
Abstract
Liu, Bo, Minlan Yuan, Mei Yang, Hongru Zhu, and Wei Zhang. The effect of high-altitude hypoxia on neuropsychiatric functions. High Alt Med Biol. 25:26–41, 2024.
Background:
In recent years, there has been a growing popularity in engaging in activities at high altitudes, such as hiking and work. However, these high-altitude environments pose risks of hypoxia, which can lead to various acute or chronic cerebral diseases. These conditions include common neurological diseases such as acute mountain sickness (AMS), high-altitude cerebral edema, and altitude-related cerebrovascular diseases, as well as psychiatric disorders such as anxiety, depression, and psychosis. However, reviews of altitude-related neuropsychiatric conditions and their potential mechanisms are rare.
Methods:
We conducted searches on PubMed and Google Scholar, exploring existing literature encompassing preclinical and clinical studies. Our aim was to summarize the prevalent neuropsychiatric diseases induced by altitude hypoxia, the potential pathophysiological mechanisms, as well as the available pharmacological and nonpharmacological strategies for prevention and intervention.
Results:
The development of altitude-related cerebral diseases may arise from various pathogenic processes, including neurovascular alterations associated with hypoxia, cytotoxic responses, activation of reactive oxygen species, and dysregulation of the expression of hypoxia inducible factor-1 and nuclear factor erythroid 2-related factor 2. Furthermore, the interplay between hypoxia-induced neurological and psychiatric changes is believed to play a role in the progression of brain damage.
Conclusions:
While there is some evidence pointing to pathophysiological changes in hypoxia-induced brain damage, the precise mechanisms responsible for neuropsychiatric alterations remain elusive. Currently, the range of prevention and intervention strategies available is primarily focused on addressing AMS, with a preference for prevention rather than treatment.
Introduction
In recent years, there has been a surge in the popularity of engaging in high-altitude activities such as hiking, climbing, and work (Luks and Hackett, 2022; Williamson et al., 2018). These high-altitude environments are known for their challenging conditions, including reduced atmospheric pressure, low oxygen levels, increased ultraviolet radiation, and significant temperature variations (Parodi et al., 2022). Among these conditions, hypoxia poses the most significant concern as it can give rise to various altitude-related illnesses such as acute mountain sickness (AMS), high-altitude cerebral edema (HACE), and high-altitude pulmonary edema (Basnyat and Murdoch, 2003; Peacock, 1998).
The classification of high altitude is widely accepted, with divisions into high altitude (1,500–3,500 m), very high altitude (3,500–5,500 m), and extreme high altitude (above 5,500 m) (Wilson et al., 2009). Among these divisions, the altitude of 2,500 m is considered the threshold for AMS and 8,000 m is known as the “death line” (Lankford, 2021; Luks et al., 2017). With every 100-m increase in altitude, the barometric pressure (PB) decreases by ∼5 mmHg, subsequently leading to a corresponding reduction of 1 mmHg in pressure of inspired oxygen (P

The relationship between altitude, PB, and inspired oxygen pressure. Figure adapted from Wilson et al. (2009). P
To counter the challenge of hypoxia, the body uses compensatory mechanisms to maintain homeostasis. These include hyperventilation (Schneider et al., 2021), tachycardia (Zhang et al., 2014), redistribution of blood flow (Kamitomo et al., 1993), metabolic adaptations (Chicco et al., 2018), and increased red blood cell or hemoglobin production (Ryan et al., 2014). However, if these adaptive strategies prove ineffective, the respiratory system (Swenson and Bärtsch, 2012), cardiovascular system (Holloway et al., 2011), and nervous system (Swenson, 2020) become the primary targets of hypoxia.
This review primarily focuses on hypoxia-induced brain damage. Despite the brain's small weight accounting for only 2% of the total body weight, it requires a substantial amount of energy, ∼20%, owing to its extensive neural network comprising around 100 billion neurons, a comparable number of glial cells (von Bartheld et al., 2016), and trillions of intricate synapses (Goyal et al., 2014; Magistretti and Allaman, 2015). Even brief episodes of hypoxia lasting less than 5 minutes can lead to a significant depletion of adenosine triphosphate (ATP), up to 90%, in brain tissue (Mukandala et al., 2016). Consequently, numerous neurons and their synaptic connections may undergo degeneration (Bailey et al., 2009a).
In addition to the well-recognized acute altitude headache and HACE, hypoxia has been implicated in various chronic neurophysiological disorders, including cerebrovascular diseases (Liu et al., 2021) and neuropsychiatric disorders (Wilson et al., 2009), including alterations in perception (Hüfner et al., 2018), cognition (Sharma et al., 2019; Wang et al., 2019; Wu et al., 1998), emotion (Kious et al., 2019), and behaviors (Paulus et al., 2009) as well as sleep disorders (Bolmont et al., 2000; Dos Santos Quaresma et al., 2020). The potential pathogenesis of altitude-related cerebral diseases is believed to involve hypoxia-related neurovascular alterations, cytotoxic responses, reactive oxygen species (ROS) activation, and dysregulation of expression involving hypoxia inducible factor-1 and nuclear factor erythroid 2-related factor 2 (Nrf2) (Choudhry and Harris, 2018; Lafuente et al., 2016; Lee et al., 2020).
This review aims to provide a summary of recent findings in neuropsychiatric diseases associated with high-altitude hypoxia, with a primary focus on neuropsychiatric disorders, potential pathophysiological mechanisms, and both pharmacological and nonpharmacological strategies for prevention and intervention.
Methods
A comprehensive literature search was conducted to include relevant sources related to brain damage and high-altitude hypoxia. The search was carried out in English using PubMed and Google Scholar, utilizing keywords such as “brain damage/injury and high altitude hypoxia” or “mental illness at high altitude.” In addition, information from reputable websites focusing on high altitudes was also referenced to ensure a thorough review.
Neurological Diseases Associated with Altitude Hypoxia
While high-altitude headache, AMS, and HACE are commonly recognized as three successive stages of acute high-altitude brain damage, chronic mountain sickness (CMS) is characterized by the presence of polycythemia and hypoxemia (Villafuerte and Corante, 2016). Clinically, CMS is manifested by symptoms such as fatigue, breathlessness, headache, cyanosis, and sleep disturbances. A diagnosis of CMS is typically established when the Qinghai CMS Score exceeds 5 (Gonzales et al., 2013).
Several cerebrovascular diseases have been reported in relationship to high altitude, including transient ischemic attack, cerebral infarction, and cerebral hemorrhage (Jha et al., 2002; Zhu et al., 2021). Transient ischemic attacks occurring during ascent are believed to be linked to regional cerebral vasoconstriction (Richalet et al., 2020; Wohns, 1986). Cases of cerebral venous infarction (Kim et al., 2017) or thrombosis (Hassan et al., 2019; Song et al., 1986) at high altitude have also been documented. Therefore, when patients present with focal neurological symptoms like hemiparesis in addition to symptoms of AMS or HACE, cerebral ischemic stroke should be considered.
Furthermore, individuals at high altitudes who experience cerebral venous thrombosis often exhibit elevated hemoglobin levels, with an average value of 16.7 g/dL, and ∼50% of them have increased d-dimer levels exceeding 1,000 ng/ml, implying that the development of blood hypercoagulability may contribute to cerebral venous thrombosis (Hassan et al., 2019; Roach and Hackett, 2001). In addition, factors such as dehydration, hemoconcentration, tight clothing, low temperature, and limited physical activity in extreme conditions can also contribute to the occurrence of cerebral venous thrombosis (Gupta and Ashraf, 2012). Moreover, intracerebral hemorrhage in high-altitude regions has gained attention in recent years. A multicenter study conducted on the Qinghai-Tibet Plateau revealed that most cases of cerebral hemorrhage were associated with hypertension and exhibited a higher cardiovascular risk profile (Yuan et al., 2016).
Numerous case reports provide evidence of various neurological conditions associated with high altitudes, including chronic pain, seizures, syncope, cranial nerve palsy, and cortical atrophy (Falla et al., 2021; Wilson et al., 2009). A cross-sectional study involving 1,095 individuals residing at high altitudes found that 31.69% of the population experienced musculoskeletal pain (Vega-Hinojosa et al., 2018). In comparison, an epidemiological study reported that the prevalence of chronic musculoskeletal pain in the general population ranged from 13.5% to 47%, with factors such as pain type, age, sex, education, household income, smoking, anxiety, depression, and sleep disorders being correlated (Cimmino et al., 2011). Witness accounts and positive family history confirmed new episodes of altitude-induced seizures, suggesting that individuals with a predisposition to seizures may face an increased risk at higher elevations (Daleau et al., 2006).
High-altitude hypoxic syncope is commonly attributed to vasovagal mechanisms resulting from hypoxemia (Chiang et al., 2012), although the increased arrhythmias might be a risk factor (Woods et al., 2008). Cranial nerve palsy can occur with or without AMS or HACE (Falla et al., 2021). Although the sixth cranial nerve is most often impaired partly due to due to compression from brain swelling in adjacent regions, other cranial nerves may also be damaged (Blomquist, 2018). Reports have documented cases of facial and hypoglossal cranial nerve palsies as well (Basnyat et al., 2004). For instance, a 55-year-old woman experienced paresis of the third cranial nerve involving the pupil at high altitude (Blomquist, 2018). Furthermore, magnetic resonance imaging (MRI) has revealed altitude-induced cortical atrophy (Di Paola et al., 2008), frontal subcortical lesions (Fayed et al., 2006), and leukoaraiosis (Virués-Ortega et al., 2004) characterized by demyelination, axonal degeneration, and astrocyte hyperplasia.
Psychiatric Disorders Related to Altitude Hypoxia
Perceptual changes
Emerging evidence suggests that individuals exposed to high-altitude environments may experience impairments in perception. Studies have reported a lowered threshold for detecting various physical sensations such as smell, taste, and pain in high-altitude areas (Noël-Jorand et al., 1996; Virués-Ortega et al., 2004). A clinical study examining long-term exposure to high altitudes found a significant alteration in blue and red color perception among both local residents and migrants, although the underlying cause remains unknown (Wang et al., 2019). Conversely, an increase in the threshold for detecting visual stimuli was observed in dark-adapted individuals experiencing hypoxia, likely due to reduced visual sensitivity (Kobrick and Appleton, 1971; West, 2004). Interestingly, climbers exposed to very high or extreme altitudes have reported experiencing hallucinations, including somesthetic, visual, and auditory pseudohallucinations (Brugger et al., 1999; Hüfner et al., 2021).
Cognitive changes
Accompanying the perceptual impairments, high-altitude exposure has been associated with various cognitive changes, including learning, attention, memory, reaction time, arithmetic ability, logical reasoning, decision-making, and problem-solving (de Aquino Lemos et al., 2012; Koester-Hegmann et al., 2018; Kramer et al., 1993; West, 2004). For instance, climbers, when compared to matched control groups at sea level, have exhibited deficits in learning and memory tasks, partially attributed to the sensitivity of the hippocampus to hypoxia (Kramer et al., 1993). Furthermore, under hypobaric-hypoxic conditions, the P300 latency of event-related potentials was significantly prolonged, which returned to baseline levels after oxygen supplementation at 4,500 m (Hayashi et al., 2005). Similarly, acute exposure to hypoxic conditions resulted in decreased pupil function, reflecting cerebral function, which then returned to normal after acclimatization (Wilson et al., 2008).
Functional near-infrared spectroscopy studies have reported impaired speed and accuracy of information processing at high altitudes, leading to slower reaction times and higher decision errors (Davranche et al., 2016). In line with clinical studies, mice exposed to simulated altitudes equivalent to 5,000 m for 12 weeks exhibited deficits in learning and memory, along with alterations in the expression of synaptic proteins in relevant brain regions (Sharma et al., 2019).
Emotional changes
In addition to cognitive impairments, a range of emotional problems can manifest during high-altitude exposure, including anxiety, depression, euphoria, irritability, and hostility (Bahrke and Shukitt-Hale, 1993; Dos Santos Quaresma et al., 2020). Studies have observed an increase in negative emotions and a decrease in positive emotions with higher altitudes (Li et al., 2000). The emotional state is influenced by factors such as the duration of exposure to hypoxia, ascent speed, and individual differences (Dos Santos Quaresma et al., 2020). Anxiety episodes are relatively common during ascent to high altitudes, as reported by individuals (Fagenholz et al., 2007).
Surveys conducted among military volunteers stationed in Pakistan's Karakoram Mountains revealed that those deployed at high altitudes experienced anxiety and depression (Ahmad and Hussain, 2017). Similarly, data from the Intern Health Study indicated that individuals who relocated to high altitudes were more likely to experience depression, anxiety, and suicidal ideation over the course of a year (Kious et al., 2019). Consistent with clinical studies, experiments on mice exposed to simulated high altitudes have shown depression-like symptoms (Kanekar et al., 2015).
Behavioral changes
Psychomotor performance impairment has been observed in high-altitude hypoxic environments, as demonstrated by tests such as the Finger Tapping Test (Austin et al., 2015) or Purdue Pegboard Test (Akoudad et al., 2016). Some researchers propose that these effects may be attributed to altitude-related fatigue and anxiety. Conversely, others argue that psychomotor impairment could be a direct consequence of altitude, as studies have shown persistent motor impairment even after returning to sea level (Virués-Ortega et al., 2006). It is widely believed that in extreme conditions, behavior tends to become more intuitive and simplistic for survival. Moreover, several countries, including South Korea, Austria, and the United States, have documented a positive correlation between altitude and suicide rates, although the underlying reasons for this association remain unclear (Brenner et al., 2011; Haws et al., 2009; Kanekar et al., 2015).
Sleep disturbance
Sleep disturbance is a prevalent symptom experienced at high altitudes, often characterized by increased sleep fragmentation due to frequent brief arousals and the occurrence of periodic breathing (PB) (Jafarian et al., 2008; Weil, 2004). Consequently, there is a reduction in total sleep time and sleep efficiency (de Aquino Lemos et al., 2012). However, recent research has raised some controversy regarding the association between arousal, light sleep, and PB (Windsor and Rodway, 2012). Changes in sleep architecture typically involve an increase in light sleep stages and a decrease in slow-wave sleep and rapid eye movement (REM) sleep (de Aquino Lemos et al., 2012; Weil, 2004).
A study comparing the sleep patterns of partially acclimatized lowland soldiers with Tibetan soldiers stationed at Shangri-La (3,800 m) revealed that the former experienced less non-REM sleep and encountered difficulties in sleep initiation compared to the latter (Kong et al., 2015). Furthermore, it has been suggested that nocturnal hypoxemia accompanied by sleep fragmentation can contribute to daytime cognitive impairment (Mansukhani et al., 2019).
The Mechanisms of Hypoxia-Related Neuropsychiatric Conditions
Redox system imbalance
The redox system comprised a diverse range of pro-oxidants and antioxidants, and maintaining its homeostasis is crucial for aerobic life. ROS, a prominent pro-oxidant, are primarily generated in the electron transport chain (ETC) during mitochondrial respiration (Lee et al., 2020). The ETC consists of four complexes (I, II, III, and IV), with complex III being particularly implicated in ROS generation under hypoxic conditions, although the precise mechanisms are not fully understood (Lee et al., 2020). However, an alternative perspective suggests that some electrons directly react with oxygen, leading to ROS production (Zhao et al., 2019). ROS exhibit a dual nature in biological systems (Buonocore et al., 2010). On one hand, they play a crucial role in molecular signaling pathways, such as proliferation, differentiation, and oxidative modifications of proteins (Gaur et al., 2021). On the other hand, ROS can oxidize biological macromolecules such as proteins, lipids, and DNA (Dosek et al., 2007).
Consequently, the production of ROS is tightly regulated (Fuhrmann and Brüne, 2017). Interestingly, it is widely accepted that the overproduction of ROS is primarily associated with hyperoxia (Resseguie et al., 2015). However, under hypoxic conditions, mitochondria dysfunction can also lead to the excessive generation of ROS (Chandel et al., 1998; Gaur et al., 2021; Lee et al., 2020). As the cell's primary consumer and sensor of oxygen, mitochondria are particularly affected when oxygen levels are reduced, thereby triggering the production of substantial ROS (Chandel et al., 1998). Hypoxia promotes the transfer of electrons to oxygen by increasing the expression of a complex IV (COX) subunit, which helps maintain energy homeostasis within cells (Lee et al., 2020). ROS can also elevate calcium (Ca2+) levels, activating the CaMKK-AMPK pathway and reducing metabolic demand in response to prolonged hypoxia (Lee et al., 2020).
In high-altitude conditions, where ambient oxygen availability is diminished, the excessive production of ROS leads to heightened oxidative stress and disrupts the balance of the redox system (Gaur et al., 2021). Moreover, it has been reported that ROS production is positively associated with altitude (Gaur et al., 2021). Excessive ROS can result in mitochondrial dysfunction and initiate the cascade of cell death (Angelova and Abramov, 2018). In the brain, ROS can contribute to neuronal loss and various neurodegenerative diseases, as well as cerebral edema (Angelova and Abramov, 2018). Conversely, hypoxia-induced mitochondrial autophagy, regulated by hypoxia-inducible factor (HIF)-mediated expression of BCL2 interacting protein 3-like (BNIP3L), serves as an adaptive metabolic response to prevent excessive ROS levels and maintain cell survival (Bellot et al., 2009; Zhang et al., 2008). Excessive ROS further amplifies the oxidative stress response and disrupts the redox balance, ultimately leading to brain injury (Gaur et al., 2021) (Fig. 2).
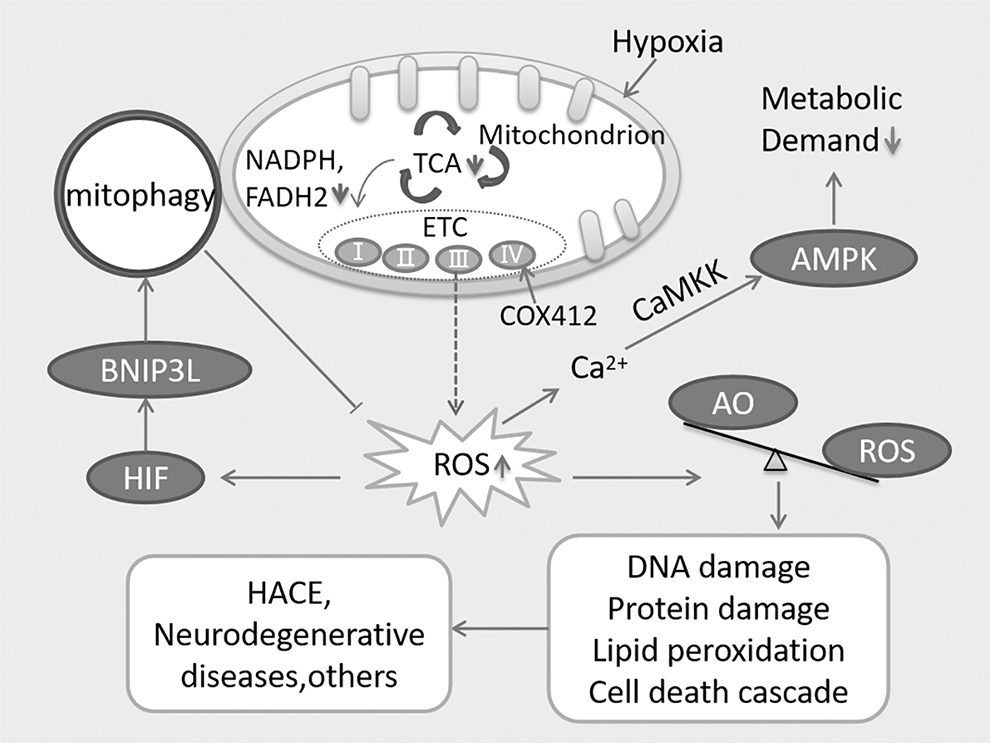
Hypoxia-induced ROS production and redox systems imbalance. Figure adapted from Lee et al. (2020). The dysfunction of ETC causes an increase in ROS production, in turn leading to the imbalance between AO and ROS, finally resulting in HACE and other cerebral diseases via oxidative damages, such as DNA damage, protein damage, and lipid peroxidation. Excessive HIF boosts the upregulation of BNIP3L, which further induces mitophagy and reduces the production of ROS. Hypoxia-induced expression of COX412 enhances the transfer of electrons. ROS can also induce the cumulation of Ca2+, further activates the CaMKK-AMPK pathway and decreases the metabolic demand. AMPK, AMP-activated protein kinase; AO, antioxidants; BNIP3L, BCL2 interacting protein 3 like; CaMKK, Ca2+/calmodulin-dependent protein kinase; COX412, a complex IV (COX) subunit; ETC, electron transport chain; HACE, high-altitude cerebral edema; HIF, hypoxia inducible factor; ROS, reactive oxygen species; TCA, tricarboxylic acid cycle.
Expression dysregulation of hypoxia-related genes
HIF is a transcription factor composed of HIF-α and HIF-β subunits (Semenza, 1999). Under normal conditions, HIF-α is hydroxylated by prolyl hydroxylases in the cytoplasm and subsequently degraded by ubiquitination mediated by Von Hippel–Lindau proteins (Kaelin and Ratcliffe, 2008; Lee et al., 2020; Lendahl et al., 2009). However, when cellular oxygen levels decrease or ROS levels increase, this degradation process is inhibited (Burtscher et al., 2021). Consequently, HIF-α translocates to the nucleus, where it forms a heterodimer with HIF-β and binds to hypoxia response elements in the promoter regions of HIF target genes, such as those encoding glucose transporters (GLUT), glycolytic enzymes (GE), erythropoietin (EPO), vascular endothelial growth factor (VEGF), and others (Burtscher et al., 2021; Semenza, 1999; Semenza et al., 1994). This binding enhances the transcription of these genes.
The increased expression and translocation of GLUT to the plasma membrane during hypoxia are associated with low intracellular ATP levels, leading to increased glucose uptake (Mamun et al., 2020). Due to reduced oxidative phosphorylation under hypoxic conditions, cells shift to the glycolytic pathway to meet their ATP demands, necessitating the upregulation of GE (Kierans and Taylor, 2021; Lendahl et al., 2009). EPO upregulation improves oxygen-carrying capacity by promoting erythropoiesis at high altitudes (Haase, 2013). The upregulation of VEGF expression contributes to alleviating inadequate oxygen supply by promoting angiogenesis and increased blood supply. These molecular changes play a crucial role in coordinating the oxygen supply-demand relationship and ultimately facilitate acclimatization to altitude hypoxia conditions (Hochachka et al., 1996) (Fig. 3).

Expression regulation of HIF and Nrf2 in response to hypoxia. Figure adapted from Burtscher et al. (2021). Hypoxia-induced ROS blocks the degradation of HIF-α and Nrf2 by proteasome and boosts them translocation into nucleus. After binding to their partners (HIF-β or Maf respectively), they further activate the expression of related genes, heightening the adaptation of cells in response to hypoxia. ARE, antioxidant response elements; ATP, adenosine triphosphate; EPO, erythropoietin; GE, glycolytic enzymes; GLUT, glucose transporters; GPx, glutathione peroxidase; GR, glutathione reductase; H2O2, hydrogen peroxide; HRE, hypoxia response element; HIF-α, hypoxia inducible factor α-subunit; HIF-β, hypoxia inducible factor β-subunit; Keap1, Kelch-like ECH-associated protein 1; Maf, musculoaponeurotic fibrosarcoma protein; Nrf2, nuclear factor erythroid 2-related factor 2; O2, oxygen; O2•−, superoxide radical; PHD, prolyl hydroxylase; SOD, superoxide dismutase; VEGF, vascular endothelial growth factor; VHL, Van Hippel–Lindau proteins.
Furthermore, the hypothesis that HIF-1 may be associated with mood disorders has been explored by Shibata et al. (2013), who observed increased expression of HIF-1 messenger RNA in peripheral blood cells of patients with mood disorders, suggesting the involvement of HIF-1 in the pathophysiology of depression.
Nrf2 acts as a critical transcription factor that regulates cellular redox states and activates the transcription of antioxidant and anti-inflammatory genes (Baird and Yamamoto, 2020; Sharma et al., 2020; Vasconcelos et al., 2019). Under normal conditions, Nrf2 is bound to Kelch-like ECH-associated protein 1 (Keap1) in the cytoplasm, leading to its degradation. However, in response to hypoxia-induced ROS, Nrf2 is released from Keap1, allowing it to translocate into the nucleus, where it forms a complex with musculoaponeurotic fibrosarcoma protein (Maf). This complex then binds to antioxidant response elements and activates the transcription of antioxidant and anti-inflammatory genes, including key enzymes such as glutathione reductase (GR), glutathione peroxidase (GPx), and superoxide dismutase (SOD) (Zhang et al., 2013). GR plays a crucial role in maintaining a sufficient supply of reduced glutathione (GSH), which is vital for regulating cellular ROS levels (Couto et al., 2016).
SOD catalyzes the dismutation of the superoxide radical (O2•−) into oxygen (O2) and hydrogen peroxide (H2O2), while GPx reduces H2O2 to water, mitigating the harmful effects of ROS (Margis et al., 2008; Miao and St Clair, 2009). This cascade of events enhances the antioxidant capacity and improves adaptability to high-altitude hypoxia (Fig. 3). Therefore, these signaling pathways hold promise as potential targets for the prevention and treatment of altitude-related neuropsychiatric disorders.
The vasogenic hypothesis
The blood–brain barrier (BBB) is a selectively permeable barrier consisting of endothelial cells, pericytes, and astrocytes, which serves to protect the brain by preventing potentially harmful substances in the blood from entering the extracellular fluid surrounding neurons (Menard et al., 2017). Hypoxia-induced oxidative stress can directly impact the stability of cell membranes, leading to lipid peroxidation and inflammation, which in turn increase membrane permeability (Bailey et al., 2006; Lafuente et al., 2016). Meanwhile, the damage to the tight junctions between endothelial cells caused by hypoxia occurs, further aggravating the permeability of the BBB (Bailey et al., 2009b; Yang et al., 2019). Particularly, hypoxia-induced upregulation of VEGF can weaken the capillary basement membrane and enhance vascular permeability (Severinghaus, 1995). Moreover, hypoxia-induced activation of the sympathetic nervous system (SNS) can lead to an increase in cardiac output.
In contrast, various factors such as prostaglandins, adenosine, and hypoxia can induce local cerebral vasodilation, leading to an increase in cerebral blood volume (Pearce, 1995). Consequently, this results in an elevation of cerebrovascular hydrostatic pressure. The buffering capacity of cerebrospinal fluid plays a crucial role in reducing the risk of cerebral edema. According to the classical tight-fit hypothesis, anatomical variations can impede the outflow of sinus veins, thereby exacerbating intracranial hypertension (Lafuente et al., 2016). The imbalance between the inflow and outflow of cerebrospinal fluid ultimately contributes to the development of vasogenic cerebral edema (Fig. 4). This hypothesis is supported by MRI studies that have demonstrated cerebral microhemorrhages (Kallenberg et al., 2008). In a retrospective study of HACE using 3.0T MRI, reversible white matter edema accompanied by microbleeds was observed (Hackett et al., 2019).

The vasogenic hypothesis of HACE. Hypoxia activates the SNS, leading to increased cardiac output and local cerebral vasodilation, as well as secretion of vasodilatory metabolites such as prostaglandins and adenosine, then causing heightened hydrostatic pressure. Hypoxia also induces enhanced oxidative stress, leading to lipid peroxidation, damage to tight junctions and increased VEGF, which in turn leads to increased BBB permeability. In addition, lower buffer capability of cerebrospinal fluid due to anatomical variations contributes to compromised outflow of cerebrospinal fluid. Ultimately, the combination of these factors leads to intracranial hypertension and brain swelling. BBB, blood–brain barrier; SNS, sympathetic nervous system.
The cytotoxic hypothesis
MRI has demonstrated the presence of cytotoxic cerebral edema in HACE, indicating dysfunction in cellular water balance (Lafuente et al., 2016). Several possible explanations for cytotoxic edema have been proposed. Insufficient ATP production caused by hypoxia can disrupt the exchange of Na+ and K+ through the Na+/K+ ATPase, leading to Na+ retention and cellular edema (Miserocchi et al., 2001). Recent research has shown that hypoxia-induced influx of Na+ depolarizes the membrane potential of neurons. Subsequently, Cl− enters the cells through a voltage-gated Cl− channel (SLC26A11) when the membrane potential drops below −20 mV, resulting in intracellular sodium and chloride accumulation and subsequent neuronal swelling (Rungta et al., 2015).
Furthermore, depolarization of the membrane activates voltage-gated Ca2+ channels, which in turn trigger the activation of calcium-dependent proteases and phospholipases. These enzymes can hydrolyze cellular components and exacerbate cell swelling, ultimately leading to cell necrosis (Michiels, 2004) (Fig. 5). It is worth noting that cytotoxic and vasogenic edema may coexist in the pathophysiology of HACE (Hackett, 1999).

The cytotoxic hypothesis of HACE. ATP deficiency caused by hypoxia inactivate the Na+/K+ ATPase, leading to Na+ retention and cell membrane depolarization. When the potential drops to −20 mV, SLC26A11 is activated and a large amount of Cl− enters the cytoplasm. The retention of Na+ and Cl− results in osmotic stress, which in turn causes cell swelling. Furthermore, as the cell membrane depolarizes, voltage-gated Ca2+ channels are also activated. The latter triggers the release of calcium-dependent proteases and phospholipases, ultimately leading to cell necrosis.
The potential mechanisms of hypoxia-related psychiatric disorders
High-altitude hypoxia leads to an increased hypoxic ventilatory response, resulting in hypocapnia and the development of PB, which in turn contributes to sleep disturbance (Weil, 2004). PB can cause alternating episodes of hypoxemia and hypocapnia, which are significant factors in the onset of AMS (Burgess et al., 2004). A meta-analysis has shown that sleep-disordered breathing is associated with cognitive impairment and executive dysfunction (Leng et al., 2017). Sleep disorders are commonly comorbid with mood disorders, with nearly all emotional disorders being linked to one or more sleep disturbances (Goldstein and Walker, 2014). Studies simulating high-altitude conditions have demonstrated that changes in sleep patterns can modulate cognitive function and mood state (de Aquino Lemos et al., 2012).
Similar findings have been observed in chronic hypoxia-related conditions such as chronic obstructive pulmonary disease (Jaffe et al., 2006; Olaithe et al., 2018). Cognitive and behavioral impairments are not only associated with sleep disturbances but also directly influenced by blood gas abnormalities (Olaithe et al., 2018). Particularly, low Pa

The underlying mechanisms for hypoxia-induced psychological disorders. At bedtime, HVR induced by hypoxia causes hypocapnia, alternating with hypoxemia, leading to PB and PSaO2, in turn intensifying the symptom of sleep disturbance. At daytime, hypoxia causes decreased Pa
Other altitude-related hypotheses
There are other hypotheses that have been proposed regarding depression and suicide associated with altitude, including changes in neurotransmitter metabolism and alterations in brain bioenergetics (Kious et al., 2018). For example, Kious et al. (2018) proposed a hypothesis indicating that when the levels of Pa
In addition, the decrease in Pa
Prevention and Intervention
Nonpharmacological approaches
Nonpharmacological interventions for preventing or managing AMS include oxygen supplementation and acclimatization (Peacock, 1998). However, the use of oxygen supplements is often limited, especially in extreme environments, and a comprehensive assessment of factors such as oxygen concentration, rate, and individual condition is necessary due to the potential for reoxygenation-related brain damage (Kim et al., 2018). Consequently, greater emphasis has been placed on adequate acclimatization before ascent, as recommended by various guidelines (Bärtsch and Swenson, 2013; Luks et al., 2019). Gradual ascent with a recommended altitude gain of 300–500 m per day above 3,000 m, along with rest days every 3–4 days, is advised to prevent AMS and HACE (Luks, 2012).
In addition, several suggestions include regular endurance training, maintaining hydration, avoiding excessive physical exertion, abstaining from substances such as alcohol and caffeine, and providing proper psychological support from companions or online resources (Rozenberg et al., 2019). In cases where HACE develops, the most crucial and effective strategy is immediate descent until symptoms subside. If immediate descent is not feasible due to limitations, the use of a portable hyperbaric chamber (Bärtsch et al., 1993) and adequate oxygen supplementation can be helpful in halting the progression of the illness (Rodway et al., 2007).
Pharmacological approaches
Acetazolamide, a carbonic anhydrase inhibitor, is widely used as a diuretic and has proven efficacy in preventing AMS (Low et al., 2012; van Patot et al., 2008). It works by enhancing ventilation and arterial oxygenation, thereby facilitating acclimatization through acidification of the blood and buffering of the respiratory alkalosis induced by high altitudes (Williamson et al., 2018). The increased oxygenation achieved with acetazolamide can reduce the production of ROS and alleviate cerebral edema (Lu et al., 2020). Common side effects of acetazolamide include paresthesia, skin rash, taste disturbances, increased urine production, nausea, and fatigue (Low et al., 2012).
In addition, individuals taking acetazolamide may experience significant reductions in diastolic, systolic, and mean blood pressure (Parati et al., 2013). Methazolamide, a lipophilic analog of acetazolamide that contains a methyl group, is also effective in preventing AMS and has the advantage of not impairing skeletal muscle function compared to acetazolamide (Teppema et al., 2020).
For individuals who experience intolerable side effects from carbonic anhydrase inhibitors, an alternative option is the use of corticosteroids. Dexamethasone is commonly used for the prevention and treatment of altitude-related illnesses (Ellsworth et al., 1987). In addition, inhaled budesonide at a dosage of 200 μg, twice daily, can be an effective alternative with fewer side effects compared to oral dexamethasone (Zheng et al., 2014). Glucocorticoids work by reducing excessive inflammation and alleviating clinical symptoms associated with altitude-related conditions (Swenson, 2016). However, considering the potential side effects associated with long-term glucocorticoid usage, such as adrenal suppression, hyperglycemia, and psychosis, their use should be guided by specific indications and limited to no more than 1 week (Bärtsch and Swenson, 2013).
Nonsteroidal anti-inflammatory drugs (NSAIDs) are commonly utilized in the field of altitude-related illnesses. Randomized controlled trials (RCTs) have demonstrated that ibuprofen (600 mg every 8 hours) effectively reduces the incidence of AMS compared to placebo (Lipman et al., 2012). However, its efficacy is slightly lower than that of acetazolamide (Burns et al., 2019). Similarly, aspirin (Burtscher et al., 2001) or acetaminophen (Kanaan et al., 2017) can also serve a prophylactic role in alleviating AMS-related headaches. Nevertheless, it is essential to consider the risks of gastrointestinal bleeding associated with the long-term use of NSAIDs.
In addition, based on redox mechanisms underlying high-altitude sickness, various food- or drug-derived antioxidants are applied to prevent these diseases. An RCT study utilizing a mixture of antioxidants, including α-lipoic acid, vitamin E, and vitamin C, demonstrated that exogenous antioxidant supplementation can alleviate the symptoms of AMS (Bailey and Davies, 2001). Another study revealed that antioxidant supplementation with N-acetylcysteine and vitamin E attenuated the adverse effects of high-altitude exposure (Vats et al., 2008). However, contrasting results were reported in another study that found no significant difference in the incidence of AMS between the antioxidant and placebo groups (Baillie et al., 2009). In addition, ginkgo extract, known for its antioxidant effects, has been considered as a potential preventive drug for high-altitude illnesses (Gaur et al., 2021).
However, a prospective double-blinded RCT study involving 614 healthy trekkers indicated that ginkgo extract only had a placebo-like effect (Gertsch et al., 2004). Finally, while Tibetan turnip (Chu et al., 2017) and Rhodiola crenulata (Chiu et al., 2013) have been suggested to potentially help prevent AMS, further validation is required.
Benzodiazepines (BZDs) are commonly prescribed as hypnotic drugs in clinical settings. They are also frequently used by hikers to manage insomnia during ascents. RCTs have demonstrated that temazepam can improve sleep quality and reduce periodic fluctuations in oxygen saturation during sleep (Dubowitz, 1998; Nickol et al., 2006). However, diazepam is not recommended due to its potential to cause hypoventilation and worsen hypoxia at high altitudes (Luks, 2008). In addition, non-BZDs, known as Z-drugs, are another type of hypnotic drug that is similar to BZDs but considered relatively safer. A meta-analysis involving 152 participants indicated that Zolpidem and Zaleplon have been effective in improving sleep quality without impacting ventilation (Kong et al., 2018). Nevertheless, even though hypnotic drugs are relatively safe, they should be used strictly according to prescriptions and instructions.
A systematic review encompassing 13 studies and involving 468 participants concluded that the evidence regarding the effects of either approach in the treatment of acute high-altitude illness is limited (Simancas-Racines et al., 2018). Moreover, due to the neurophysiological changes associated with hypoxia-related psychotic symptoms, the use of antipsychotic drugs is not recommended. Finally, we provide a summary of the findings from recent RCTs (Beaumont et al., 2007, Furian et al., 2018, Lichtblau et al., 2019, Lipman et al., 2020) investigating the drugs discussed above (Table 1).
Evidences from Clinical Randomized Controlled Trials of Common Drugs for the Prevention or Intervention of Acute Mountain Sickness and Sleep Disturbance
AMS, acute mountain sickness; BDZs, benzodiazepines; COPD, chronic obstructive pulmonary disease; NA, not available; NSAIDs, nonsteroidal anti-inflammatory drugs; Pa
Conclusion and Perspective
High-altitude hypoxia has detrimental effects on aerobic processes, leading to decreased ATP supply, disruptions in cellular metabolism, and increased production of ROS. These molecular and cellular changes can result in cerebral cell swelling or death. However, it is important to note that both hypoxia and reoxygenation can contribute to brain damage, similar to the process observed in ischemia and reperfusion (Torres-Cuevas et al., 2019), suggesting a potential risk of secondary damage when supplementing oxygen at high altitudes. On the contrary, the antioxidant system plays a crucial role in counteracting ROS and maintaining the delicate balance of the redox system. The GSH antioxidant system, in particular, plays a vital role in protecting cells against oxidative stress (Doll et al., 2019).
As mentioned earlier, various antioxidant drugs and foods help mitigate oxidative stress. However, due to individual medical conditions and the complex natural environment at high altitudes, personalized approaches should be considered rather than adopting a “one size fits all” approach to address altitude mountain sickness (Yang et al., 2022). Genomic studies have identified the genetic basis of high-altitude adaptations (Bigham and Lee, 2014; Simonson et al., 2012; Simonson et al., 2010), with a significant number of genes involved in the HIF pathway (Bigham, 2016). Further exploration of the genetic mechanisms underlying high-altitude adaptation will enhance our understanding of the molecular foundations involved (Bigham, 2016). With advancements in identifying key genes, it is hoped that future detection of risk genes will enable the prediction of susceptibility to high-altitude sickness.
Interestingly, studies have found that altitude is associated with lower rates of certain psychiatric disorders, such as attention-deficit/hyperactivity disorder (Huber et al., 2018). Moreover, a recent study reported a decrease in the mortality rate of Alzheimer's dementia with increasing altitude (Thielke et al., 2015). These findings suggest that altitude might act as a protective factor against the occurrence of psychosis. However, it is important to note that the altitudes examined in these studies were no more than 2,000 m. Therefore, controlled exposure to hypoxia may potentially serve as a treatment strategy for certain neuropsychiatric illnesses (Burtscher et al., 2021).
Although the high-altitude environment is generally considered a form of chronic hypoxia, it is worth noting that the mechanisms underlying the acute and chronic changes in neuropsychiatric function associated with high-altitude hypoxia are different. In the acute period, the neuropsychiatric symptoms mentioned earlier may arise directly from inadequate oxygen supply to the brain cells. For example, the presence of a low Pa
In contrast, the complex neuropsychiatric syndrome observed can be attributed to the detrimental effects of prolonged altitude hypoxia on neural structure and function. For example, as the duration of hypoxic exposure increased, there was a tendency for atrophy in the gray matter throughout the entire brain, indicating nonspecific injury characteristics (Zhang et al., 2010). The impact of long-term high-altitude hypoxia on cognitive function stems from alterations in multiple physiological mechanisms, encompassing oxidative stress, polycythemia, and heightened levels of HIF-1α (Li and Wang, 2022).
It is important to acknowledge that many studies investigating the effects of hypoxia have been conducted in simulated hypobaric hypoxic chambers. The generalizability of these findings to real-life high-altitude conditions remains a topic of debate. Furthermore, there are substantial variations among studies, including differences in participant characteristics, altitude levels, duration of exposure, specific tasks, and measurement methods (Bahrke and Shukitt-Hale, 1993). In addition, in actual high-altitude environments, climbers face not only oxygen insufficiency but also various other factors such as cold weather, lack of energy, and fatigue (Brugger et al., 1999). However, several challenges still need to be addressed, including the limited availability of large clinical samples, the harsh natural environment, and sufficient funding. Consequently, there is an urgent need for large-scale RCTs investigating the cerebral effects of hypoxia conducted directly in high-altitude areas.
Footnotes
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
This work was supported by grants from the 1.3.5 project for disciplines of excellence, West China Hospital, Sichuan University (ZYJC21004).