
Abstract
Sharma, Narendra Kumar, Mansi Srivastava, Tikam Chand Dakal, Vipin Ranga, and Pawan Kumar Maurya. Acute hypobaric hypoxia causes alterations in acetylcholine-mediated signaling through varying expression of muscarinic receptors in the prefrontal cortex and cerebellum of rats’ brain. High Alt Med Biol. 26:156–164, 2025.
Background:
Muscarinic receptor (CHRM) proteins are G-protein-associated acetylcholine receptors found in neuronal membranes. Five major subtypes, CHRM1–CHRM5, modulate acetylcholine in central nervous system signaling cascades. CHRM1, CHRM3, and CHRM5 are linked to Gαq/Gα11 proteins, whereas CHRM2 and CHRM4 are linked to Gαi/Gαo proteins.
Objective:
Limited research has been conducted to explore the impact of HH on CHRM gene expressions. It is caused by low oxygen availability at high altitudes, which impairs neurotransmission, cognitive performance, and physiological functions. Previous studies have shown that exposure to hypoxia leads to a reduction in CHRM receptors, which in turn causes alteration in signal transduction, physiological responses, cognitive deficits, and mood alterations.
Method:
In the present study, we have used semiquantitative PCR to measure muscarinic receptor gene expression after 6, 12, and 24 hours of HH exposure at 25,000 feet using a decompression chamber in rat brain’s PFC and cerebellum.
Result:
We have found that CHRM1–CHRM5 downregulated after acute exposure to hypoxia until 12 hours, and then, the expression level of these receptors increased to 24 hours when compared with 12 hours in PFC. All subtypes have shown a similar pattern in PFC regions under hypoxia exposure. On the other hand, these receptors have shown altered expression at different time points in the cerebellum. CHRM1 and CHRM4 acutely downregulated, CHRM2 and CHRM5 downregulated, while CHRM3 upregulated after hypoxia exposure.
Conclusion:
Our study, for the first time, has shown the altered expressions of muscarinic receptors under temporal hypoxia exposure. The altered expression pattern has shown an association with acclimatization and protection against necrosis due to hypoxia. This study may pave further investigations for understanding and addressing the cognitive, behavioral, and physiological impacts of hypoxia and therapeutic development.
Introduction
During high-altitude flights and military operations, conditions of low atmospheric pressure of oxygen arise with consummate disruption of homeostasis consequently resulting in a state commonly referred to as ‘hypobaric hypoxia’ (HH) (Sharma et al., 2022; Sharma and Arya, 2022; Utrera et al., 2022). In response to an abrupt alteration in oxygen demand, the human body undergoes physiological adjustments to acclimatize. These adjustments include hyperventilation, tachycardia, and heightened sympathetic activity. The cardiovascular and respiratory systems of the human body undergo excessive strain in order to rectify the perfusion mismatch (Lopez-Barneo et al., 2016). HH-related stimulation of reactive oxygen species (ROS) has been reported due to an imbalance of oxygen concentration at the cellular level, affecting metabolism (Sharma et al., 2012; Sharma et al., 2013; Sethy et al., 2014; Gaur et al., 2021). By gaining a deeper understanding of the implications of HH, we can potentially develop more effective strategies for managing the challenges associated with high-altitude environments and optimizing human performance and health in such conditions.
The synthesis of acetylcholine is facilitated by cholinergic neurons that possess the enzyme choline acetyltransferase. The cholinergic deficit is regulated by two types of receptors: nicotinic receptors, known for their rapid and excitatory properties, and muscarinic receptors, which have slower kinetics and may have inhibitory or excitatory effects (Wallace and Bertrand, 2013). The muscarinic acetylcholine receptors (mAChRs) may be classified as G-type protein-coupled receptors, characterized by their possession of seven hydrophobic transmembrane domains. The location of mAChRs inside the brain exhibits heterogeneity, although they are ubiquitously present. The primary subtypes are M1, M2, and M4, but the expression levels of M3 and M5 are much lower. M1, M3, and M5 receptors exhibit a phylogenetic resemblance and play a role in the activation of phospholipases C, A2, and D, as well as the activation of tyrosine kinase. This activation leads to the following influx of calcium ions and is often seen in postsynaptic regions. Both M2 and M4 exhibit comparable behavior by inhibiting adenylate cyclase, resulting in a decrease in intracellular levels of cAMP. Muscarinic M1–M5 receptors have been shown to control the metabotropic actions of acetylcholine in the brain (Eglen, 2006). The autoinhibition of ACh release requires different mAChRs that are located throughout the brain with varied localization (Zhang et al., 2002; Wong et al., 2023). It has been previously established that muscarinic receptors are novel to the physiology of the brain and have been a contributing factor in various developmental disorders (Volpicelli and Levey, 2004). Nevertheless, the impact of hypoxia on muscarinic receptors and their expression in the cellular membrane remains to be fully understood.
Acetylcholine is a neuromodulator that alters the excitability of neurons, influences synaptic transmissions, modulates synaptic plasticity, and induces neuronal firing in groups (Wonnacott, 1997; Rice and Cragg, 2004; Changeux, 2010; Picciotto et al., 2012). The coordination abilities of acetylcholine are the regulators of various complex behavior due to its involvement in the stimulation of all major brain areas (Picciotto et al., 2012). A change in expression of muscarinic receptors directly affects acetylcholine-mediated signaling affecting the overall physiology and cognition drastically.
The current investigation aims to examine the impact of HH on the expression of muscarinic genes in mouse brain tissue, specifically focusing on the prefrontal cortex (PFC) and cerebellum. The PFC is essential for higher order cognitive functions such as decision-making, attention, and working memory (Friedman and Robbins, 2022); while the cerebellum is traditionally associated with motor control and coordination, it also plays a role in cognitive functions (Clark et al., 2021). The cerebellum contains a high density of Purkinje cells that are sensitive to hypoxia and can reveal how motor coordination is affected under low oxygen conditions (Kaur et al., 2005); on the other hand, basal ganglia are involved in the moment regulation (Lanciego et al., 2012). The study of the effect of muscarinic receptors in PFC and cerebellum can provide insights into the mechanisms underlying motor and cognitive impairments seen in conditions like stroke, traumatic brain injury, and chronic obstructive pulmonary disease (Jones and Harvey, 2023) and lead to the development of targeted therapies. These receptors contribute to cell survival; reduce oxidative stress (Intachai et al., 2022), synaptic plasticity, neurotransmitters (Yang et al., 2021), behavior and psychological states (anxiety, depression, and impaired motor coordination); and mitigate inflammation under hypoxic conditions (Okan et al., 2021). The effects of muscarinic receptors on the brain’s PFC and cerebellum under hypoxia are critical for understanding and addressing the cognitive, behavioral, and physiological impacts of reduced oxygen supply. Studying the interplay between muscarinic receptors, hypoxia, and brain regions such as the PFC and cerebellum can offer insights into other fields including vascular biology, respiratory medicine, and even sports science, where hypoxia is a common concern. This research has the potential to inform clinical practices and improve outcomes for individuals experiencing hypoxia-related conditions. The study used semiquantitative PCR as a method to assess the expression levels of muscarinic genes in the PFC and cerebellum at four time points: 0, 6, 12, and 24 hours of HH exposure. Subsequently, the PCR results were subjected to densitometry, and the resulting data underwent statistical analysis. Additional computational investigations were conducted to get insight into the involvement of additional significant factors in the alterations in gene expression generated by hypoxia.
Material and Methods
Animal model
We have used male Sprague-Dawley (SD) rats (n = 24) weighing 220 ± 10 g in the present study. The Institutional Animal Ethics Committee approved all animal procedures and the experimental methodology. All attempts were made to reduce the number of animals used and minimize animal suffering. The experiments were carried out according to established procedures.
Hypoxia exposure
We randomly divided the animals into four groups for the study, hence six rats per group were assigned. Rats from group I were kept in normal conditions and termed as control or normoxia (0 hour). Groups II, III, and IV were exposed to HH for 6, 12, and 24 hours, respectively. The animals were exposed to hypoxia using a decompression chamber (Seven Star, India) for an altitude equivalent to 25,000 feet (282 mmHg). Temperature and humidity were kept at 28 ± 2°C and 60 ± 5%, respectively, and were precisely controlled during exposure. The rate of rise to height was kept at 300 meters per minute, and it took 20–25 minutes to reach the target altitude.
Sample preparation
After the exposure to hypoxia, all rats including control (0 hour) and hypoxia-exposed rats (6, 12, and 24 hours) were deeply anesthetized with 100 mg/kg, i.p., sodium pentobarbital, transcardially perfused with chilled PBS, and decapitated. The PFC and cerebellum were carefully dissected out from the rat brain tissue for further experiments. In brief, the rat was quickly decapitated using sharp scissors, and immediately proceeded to remove the skull to extract the brain. Then, made an incision along the midline of the scalp, peeled back the skin, and removed the skull cap using scissors. The brain was gently lifted out from the skull using a spatula by cutting cranial nerves and blood vessels holding the brain in place. After removal, the intact brain was placed in a cold saline solution to reduce degradation. The brain was kept on ice, and the PFC regions, located in the anterior part of the brain, were dissected out by the corneal section. The cerebellum located at the posterior part and bottom of the brain was separated from the brainstem using a fine blade.
Total RNA isolation from the PFC and cerebellum
For total RNA isolation, 100 mg (wet weight) of the tissue was weighed and then macerated in liquid nitrogen using a mortar and pestle. The powdered samples of each tissue were weighed 30 mg, 1 ml of Trizol was added to it, and RNA was isolated using RNeasy Mini Kit (Qiagen Inc. Valencia, USA). The quality and quantity of RNA were measured by nanodrop and samples having A260/280 ratio between 1.8 and 2.0 were taken for further proceedings.
cDNA synthesis from the RNA isolated from PFC and cerebellum
To study the expression profile of muscarinic receptors, isolated RNA was reverse transcribed using the Enhanced AMV Reverse Transcriptase Enzyme System (Sigma, St Louis, MO). In brief, 1 μg of RNA was taken per sample, and cDNA synthesis was done by adding 500 μM dNTPs, 2.5 μM random nonamers and reaction buffer, RNase inhibitor, and enzyme for the final volume of reaction to 10 µl. Then sample tubes were incubated at 42°C for 50 minutes. The cDNA solution was then kept at 20°C before being utilized as a template for PCR amplification.
Semiquantitative RT-PCR of M1–M5 genes in PFC and cerebellum
Semiquantitative RT-PCR analysis of CHRM1–CHRM5 was executed to validate the differential expression of the muscarinic receptors in PFC and cerebellum. The sequence of the primers of receptors was designed using the Primer Premier 5.0 Software (Table 1). To test their specificity, SequencherTM software was employed to avoid the amplification of any other related gene members. The previous report indicated that exposure to hypoxia altered the expression of GAPDH (Sharma et al., 2013), while, 18S rRNA has stable expression, low variability, and is less affected by experimental conditions. Hence for normalization, 18s rRNA (ubiquitous housekeeping gene) was used as a loading control. TaKaRa Taq was used to carry out PCRs on all the samples. Multiplex PCRs were done to optimize the temperature of primers, and varied cycles were run for all subtypes.
Details of the Muscarinic Receptor Genes Studied in This Research Elucidating the Gene Name, Accession Number, Primer Labeling Name, Primer Sequence in 5′ to 3′ Direction, Number of Bases in the Primer Sequence and the Size of the Gene for Six mRNAs Studied as Annotated in nBLAST (M1, M2, M3, M4, M5, and 18sRNA-Control)
Agarose gel electrophoresis of the PCR products
Electrophoretic separation on a 1.2% agarose gel was employed to study the gene amplification of the PCR products. Ethidium bromide was used as a staining agent to ensure the visualization of DNA on the gel. To study the expression, densitometric analysis of the PCR products was done using Image J software and represented as graphs.
Brain bulk tissue gene expression analysis
The bulk tissue gene expression profiling was performed using GTEx Analysis Release V8 (dbGaP Accession phs000424.v8.p2) (https://www.gtexportal.org/). The Genotype-Tissue Expression (GTEx) initiative is building a public archive for studying tissue-specific gene expression and regulation. A population of 1000 persons was sampled from 54 disease-free tissue sites. The main reason for collecting these samples was to do molecular assays like whole genome sequencing (WGS), whole exome sequencing (WES), and RNA-Sequencing (RNA-Seq). Additional samples are available from GTEx Biobank. Gene expression, quantitative trait loci (QTLs), and histology images are freely available on the GTEx portal.
Statistical analysis
A minimum of three replicates (n = 3) for each experiment were taken to ensure reproducibility. The results of semiquantitative RT-PCR were analyzed by one-way ANOVA, followed by the Tukey multiple comparison test. All the graphs were prepared using GraphPad Prism 8.
Results
Expression analysis of muscarinic receptor genes in the PFC
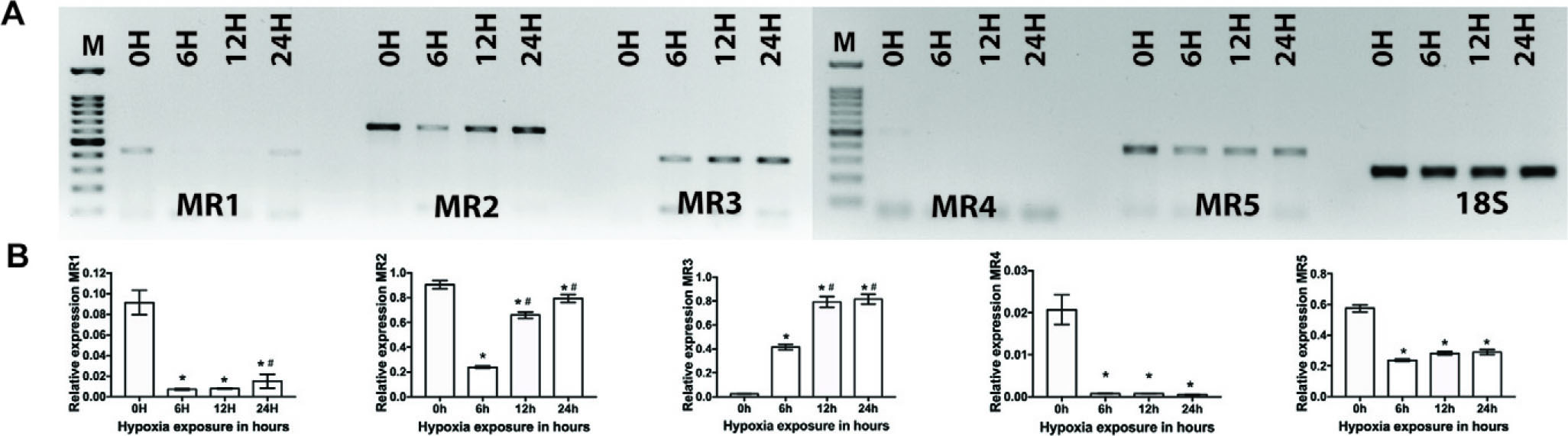
The expression pattern of muscarinic receptor genes (Chrm1, Chrm2, Chrm3, Chrm4, and Chrm5 also known as MR1, MR2, MR3, MR4, and MR5, respectively) in the PFC was determined using semiquantitative PCR. This analysis was conducted at four time points (0, 6, 12, and 24 hours) after exposure to HH. The analysis of the expression profile of MR1–MR5, in conjunction with 18sRNA, has been conducted in the PFC (Fig. 1).

Expression profiling of M1–M5 genes using semiquantitative RT-PCR from prefrontal cortex tissue after hypobaric hypoxia (0, 6, 12, and 24 hours, respectively). Panel
The results of MR1 indicated a decrease in the levels of expression in the samples collected at the 12-hour exposure. Subsequently, in the 24-hour group, a restoration of expression levels was found. In the MR2 gene, there was a decrease in gene expression seen from 6 to 12 hours of exposure. Subsequently, in the 24-hour samples, there was an observed increase in gene expression. Likewise, a similar pattern of downregulation up to a duration of 12 hours followed by a subsequent rebound beyond 24 hours was found in MR3, MR4, and MR5. In the PFC, it was seen that MR4 and MR5 were present in minimal quantities relative to MR1, MR2, and MR3.
Expression analysis of muscarinic receptor genes in the cerebellum
In a similar vein, the semiquantitative PCR technique was used to examine the expression patterns of the muscarinic receptor genes (Chrm1, Chrm2, Chrm3, Chrm4, and Chrm5), in the cerebellum of rats subjected to HH for durations of 0, 6, 12, and 24 hours.
The densitometry data of the cerebellum revealed that the expression levels of all receptors were comparatively lower than those seen in the PFC (Fig. 2). The MR1 gene was detected in minimal quantities in the control group at the start of the experiment (0 hour). Subsequently, its presence became insignificant after exposure to hypoxia. However, a little increase in expression levels was noted in the samples collected after 24 hours, indicating a limited recovery. The MR2 gene exhibited a notable decrease in expression levels within the 6-hour exposure to hypoxia, followed by a subsequent recovery in expression levels in the 12- and 24-hour exposures. In the control (0 hour) sample, the expression of MR3 in the cerebellum was almost negligible. However, after 6 hours of exposure to hypoxia, there was a notable deviation from the overall pattern, with a four-fold increase in expression levels. This increase continued to rise, reaching eight fold in the 12- and 24-hour exposure to hypoxia. MR4 was detected in minimal quantities inside the cerebellum and exhibited significant downregulation, with no subsequent restoration seen in its expression levels. The MR5 gene exhibited a decrease in expression in the 6-hour sample, followed by an elevation in expression levels at the 12-hour hypoxia exposure.
Expression profiling of M1–M5 genes using semiquantitative RT-PCR from cerebellum tissue after hypobaric hypoxia (0, 6, 12, and 24 hours, respectively). Panel
Bulk tissue gene expression profiling of five muscarinic receptors in humans
The bulk tissue gene expression profiling was performed using GTEx Analysis Release V8 (dbGaP Accession phs000424.v8.p2) (https://www.gtexportal.org/). The GTEx project is an ongoing endeavor aimed at constructing a comprehensive publicly accessible repository for investigating the expression and regulation of genes unique to various tissues. A total of 54 tissue locations that were not affected by any diseases were selected for sample collection, including a population of around 1000 people. The primary purpose of collecting these samples was to conduct molecular tests, namely, WGS, WES, and RNA-seq. Additional samples may be obtained via the GTEx Biobank. The GTEx portal offers unrestricted access to a range of data, including gene expression, QTLs, and histological pictures.
We found that CHRM1 is highly expressed in most regions of the brain, including amygdala, basal ganglia, BA24, cortex, frontal cortex, hippocampus, nucleus accumbens, and putamens. CHRM2 showed no expression in the brain’s major regions. CHRM3′s expression was limited to BA24, cortex, and frontal cortex, while CHRM4’s expression was seen in BA24, cortex, frontal cortex, nucleus accumbens, and putamen. CHRM5 displayed expression in the hippocampus, hypothalamus, spinal cord, and substantia nigra. The expression profiles of all muscarinic receptor proteins were found to be highly correlated with its expression in the rodent model suggesting that findings in the rodent model could reflect the predictive outcomes in any therapeutic interventions as well (Fig. 3).
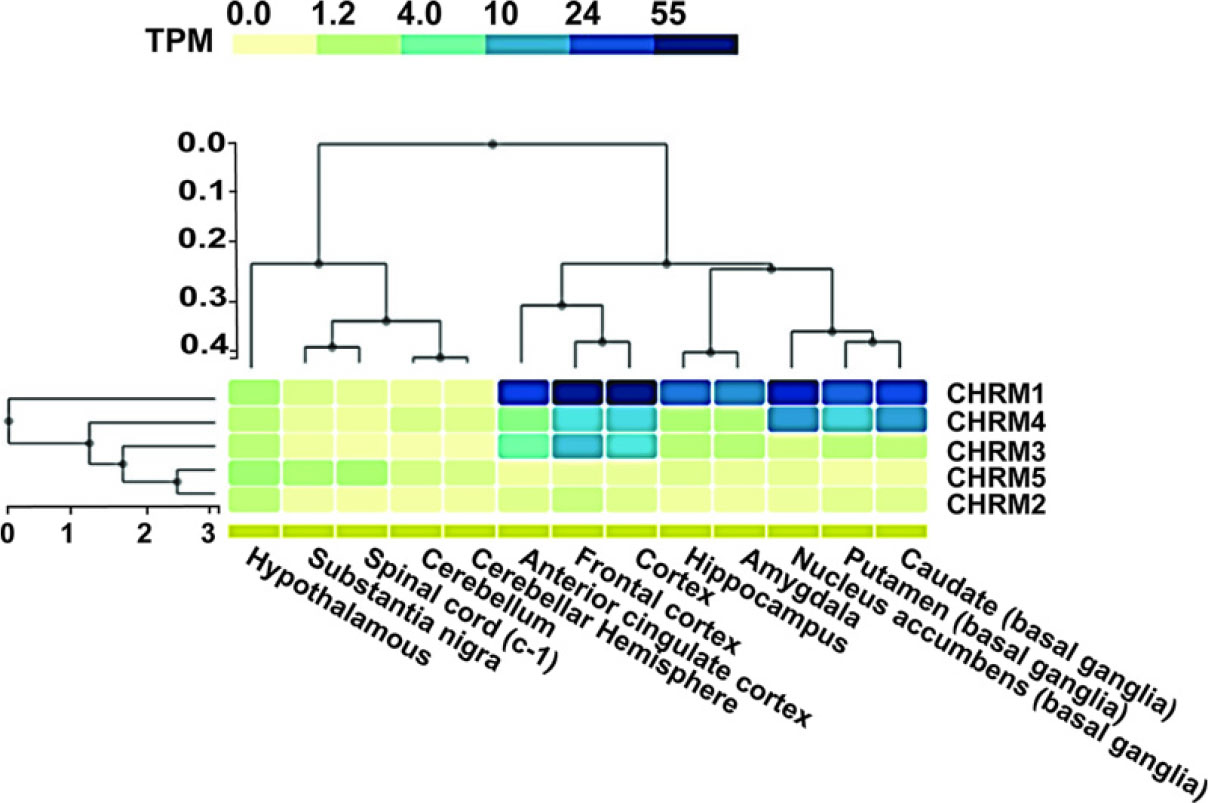
Heat map of the bulk tissue gene expression of CHRMs in different regions of brain.
Discussion
The gene expression data pertaining to all five subtypes of muscarinic receptors in this research exhibits a discernible pattern in their expression. In both the PFC and the cerebellum, a consistent pattern of decrease was seen in all subtypes, with the exception of subtype M3 in the cerebellum. This drop was evident during the initial hours of hypoxia exposure hours, but later on, the expression of these subtypes rebounded, which shows the role in adaptation to hypoxia. Exposure to hypoxia is afterward accompanied by two distinctive circumstances that impact genetic expression and protein synthesis inside the brain (Rybnikova and Samoilov, 2015). During the early stages after hypoxia, the phenomenon of tolerance induction takes place, facilitating the production and translation of immediate protective proteins. Following the occurrence of tolerance expression, the succeeding stage entails more gene expression and protein synthesis to fulfill the regular genomic needs, hence granting neurons enduring resilience against further hypoxic stress (Burtscher et al., 2021). Muscarinic receptors are considered proteins associated with endurance rather than quick responses. In the cerebellum, M3 expression, initially minimal in control mice, significantly increases with hypoxia exposure. M3 has a protective effect against necrosis by stabilizing the caspase-8 cascade. This may explain its increased expression in hypoxic settings, therefore classifying it as a constituent of the rapid response group in hypoxia conditioning (Huang et al., 2020). With the focus on physiological hallmarks of hypoxia, an association was found between the symptoms of hypoxia and mechanisms affected due to dysfunction related to muscarinic genes. Cognition and memory are the two main functions of the primary M1 muscarinic receptor. It is one of the first agonist-based targets being used for therapeutics (Verma et al., 2018). M1 receptor signaling blockage has been found to impair attention (Nicklas et al., 2021). It has been reported to have a significant impact on cognitive functioning and has also been associated with schizophrenia at low expression levels (Dean and Scarr, 2016). During peripheral inflammation, a steep decline in the M1 receptors in the PFC was observed. M1 and M4 receptors are also reported to reduce the positive, negative, severity, and cognitive symptoms of schizophrenia (Dean et al., 2023). Recent findings strongly suggest the possible advantages of activating important muscarinic receptors for the improvement of general cognition and the treatment of psychotic symptoms (Jones and Harvey, 2023). In the conducted research, it was determined that the variable under investigation was significantly influenced by exposure to hypoxia. The drop in M1 receptor expression levels may contribute to the reported short-term cognitive deficits associated with HH (Biswal et al., 2016).
The modulation of cardiovascular systems in the cerebellum involves the participation of M2 receptors. The activation of the receptor elicits substantial depressor and bradycardic responses within the cardiac system (Zhang et al., 2016). Recent research demonstrates, for the first time, that the presence of muscarinic receptors is crucial for mitigating spatial memory retrieval deficits resulting from traumatic brain injury, particularly in males, indicating sexual dimorphism in their ability to alleviate retrograde amnesia (Rashid and Ahmed, 2023). Under hypoxic stress, the decline in the M2 receptors could be a potential contributor to tachycardia, which also works as a compensatory response to restore oxygen levels by increasing the blood flow rate. The M3 receptor-mediated reaction with microglia has been shown to be of significance for positive outcomes following an ischemic stroke (Costa et al., 2021). One of the critical events in a brain ischemic stroke is the development of hypoxia (Sarkar et al., 2019). Hypoxia can trigger regulatory cascades contributing to stroke symptoms. While hypoxia induces downregulation of M3 receptors in the PFC, an atypical overexpression is observed in the cerebellum. Previous research has shown a correlation between the upregulation of M3 and the hypoxia involving various physiological and pathophysiological mechanisms. Hypoxia triggers adaptive responses that include upregulation of the M3 receptor to modulate smooth muscle activity and maintain tissue functions under low oxygen conditions, which is evident in conditions like bladder ischemia (Chess-Williams and Sellers, 2023) and pulmonary hypertension (Pullamsetti et al., 2020). In certain tissues, increased M3 expression could potentially enhance vasodilation or improve blood flow to hypoxic areas, thus aiding in oxygen delivery.
M2 and M3 receptors have been reported as major players in the pathology of bipolar and major depressive disorders (Gibbons et al., 2009). The existing correlation between pre- and perinatal hypoxia and bipolar disorder is being explored on a molecular level in which we propose that the muscarinic receptor might have a role to play (Haukvik et al., 2014).
Respiration modulation is the process of regulating breathing rate with the involvement of the M4 receptor (Lima et al., 2019). Our results have shown the alterations in M4 expression under HH. In hypoxic conditions, active expiration takes place, while the respiratory process is modulated by M4 receptor-based signaling through the cholinergic pathway and inhibitory protocols (Silva et al., 2019). Similarly, in a recent study, M4 and M5 receptors were used as potential therapeutic targets for the treatment of alcohol use disorders (Walker et al., 2023).
The muscarinic receptor subtype M4 is prominently implicated in neurodegenerative disorders such as Parkinson’s and Alzheimer’s, primarily through M4-mediated cholinergic transmission associated with motor dysfunctions in Parkinson’s disease (Wang et al., 2016). M5 receptors play a key role in modulating behavior reward-based activities of rats (Forster et al., 2002). Similar to other receptor subtypes, M5 has correlations with neurodegenerative disorders as well, and allosteric sites of M5 receptors were being explored as putative drug targets (Bock et al., 2018). The augmentation of M5 receptors’ activation has been identified as a potentially valuable therapeutic objective for mitigating the manifestations and indications of anxiety and depression (Razidlo et al., 2022). The M5 receptors have emerged as a promising and innovative therapeutic target for addressing several addiction illnesses, such as opioid use disorder. Neurodegenerative illnesses often occur subsequently due to hypoxia, suggesting a potential causal relationship between the two. Changes in the expression of muscarinic receptors impede physiological processes, leading to negative consequences.
During periods of hypoxia exposure, the brain undergoes oxidative stress, resulting in the activation of hypoxia-inducible factor and the formation of ROS. The downregulation of muscarinic receptors is a result of processes such as lipid peroxidation, caspase-mediated proteolysis, and DNA damage. The consequence of this phenomenon is a general decrease in the production of acetylcholine, which subsequently triggers additional cell death and disruption of metabolic processes. These effects contribute to the manifestation of hypoxia-related symptoms, including memory impairment, cognitive impairments, and various complications in the functioning of the heart, lungs, and digestive system. These physiological processes are governed by the signaling pathways, as discussed earlier, mediated by muscarinic receptors, and illustrated in Fig. 4, highlighting their central role in regulating the effects of hypoxia exposure.

The cascades have been enlisted that get activated when the organism is exposed to hypoxia. Muscarinic receptor genes are correlated with guanine nucleotide-binding proteins and acetylcholine nicotinic receptors (STRING DATABASE). Downregulation of muscarinic receptors thus leads to faulty functioning of the associated proteins resulting in cognitive dysfunction, ischemic stroke, respiratory issues, and behavioral alterations in the organism.
Limitations
Multi-gene Transcript Expression of all five human CHRMs in the brain was assessed using the GTEx portal. We also assessed the expression of all CHRMS in different regions of the rat brain using experimental in vivo conditions and found congruent results. Lastly, we address our findings in light of the analyses’ limitations as biological interpretation and implications of differential gene expression of all five human CHRMs from GTEx, although largely found congruent, could not be correlated with the gene expression of all rat CHRMs. However, it remains a limitation whenever one attempts to determine and interpret the differences in gene expression profiles between rats and its counterpart humans as the study could not be performed in the absence of human samples.
Additionally, we have not monitored the formation of CO2 and ammonium in hypoxia chamber. As a result, the toxic impact of the CO2 and ammonium and its contribution to hypoxic stress couldn’t be measured as well. This has been one of the limitations of the current study.
Conclusion
The alteration in the amounts of muscarinic receptors seen in the PFC and cerebellum signifies the involvement of oxidative stress in the decrease of receptor expression. The onset of hypoxic circumstances has been seen to impact brain tissue after around 6–12 hours. This subsequently results in a decrease in the numbers of mAChRs due to the disintegration and depletion caused by ROS. Nevertheless, the human body’s capacity to adjust prevents further depletion and develops a tolerance to reduced oxygen levels, resulting in the restoration of expression levels within a span of 12 hours. The study aimed to investigate the relationship between symptoms of HH and gene expression. The aberrant functioning of the M3 receptor elucidates variations in functionality across its subtypes. Additional investigation into the biochemical characteristics of the receptors may provide corroborating information substantiating the involvement of the muscarinic receptor in the manifestation of hypoxic symptoms.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Life Science Research Board (LSRB), Defence Research and Development organization (DRDO), Government of India, India, grant reference: LSRB/01/1500 1/LSRB- 407/PEE&BS/2023.