
Abstract
Cao, Yinhang, Naoto Fujii, Tomomi Fujimoto, Yin-Feng Lai, Takeshi Ogawa, Tsutomu Hiroyama, Yasushi Enomoto, and Takeshi Nishiyasu. CO2-enriched air inhalation modulates the ventilatory and metabolic responses of endurance runners during incremental running in hypobaric hypoxia. High Alt Med Biol. 23:125–134, 2022.
Aim:
We measured the effects of breathing CO2-enriched air on ventilatory and metabolic responses during incremental running exercise under moderately hypobairc hypoxic (HH) conditions.
Materials and Methods:
Ten young male endurance runners [61.4 ± 6.0 ml/(min·kg)] performed incremental running tests under three conditions: (1) normobaric normoxia (NN), (2) HH (2,500 m), and (3) HH with 5% CO2 inhalation (HH+CO2). The test under NN was always performed first, and then, the two remaining tests were completed in random and counterbalanced order.
Results:
End-tidal CO2 partial pressure (55 ± 3 vs. 35 ± 1 mmHg), peak ventilation (163 ± 14 vs. 152 ± 12 l/min), and peak oxygen uptake [52.3 ± 5.5 vs. 50.5 ± 4.9 ml/(min·kg)] were all higher in the HH+CO2 than HH trial (all p < 0.01), respectively. However, the duration of the incremental test did not differ between HH+CO2 and HH trials.
Conclusion:
These data suggest that chemoreflex activation by breathing CO2-enriched air stimulates breathing and aerobic metabolism during maximal intensity exercise without affecting exercise performance in male endurance runners under a moderately hypobaric hypoxic environment.
Introduction
Peak oxygen uptake (
It is well known that elevating carbon dioxide (CO2) can enhance ventilatory responses by stimulating respiratory chemoreflexes. This is because CO2 that diffuses into the interstitial fluid in the brain and periphery reacts with the water to form carbonic acid (H2CO3), which then dissociates into bicarbonate (HCO3−) and hydrogen ion (H+). The increased H+ then activates both central chemoreceptors in the medulla and peripheral chemoreceptors in the carotid bodies, ultimately increasing ventilation (Dempsey et al., 1984; Duffin, 2005). In line with this, breathing CO2-enriched air increases peak ventilation during maximal exercise at sea level (Babb, 1997; Kato et al., 2005; Fan et al., 2012). On the contrary, the effect of breathing CO2-enriched air on peak ventilation during maximal exercise at high altitude remains unclear, as the results from earlier studies are inconsistent (Subudhi et al., 2011; Fan and Kayser, 2013; Doutreleau et al., 2017).
Furthermore, the three abovementioned studies were all conducted under normobaric or hypobaric hypoxic conditions equivalent to 4,000–5,000 m. No study has yet investigated the effect of breathing CO2-enriched air on peak ventilation under moderately hypobaric hypoxic conditions (e.g., 2,500 m), which is an altitude frequently used by athletes for “live high-train low” model of altitude training (Chapman et al., 2014).
It should be noted that higher peak ventilation during high-intensity exercise increases the work of breathing, which can induce a redistribution of blood flow from the locomotor to respiratory muscles (Harms et al., 1997; Sheel et al., 2001) or exacerbate the perception of dyspnea and respiratory muscle fatigue under normobaric normoxic conditions (Harms et al., 2000; Yates, 2019). These factors may result in an early termination of exercise (Dempsey et al., 2008b; Romer and Dempsey, 2014). Also, arterial oxygen saturation is already near maximal at sea level; hence, increases in ventilation do not mediate a substantial increase in arterial oxygen saturation. Consequently, the increases in peak ventilation and peak aerobic energy production associated with breathing CO2-enriched air, if they occur, do not necessarily translate into improved exercise performance under those conditions.
However, this may not occur under hypobaric hypoxic conditions. Breathing helium–oxygen (He-O2), which has a lower gas density than normobaric air, can reduce airflow resistance and increase peak ventilation,
To address this question, the present study was designed as a mechanistic study to test the hypothesis that breathing CO2-enriched air increases peak ventilation by stimulating chemoreflexes, thereby increasing arterial O2 saturation and oxygen delivery to locomotor muscles, ultimately improving
Materials and Methods
Ethics approval
This study was approved by the Human Subjects Committee of the University of Tsukuba (No. 30-121) and adhered to the guidelines in the Declaration of Helsinki, except for registration in a database. All participants provided written informed consent before participating in the study.
Participants
Ten healthy young male endurance runners participated in this study. Participant characteristics and pulmonary function values are presented in Table 1. All participants were lowlanders with no previous exposure to hypoxic conditions equal to or above 1,000 m for >6 months before participating in the present study. All participants were members of the university track and field team. They were all free from cardiopulmonary disease and were not cigarette smokers.
Participant Characteristics and Pulmonary Function
Values are mean ± SD.
FEV1, forced expired volume in 1 second; FVC, forced vital capacity; MEF25–75, maximal expiratory flow between 25% and 75% of forced vital capacity; MVV, maximal voluntary ventilation; PEFR, peak expiratory flow rate; SD, standard deviation.
Experimental procedure
Participants visited the laboratory before the main experiment and were familiarized with the experimental protocols and equipment. Thereafter, experimental sessions were initiated, in which the participants completed exhaustive incremental running tests in a custom-made environmental chamber (Shimazu, Kyoto, Japan) under three conditions: (1) NN, (2) HH, and (3) HH with CO2-enriched air (5% CO2 [25.7 mmHg], 21% O2 [107.7 mmHg], and balance N2) (HH+CO2). The custom-made environment chamber can simulate hypobaric hypoxic conditions up to 8,000 m above sea level. In addition, the temperature in the chamber can be controlled, and the room air can be continuously ventilated to minimize any increase in CO2 inside the chamber. The three trials were separated by 4–7 days, with the last two trials completed in a random and counterbalanced order. The 5% CO2-enriched air, which can increase peak ventilation under normobaric hypoxia (Doutreleau et al., 2017), was adopted in the present study. The inspired O2 concentration and O2 tension were maintained at their same respective levels in the HH and HH+CO2 trials by adjusting the N2 concentration. The NN trial was always performed first, which enabled us to estimate the total amount of gas that would be inhaled during the HH and HH+CO2 trials.
In all three trials, the participants breathed through the same circuit connected to a large prefilled reservoir held in three to four bags (total 2,100–2,800 l) and were unaware of which gas they were breathing. The gas in the prefilled reservoir was humidified. In the HH+CO2 trial, CO2-enriched air inhalation was initiated at 3 minutes into the 6-minute rest period and was continued until the end of the exercise. Under hypobaric hypoxic conditions, the chamber was gradually decompressed at a rate of ∼130 m/min (∼20 minutes) to achieve a barometric pressure equivalent to that at 2,500 m (560 mmHg). Each exhaustive incremental running test began within 20 minutes after completing the decompression.
Pulmonary function assessment
Pulmonary function was assessed under NN condition using a portable spirometer (AS-507; Minato Medical Science, Osaka, Japan) as previously described (Crapo et al., 1995).
Maximal incremental running tests
On experimental days, all participants performed self-selected 10-minute warm-up exercises (e.g., stretching and jogging) outside the laboratory. The structure of the warm-up was matched for all three conditions. They then entered the environmental chamber, where the room temperature was regulated to 20.2°C ± 0.3°C. The room air was continuously ventilated to minimize any increase of CO2 inside the chamber. After a 6-minute rest period, participants initiated a 2-minute warm-up at a speed of 180 m/min on the treadmill, followed by a 1-minute rest. An exhaustive incremental running test then commenced at a speed of 220 m/min. The running speed was increased by 20 m/min every 2 minutes until 280 m/min was reached, after which the running speed was increased by 10 m/min every 1 minute until volitional fatigue. Participants were verbally encouraged throughout the running test. Near the end of the test, expiratory gases were directly assessed using the Douglas bag method.
We elected to use the Douglas bag method as the measurement error can be large if data were assessed by a mass spectrometer (ARCO-2000; ARCO SYSTEM, Chiba, Japan), especially at maximal intensity exercise. All participants met at least two of the following three criteria for
Measurements
Participants breathed through a face mask attached to a two-way nonrebreathing valve (Hans Rudolph, Shawnee, KS) with the expiration side connected to a Douglas bag (200–250 l) via a smooth bore hose. A pneumotachograph transducer (PN-230; ARCO SYSTEM, Chiba, Japan) and a gas sampling tube with a sampling rate of <0.5 l/min were attached between the mask and the two-way valve, and the expired volume and gases were analyzed using a mass spectrometer (ARCO-2000; ARCO SYSTEM). The temperature of expired gas proximal to the pneumotachograph transducer was measured to express volume parameters (i.e., minute ventilation) under body temperature, pressure, and saturated conditions, whereas metabolic parameters (i.e.,
This information was used to assess peak ventilation,
Estimated respiratory muscle oxygen uptake
Respiratory muscle oxygen uptake (
Data analysis
During rest and the first four stages of the exercise (220–280 m/min), data averaged over the last 1 minute of each stage were used for analysis. Data collected during the last 1 minute of the last stage that participants completed were defined as the peak value. If a participant did not complete the 1-minute stage, data at this speed were not included in the data analysis. The incremental test time was determined as the time from the start of the exercise to the point where the participant could no longer continue to run. In addition, the percent differences in
The relationships between the percent change in
Statistical analyses
Data are presented as mean ± standard deviation (SD). Two-way repeated-measures analysis of variance (ANOVA) was used to compare the submaximal exercise data; the two factors were trial (NN, HH, and HH+CO2) and treadmill speed (220–280 m/min). One-way repeated-measures ANOVA was used to compare values recorded at maximal exercise; the factor was trial (NN, HH, and HH+CO2). When main effects or interactions were detected, post hoc multiple comparisons were made using the Holm–Bonferroni method. Values of p < 0.05 were considered statistically significant. Pearson product moment correlations were used to analyze the relationships between the percent changes in
Results
Respiratory and cardiovascular responses at maximal exercise
As designed, breathing CO2-enriched air increased end-tidal CO2 partial pressure compared with breathing room air (55 ± 3 vs. 35 ± 1 mmHg, p < 0.01, Fig. 1).

End-tidal CO2 partial pressure during maximal exercise. Dotted lines represent each participant's data. Data are mean ± SD. *p < 0.05 versus normobaric normoxia; †p < 0.05 hypobaric hypoxia+CO2 versus hypobaric hypoxia. SD, standard deviation.
In the HH+CO2 trial, peak ventilation was higher than in the HH trial (163 ± 14 vs. 152 ± 12 l/min) (Fig. 2A), and this effect was mediated by an increment in tidal volume compared with HH alone (Fig. 2B) (all p < 0.01). Breathing frequency did not differ between HH and HH+CO2 trials (Fig. 2C). Moreover, at maximal exercise,

Peak ventilation
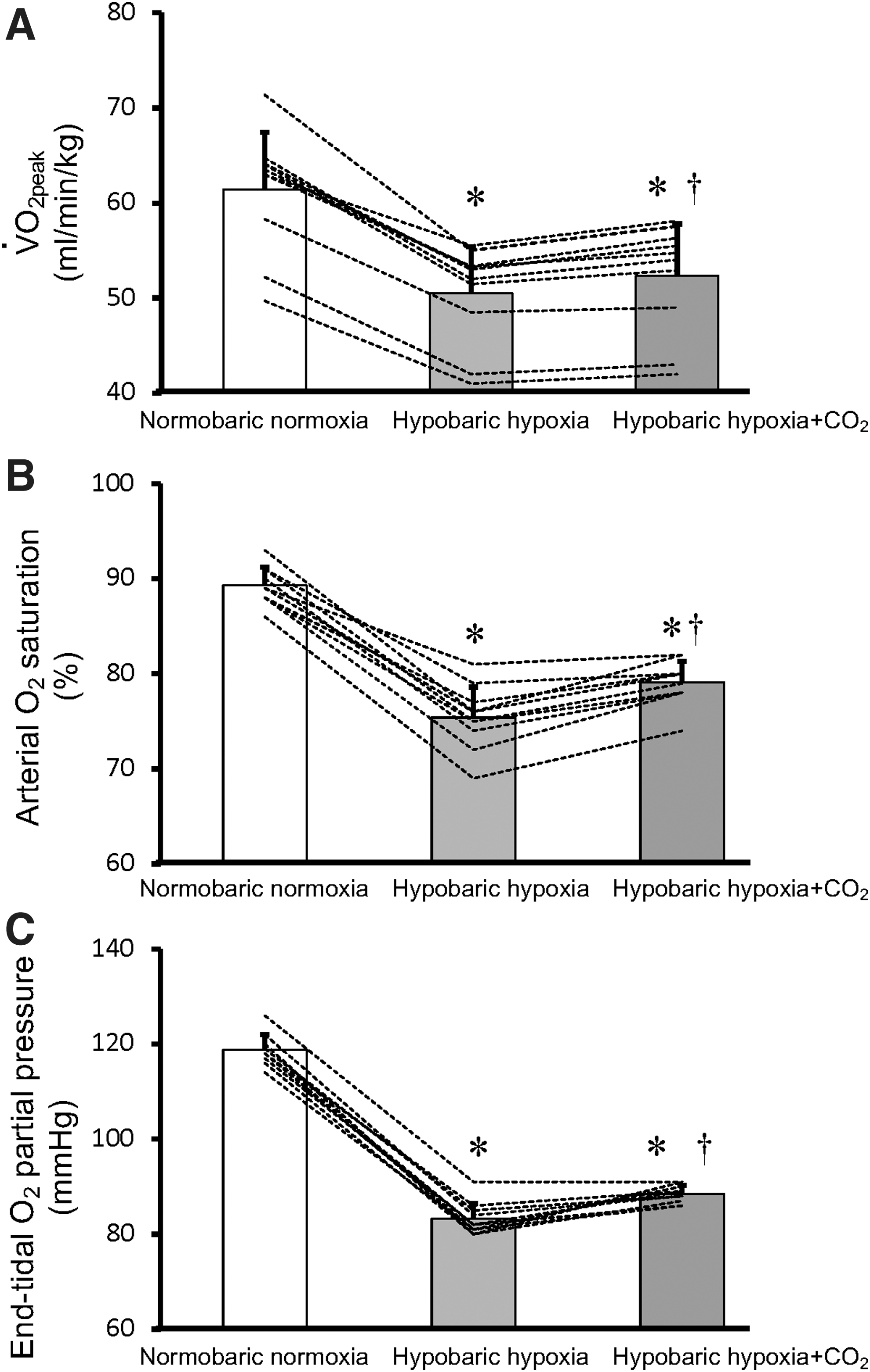
Peak oxygen uptake (
In addition, the percent increase in
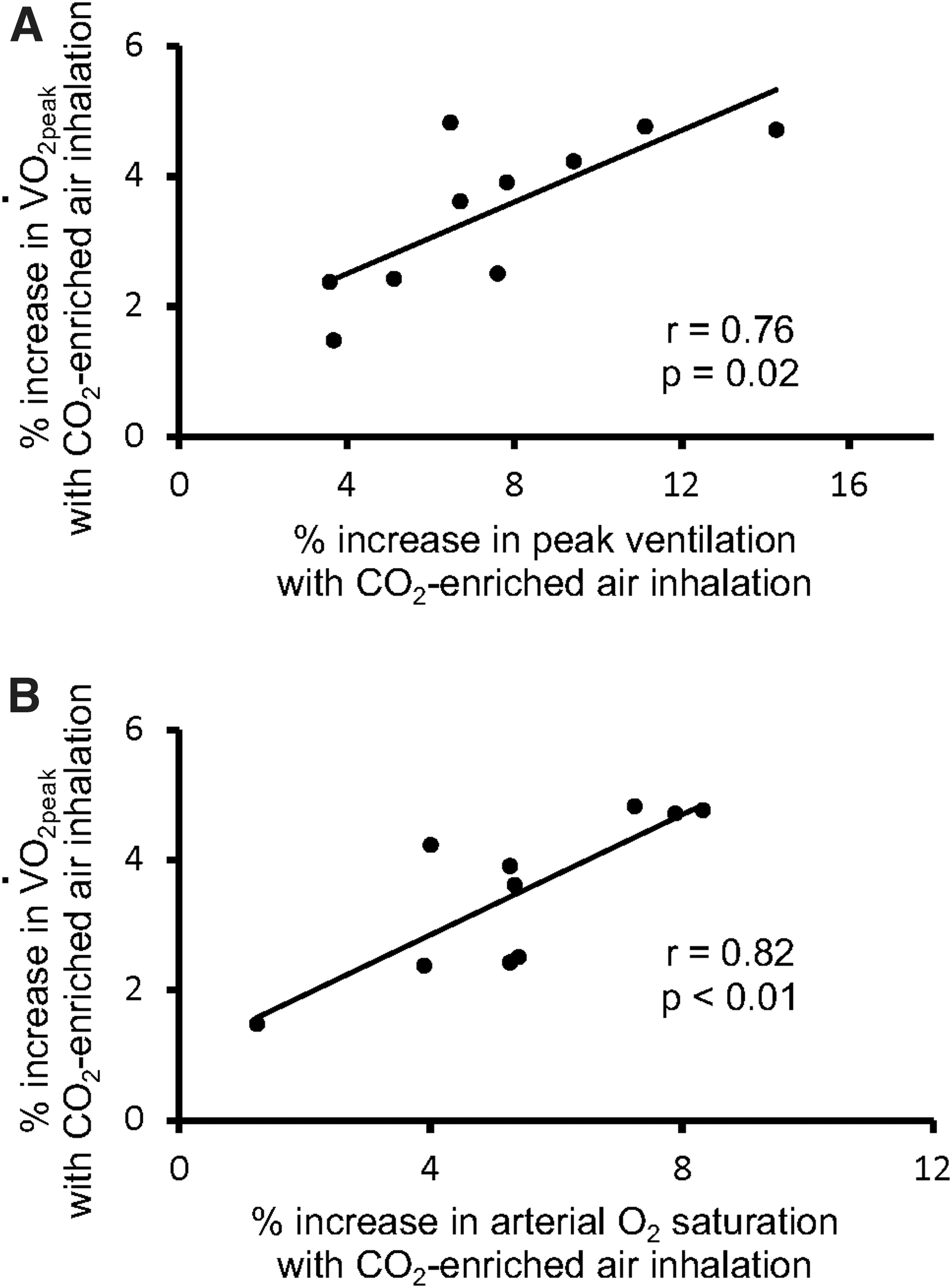
Relationships between the percent increase in peak oxygen uptake (
Ventilatory equivalents for O2 (p < 0.01) and CO2 (p < 0.01) were higher, whereas the respiratory exchange ratio was lower at maximal exercise in the HH+CO2 trial than in the HH trial (Table 2). Peak CO2 output (p = 0.10) and peak heart rate (p = 0.73) did not differ between the HH and HH+CO2 trials (Table 2).
Variables Obtained During Maximal Exercise
Values are mean ± SD.
p < 0.05 versus normobaric normoxia.
p < 0.05 hypobaric hypoxia+CO2 versus hypobaric hypoxia.
Perceptions of dyspnea and exertion at maximal exercise
Perception of dyspnea at maximal exercise was higher in the HH+CO2 trial than in the HH trial (p = 0.02, Table 2). There was no difference in the rating of perceived exertion between the HH and HH+CO2 trials (p = 0.89, Table 2).
Incremental test time
Incremental test time did not differ between the HH and HH+CO2 trials (p = 0.25, Fig. 5).
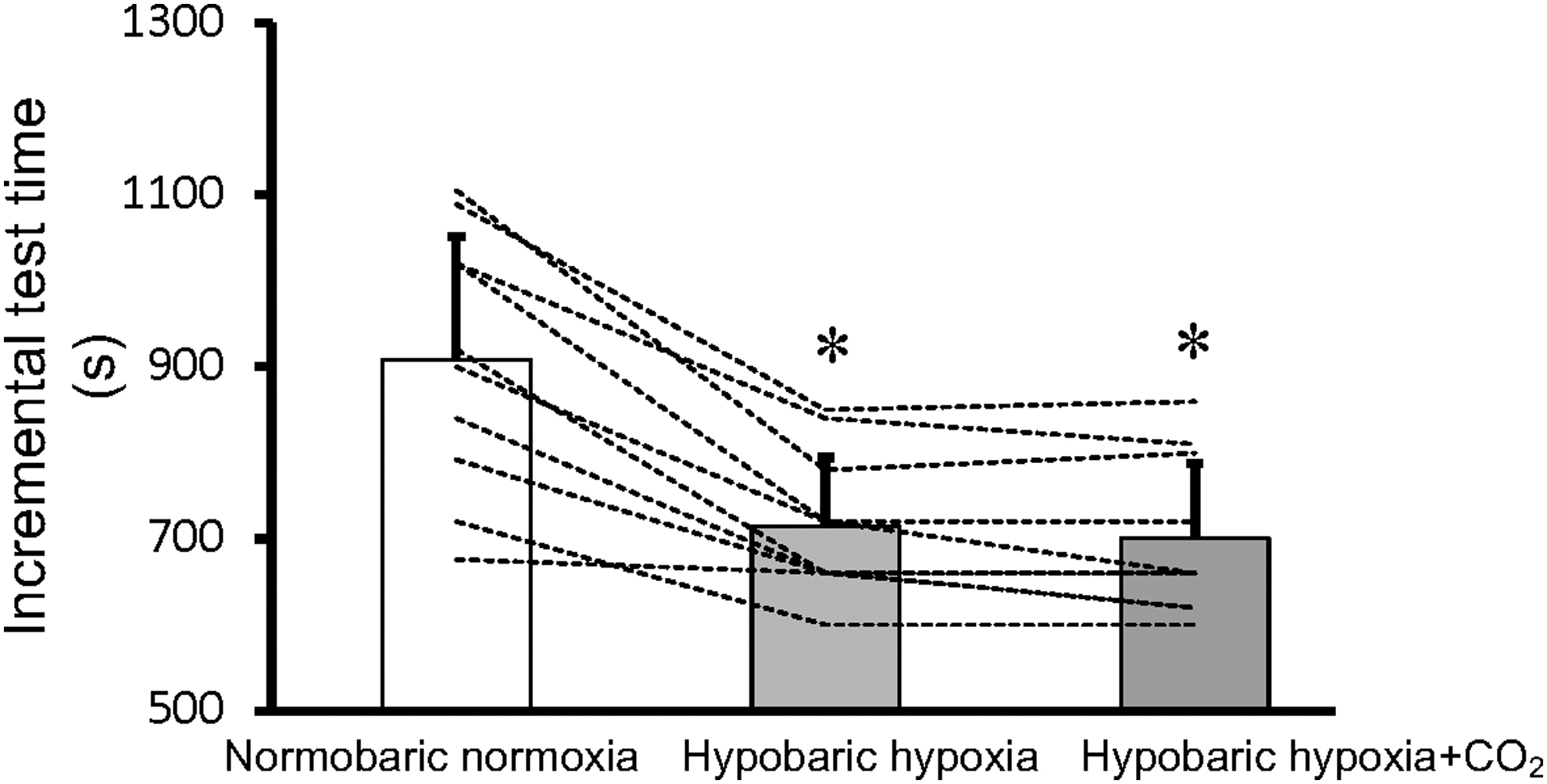
Total exercise time. Data are mean ± SD. *p < 0.05 versus normobaric normoxia. Dotted lines represent each participant's data.
Estimated respiratory muscle oxygen uptake at maximal exercise
In parallel with the increase in peak ventilation from the HH to the HH+CO2 trials (Δ11 ± 5 l/min), the estimated
Respiratory and cardiovascular responses during rest and submaximal exercise
In resting subjects, minute ventilation, tidal volume, arterial O2 saturation, end-tidal O2 and CO2 partial pressures, and ventilatory equivalents for O2 and CO2 were all higher, whereas CO2 output and respiratory exchange ratio were lower in the HH+CO2 than in the HH trial (Table 3, all p < 0.05). In addition, minute ventilation, tidal volume,
Variables Obtained During Rest and Submaximal Exercise
Values are mean ± SD.
p < 0.05 versus normobaric normoxia.
p < 0.05 hypobaric hypoxia+CO2 versus hypobaric hypoxia.
NA, not available;
Perceptions of dyspnea and exertion during submaximal exercise
Perceptions of dyspnea and exertion at running speeds of 260 and 280 m/min were also higher in the HH+CO2 than in the HH trial (all p < 0.05, Table 3).
Discussion
The major findings of the present study are as follows: (1) breathing 5% CO2-enriched air increases end-tidal CO2 partial pressure, peak ventilation, and
Ventilatory response
Consistent with an earlier study on the effects of normobaric hypoxia (4,000 m) (Doutreleau et al., 2017), we found that breathing CO2-enriched air increased peak ventilation during maximal exercise (Fig. 2A). This may be the result of H+-mediated central and peripheral chemoreceptor activation related to the increased CO2, which combines with water to form H2CO3 and then ionizes to HCO3− and H+ (Dempsey et al., 1984; Duffin, 2005). However, in contrast to our findings and those of Doutreleau et al. (2017), two earlier studies reported no increase in peak ventilation with end-tidal CO2 partial pressure clamped at 45–50 mmHg under HH or normobaric hypoxia (4,875 or 5,000 m) (Subudhi et al., 2011; Fan and Kayser, 2013).
A possible explanation for the disparate findings is differences in the level of end-tidal CO2 partial pressures. The end-tidal CO2 partial pressures in the present study and the study by Doutreleau et al. (2017) were 55–60 mmHg, which are higher than in the other two studies (Subudhi et al., 2011; Fan and Kayser, 2013). This suggests that end-tidal CO2 partial pressures greater than 50 mmHg may be necessary to increase peak ventilation in a moderately hypobaric hypoxic environment. Alternatively, the different levels of hypoxia may underlie the disparate results. The level of hypoxia achieved in the two earlier studies (4,875–5,000 m) was greater than in the present study and the study by Doutreleau et al. (2017) (4,000 m).
In addition, it has been reported that another respiratory neural mechanism independent of chemoreceptor feedback, defined as short-term modulation, is also involved in the regulation of exercise ventilatory response to maintain normal blood gas homeostasis (Wood et al., 2008). Furthermore, Dempsey and Smith (1994) proposed that enhanced ventilatory response during high-intensity exercise may be, in part, due to the increased descending activity from the locomotor-linked central nervous system. Thus, the role of short-term modulation and feedforward mechanisms in increasing peak ventilation cannot be ruled out in the present study.
During maximal or near-maximal exercise under normobaric normoxic conditions, the ventilatory responses of some endurance athletes may be restricted because of mechanical limitations classically termed expiratory flow limitation (Johnson et al., 1992; Dominelli et al., 2012). However, we previously observed that expiratory flow limitation does not affect ventilatory responses during incremental running in a moderate hypobaric hypoxic environment (2,500 m) (Cao et al., 2019). Thus, it does not appear that expiratory flow limitation, if there was any, affected ventilatory responses in the present study.
Metabolic response and exercise performance
As we hypothesized, there was an increase in
Despite the elevated provision of aerobic energy in the HH+CO2 trial, as reflected by the higher
This notion is supported by our results showing that the peak heart rate in the HH+CO2 trial was slightly lower than that in the HH trial (Table 2). Third, the lower pH associated with CO2-enriched air inhalation may shift the hemoglobin O2 dissociation curve to the right, decreasing arterial O2 saturation for a given arterial O2 partial pressure (Doutreleau et al., 2017). Thus, the Bohr effect may prevent elevations in arterial O2 saturation and O2 content. Fourth,
Previous studies reported that increased work of breathing with higher minute ventilation may result in an early termination of exercise under normobaric normoxic conditions (Dempsey et al., 2008b; Romer and Dempsey, 2014). However, our results demonstrated that breathing CO2-enriched air increased peak ventilation and
Furthermore, Esposito and Ferretti (1997) reported that breathing He-O2 increased peak ventilation,
Variables during submaximal exercise
Consistent with earlier studies (Subudhi et al., 2011; Doutreleau et al., 2017), we found that breathing CO2-enriched air increased end-tidal CO2 and O2 partial pressures, minute ventilation, and arterial O2 saturation, whereas it had no effect on heart rate or CO2 output during maximal and submaximal intensity exercise (Table 3). Thus, breathing CO2-enriched air appears to modulate ventilatory and metabolic responses independently of exercise intensity. Because breathing CO2-enriched air increased the submaximal arterial O2 saturation, it may enhance prolonged submaximal exercise performance (e.g., duration of exercise) under hypobaric hypoxic conditions, but this needs to be directly assessed in the future.
Limitations
As the calculation of estimated
Fifth, only young males were included in the present study, which limits the generalizability of our findings. In comparison with males, females have smaller lungs and conducting airways (Sheel et al., 2009). Consequently, expiratory flow limitation is a factor limiting exercise ventilation in females more often than in males (Guenette et al., 2007). Therefore, the increase in peak ventilation with breathing CO2-enriched air may be smaller in females than in males. This warrants future investigation.
Conclusion
Chemoreflex activation by breathing CO2-enriched air was shown to augment ventilatory and metabolic responses, but the incremental exercise performance by competitive endurance runners remained unchanged under moderately hypobaric hypoxic conditions (2,500 m above sea level).
Footnotes
Acknowledgments
We sincerely thank the participants for their time and efforts to participate in this study.
Authors' Contributions
C.Y., N.F., F.T., T.O., and T.N. conceived and designed the experiments. C.Y., F. T., and L.Y. contributed to data collection. C.Y. and N.F. performed the data analysis. C.Y., N.F., F.T., L.Y., T.O., T.H., Y.E., and T.N. interpreted the experimental results. C.Y. prepared the figures. C.Y. drafted the article. C.Y., N.F., F.T., L.Y., T.O., T.H., Y.E., and T.N. edited and revised the article. All authors approved the final version of the article, and all experiments took place at the Institute of Health and Sport Sciences located at the University of Tsukuba.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the Ministry of Education, Culture, Sports, Science and Technology in Japan and the “Human High Performance (HHP) Research Project,” at the University of Tsukuba, and supported by Shanghai Key Lab of Human Performance at Shanghai University of Sport (No. 11DZ2261100).