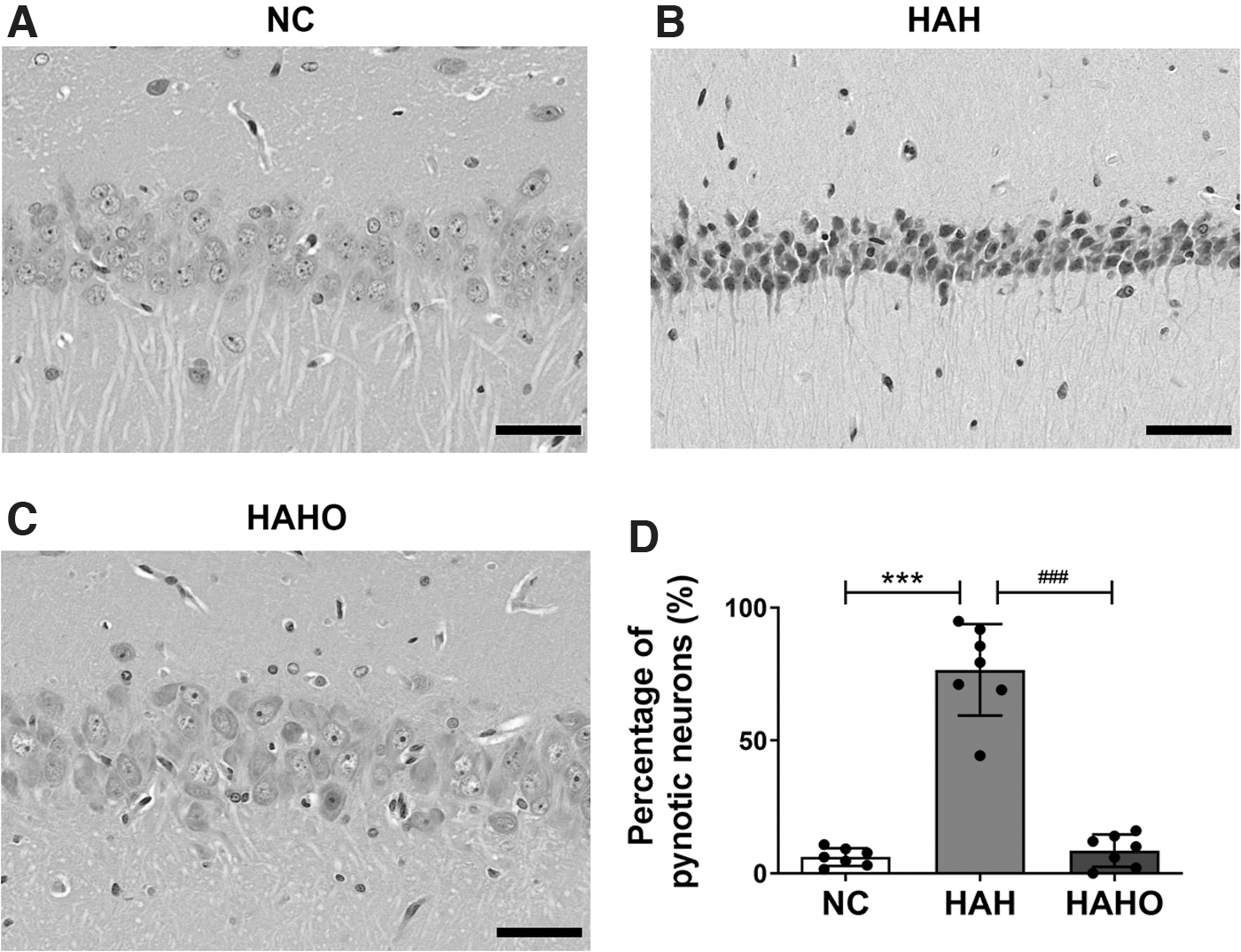
Effect of oxygen enrichment on morphology in the hippocampal CA1 region in rats exposed to 3-day hypobaric hypoxia.
The HAH group exhibited a significantly higher percentage of pyknotic neurons than that in the NC group (Fig. 4D, p < 0.001, +1144.4%). The percentage of pyknotic neurons was significantly decreased in the HAHO group compared with the HAH group (p < 0.001, −88.8%), whereas no significant difference in the percentage of pyknotic neurons was observed between the HAHO and the NC groups (p > 0.05).
Effect of oxygen enrichment on the ultrastructure of hippocampal neurons in HAH rats
The ultrastructure of neurons in the hippocampal CA1 area assessed by transmission electron microscopy is shown in Figure 5. The NC group exhibited an intact membrane, large and oval nuclei containing fine and uniform chromatin, and undamaged mitochondria with well-structured cristae. Nevertheless, acute hypobaric hypoxia exposure has detrimental effects on the ultrastructure of hippocampal neurons, as evidenced by a disrupted membrane, increased chromatin condensation, and swollen mitochondria with few cristae in the HAH group. The HAHO group showed an improved membrane, remarkably reduced chromatin condensation, and neatly arranged mitochondria cristae compared with the HAH group.

Effect of oxygen enrichment on the ultrastructure of hippocampal neurons in rats exposed to 3-day hypobaric hypoxia.
Effect of oxygen enrichment on the expression of Tau proteins in the hippocampus in HAH rats
The results of Western blotting revealed that no significant change in the expression of total Tau protein was found among the three groups (Fig. 6A, B; p > 0.05), suggesting that acute HAH exposure exerted little effect on the total protein expression of Tau (including phosphorylated and nonphosphorylated Tau protein) in rats.

Effect of oxygen enrichment on the expression of Tau proteins in the hippocampus in rats exposed to 3-day hypobaric hypoxia.
Tau phosphorylation at four specific sites (Ser262, Thr181, Ser396, and Thr231) is shown in Figure 6C
Discussion
Previous studies by our and other groups suggest that oxygen enrichment, as a safe and convenient technique, is able to increase the arterial oxygen saturation and decrease the heart rate for individuals on the plateau (Shen et al., 2013; Li and Liu 2014; Moraga et al., 2014). The potential effects of oxygen enrichment on HAH-induced cognitive impairments remain controversial based on the clinical results of neuropsychological tests (Luks et al., 1998; Gerard et al., 2000; Moraga et al., 2018), and its impacts on HAH-induced hippocampal morphological and molecular changes are still unknown.
This study is the first to investigate the efficacy of oxygen enrichment on the hippocampal neurodegeneration caused by HAH at the tissue, cellular, and molecular levels. Our results showed that oxygen enrichment has the capacity of mitigating the morphological changes of pyramidal cells, improving the ultrastructure of neurons, and suppressing the hyperphosphorylation of Tau proteins in the CA1 region of the hippocampus.
Several previous studies have demonstrated that HAH leads to reduction in the body weights of highlanders, which is possibly associated with the reduced energy intake (e.g., impaired intestinal function and decreased appetite) and increased basal metabolic rate (Lippl et al., 2010; Dünnwald et al., 2019). In the current study, we found that oxygen enrichment attenuated the reduction in body weights for rats exposed to HAH, whereas the body weights were not able to completely restore to the control level.
The open field test is a powerful paradigm for evaluating the spontaneous exploration and locomotor activity of laboratory rodents by assessing the infrared beam interruptions induced by the animal motions (Walsh and Cummins, 1976; Crawley, 1999). It has been reported that hypoxia inhibits the locomotor activity of animals via the open field test (Car et al., 2001; Braga et al., 2013). Similarly, our behavioral data showed a significant decline in the number of horizontal activity and vertical activity in HAH-exposed rats. Moreover, our open field test results showed that oxygen enrichment significantly increased the horizontal and vertical activities for rats exposed to HAH, which approximates to the control level, revealing the significantly improved exploratory and locomotor activities following oxygen enrichment.
A number of studies have shown that hypoxia has detrimental effects on the morphology and function of the brain, and even transient hypoxia can result in permanent neuronal injury (Furling et al., 2000; Li et al., 2013; Hou et al., 2019). It is well recognized that the hippocampus plays an essential role in memory and mental construction (Sheldon and Levine, 2016; Lisman et al., 2017; Ólafsdóttir et al., 2018; Rolls and Wirth, 2018), and is predominantly susceptible to the damage induced by hypoxia (Hota et al., 2007). Pyramidal neurons in the hippocampal CA1 region are especially vulnerable to HAH, leading to hippocampal neurodegeneration (Kreisman et al., 2000; Ghotbeddin et al., 2020). Growing evidence implicates that hypoxia imbalances mitochondrial fusion and fission, adversely changes mitochondrial morphology (Jain et al., 2015), and disrupts mitochondrial oxidative phosphorylation (Turrens, 2003). As a consequence, oxidative stress occurs and induces neuronal cell death and the subsequent pathogenesis of neurodegenerative disorders (Coimbra-Costa et al., 2017).
Our results, consistent with previous studies (Baitharu et al., 2012; Zhu et al., 2019), showed that pyramidal cells became pyknotic, small sized, irregular, and necrotic in the hippocampal CA1 region, suggesting the occurrence of neuronal degeneration. In addition, we assessed the ultrastructure of neurons by transmission electron microscopy, and found that HAH caused the rupture of membranes, increase in chromatin condensation, and dysregulation of mitochondrial morphology in the hippocampal neurons. To our knowledge, we found the positive effects of oxygen enrichment on the HAH-induced hippocampal damage for the first time, as evidenced by the attenuated morphological abnormalities and improved ultrastructure of neurons in the CA1 region of the hippocampus.
It is generally believed that HAH causes the adverse consequence of the cognitive functions for highlanders, including learning and working memory (Bahrke and Shukitt-Hale, 1993; de Aquino Lemos et al., 2012). In this study, based on the Morris water maze test, we observed that rats exhibited impaired spatial working memory after being exposed to simulated HAH (6,000 m) for 3 days.
Furthermore, several investigators have made efforts to identify the potential effects of oxygen enrichment on the cognitive function via the clinical trials (Luks et al., 1998; Gerard et al., 2000; Moraga et al., 2018), whereas the efficacy remains controversial. Gerard et al. (2000) found oxygen enrichment ameliorated the impairments in cognitive and motor functions at a simulated 5,000 m altitude, including shortened reaction times and improved hand/eye coordination. Moraga et al. (2018) reported that the mental cognitive state and memory of the workers at high altitudes (5,050 m) improved by oxygen enrichment based on the neuropsychological tests. However, Luks et al. (1998) reported that oxygen enrichment was beneficial to improve sleep quality at 3,800 m altitude, whereas no effect on the cognitive function in the following daytime was observed.
Furthermore, all these clinical studies only performed neuropsychological tests without hippocampal evaluations at the tissue or cellular level. In this study, we found that oxygen enrichment remarkably increased the frequency of platform crossing in rats exposed to HAH, indicating the improved cognitive function induced by oxygen enrichment. Coupled with the morphological and microstructural results of the hippocampus, our findings confirmed that oxygen enrichment, as a theoretically promising and technically feasible approach, exhibits therapeutic benefits for HAH-induced cognitive deficits.
Tau protein, a microtubule-associated protein, predominantly expresses in the nerve cells and stabilizes the internal microtubules to keep the cytoskeleton integrated and well-organized (Pîrşcoveanu et al., 2017; Venkatramani and Panda, 2019). The balance between the phosphorylation and dephosphorylation of Tau protein is essential for the normal function of neurons, and an abnormally hyperphosphorylated Tau protein is associated with the pathogenesis of a group of neurodegenerative diseases, such as Alzheimer disease, progressive supranuclear palsy, and Parkinson's disease (Spillantini and Goedert, 2013; Alonso et al., 2016).
Although all phosphorylation sites cooperate in concert to regulate tau toxicity and modulate neurotoxicity (Steinhilb et al., 2007), the function of Tau depends on site-specific phosphorylation (Johnson and Stoothoff, 2004). Among all the phospho-epitopes accessed in this study, Ser262 and Thr231 are recognized to phosphorylate early in the pathological process, while tau phosphorylation at Thr181 and Ser396 appears later in the progression of neurofibrillary tangles (Augustinack et al., 2002).
It has been well established that hypoxia can trigger and aggravate Tau phosphorylation in the brain, and thus leads to subsequent cognitive impairment (Zhang et al., 2014; Raz et al., 2019). In consistent with these results, we observed significantly increased Tau phosphorylation levels at Ser396, Ser262, Thr231, and Thr181 in HAH-exposed rats. Moreover, our results also revealed that oxygen enrichment significantly downregulated the hyperphosphorylation of Tau protein at Ser396, Ser261, Thr231, and Thr181 epitopes induced by HAH. Therefore, oxygen enrichment may be able to reduce Tau-mediated neurotoxicity and lower the risk of neurodegenerative disorders associated with Tau hyperphosphorylation in HAH-exposed rats.
Conclusion
In summary, our results revealed that oxygen enrichment exhibited beneficial effects on the locomotor activity and spatial working memory in rats exposed to acute HAH. Oxygen enrichment treatment has also proven effective to improve the morphology and ultrastructure of the neurons in the CA1 region of the hippocampus. Moreover, oxygen enrichment was found to downregulate the aberrant hyperphosphorylated Tau at multiple phosphorylated epitopes in the CA1 region of the hippocampus.
Taken together, the current study demonstrates that oxygen enrichment can attenuate HAH-induced cognitive impairments, which was associated with improved hippocampal morphology and molecular expression. Our findings indicate that the technique of oxygen enrichment may become a promising alternative treatment against neurodegeneration for workers and travelers ascending to the plateau.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Shaanxi Youth Science and Technology New Star Project (No. 2019KJXX-087) and the Shaanxi Provincial Natural Science Foundation of China (No. 2020SF-084).