
Abstract
Zhang, Wenxin, Yuting Yang, Jing Guo, Fei Hu, Yuan Ma, and Qing Ouyang. Phenothiazine confers neuroprotection via Dpp2/7 in high altitude traumatic brain injury mouse model. High Alt Med Biol. 26:256–264, 2025.
Background:
Traumatic brain injury (TBI) in high altitude areas can lead to more severe cerebral edema, higher disability, and mortality than in low altitude areas. This study was designed to evaluate the neuroprotective effects and underlying mechanisms of phenothiazine on TBI at high altitudes.
Methods:
Mice were kept in a hypobaric chamber for 7 days under simulated conditions of 5,000 m above sea level. A controlled cortical impact (CCI) model was established and followed by phenothiazine (chlorpromazine and promethazine) and Dpp2/7 inhibitor UAMC00039 treatment. Hematoxylin–eosin (HE) staining, immunohistochemistry (IHC), western blot, label-free quantitative proteomics, and real-time quantitative polymerase chain reaction (RT-qPCR) assays were performed to assess the effects of phenothiazine and UAMC00039 on TBI.
Results:
HE staining confirmed that phenothiazine treatment could ameliorate CCI-induced brain injury. IHC, western blot, and RT-qPCR showed that cell apoptosis was alleviated by phenothiazine after high altitude TBI, as proved by the reduction of cleaved-Caspase-3 and increased Bcl-2 expression. Label-free quantitative proteomics, IHC, and western blot showed that phenothiazine significantly upregulated Dpp2/7 after high altitude TBI. Western blot and IHC showed that UAMC00039 treatment significantly reversed phenothiazine-mediated Bcl-2 upregulation and cleaved-Caspase-3 downregulation after high altitude TBI.
Conclusions:
The results indicated that phenothiazine offers neuroprotective effects via antiapoptosis after high altitude TBI, and this protective mechanism is associated with Dpp2/7-mediated Bcl-2 expression and Caspase-3 cleaving.
Introduction
Traumatic brain injury (TBI) is one of the main causes of disability and mortality, which brings physical, mental, and economic burdens (Maas et al., 2022). High altitude TBI occurs in areas above 3,000 m above sea level (Yang et al., 2022). Previous study indicates that the occurrence of accidental injuries at high altitude has risen rapidly by 12.7% with the development of transportation (Wang et al., 2019). It is indisputable that high altitude TBI leads to more severe mortality and disability than that in low altitude areas. Hence, it is imperative to explore therapeutic methods to minimize mortality and disability after high altitude TBI, and neuroprotective strategies to maximize neurofunctional outcomes of patients with high altitude TBI.
Hypobaric hypoxia is the main feature in high altitude areas, which causes a different degree of damage to the central nervous system, circulation, and respiratory systems. Compared with low altitude areas, TBI following abrupt exposure to high altitude areas is more complex. At high altitude areas, the special environment of hypobaric hypoxia aggravates secondary brain damage, which induces blood–brain barrier (BBB) leakage, resulting in more severe cerebral edema after TBI, and high altitude TBI is also followed by more remarkable cell necrosis, apoptosis, and neuroinflammation, all of which result in more severe clinical outcomes than in low altitude areas (Liu et al., 2023; Wei et al., 2020).
Pathological mechanisms of TBI are classified into primary and secondary injury. Recovery from the destruction of brain tissue caused by primary injury is not possible; however, secondary injury is a dynamic process after primary injury, which is the optimal opportunity for clinical intervention. Phenothiazine is an organic compound that readily crosses the BBB. Dual therapy with chlorpromazine and promethazine, two phenothiazine drugs, has been found to have a significant protective effect on neurological tissue in both transient and permanent ischemic strokes (Geng et al., 2017). Previous studies suggest that the neuroprotective mechanisms of chlorpromazine and promethazine are only partially dependent on the hypothermia induced by artificial hibernation; besides that, scavenging free radicals, inducing antioxidant protein synthesis, targeting mitochondrial permeability transmission, and reducing apoptotic injury also play important roles (Guo et al., 2022; Guo et al., 2021a; Tong et al., 2019).
Dipeptidyl peptidases (Dpps) are zinc-dependent hydrolases involved in cleaving dipeptides of various peptide substrates, which are composed of Dpp2/7, Dpp3, Dpp4, Dpp6, Dpp8, Dpp9, and Dpp10 (Choy et al., 2021). Dpp2/7 (also known as Dpp II and quiescent cell proline dipeptidase) is localized to intracellular vesicles rather than lysozymes, which can be secreted in active form in response to calcium release. It has been confirmed that Dpp2/7 is critical for the G0 survival program of lymphocytes and neuronal cells, and inhibition of Dpp2/7 induces apoptosis of these quiescent cells (Danilova et al., 2007). Recent studies show that Dpp2/7 is expressed significantly higher in colorectal cancer (CRC) than in non-tumor tissues, and increased Dpp2/7 is significantly associated with greater malignant behavior and shorter overall survival (Shang et al., 2024; Zhang et al., 2023). The downregulation of Dpp2/7 confers hepatocellular carcinoma (HCC) cells a lower activity of proliferation and G1 cell cycle arrest and increases cell apoptosis through the upregulation of Bax/Bcl-2 ratio (Hu et al., 2019). Thus, Dpp2/7 is an integral part of the machinery maintaining cell quiescence, which is involved in cell apoptotic regulation.
In the present study, we aim to investigate the neuroprotection effects of phenothiazine on high altitude TBI mice. Our study also sought to delineate the underlying mechanisms by which phenothiazine induces neuroprotection effects. A robust understanding of the signal pathway of neuroprotection effects conferred by phenothiazine after high altitude TBI would guide future endeavors in finding therapeutic targets to decrease the disability and mortality of patients.
Materials and Methods
Animal preparation
Healthy C57BL/6 mice (male, weighing 18–22 g, and aged 6–8 weeks) were purchased from GemPharmatech LLC. The mice were well fed and housed in a pathogen-free and climate-controlled environment at room temperature (22 ± 1°C), humidity (50%–60%), and under a 12-hour/daylight cycle. Experiments were approved by the Ethics Committee of the General Hospital of Western Theater Command in Chengdu and performed in line with the guidelines for the care and use of experimental animals of the National Institutes of Health.
High altitude TBI mice construction
The mice were placed into the temperature-controlled hypobaric chamber 7 days before the TBI operation. The oxygen content (12.95%) and oxygen partial pressure (11.33 kPa) were selected to simulate the 5,000 m high altitude environments. After 7 days of high altitude exposure, the mice were removed from the chamber and anesthetized with 2.5% tribromoethanol (2.5 mg/kg). High altitude TBI was induced using the controlled cortical impact (CCI) procedure. The mice were fixed on the brain stereotactic instrument, a 3 mm diameter bone window was opened on the right parietal bone, and the center of the bone window was located 1 mm lateral to the midline and 1.3 mm posterior coronal suture. A 2 mm flat-tip impounder was dropped onto the mice dura mater with a contact time of 75 ms, an impact strength of 2 kg, and an impact depth of 2 mm by using an electronic-driven CCI device. Immediately after CCI, the mice’s scalps were sutured and sent back to the decompression chamber and then returned to normal feeding.
Experimental protocol
Twenty high altitude exposure mice underwent CCI operation, and these high altitude TBI mice were randomly divided into normal saline treatment group (TBI group) (n = 6), phenothiazine treatment group (TBI+P) (n = 6), and phenothiazine and UAMC00039 treatment group (TBI+P + U) (n = 6), and 2 mice as a backup. The TBI+P group received 1.25 mg/kg of chlorpromazine and promethazine (1:1) intraperitoneally at 2 hours after TBI and then twice each day for 2 days. TBI+P + U group received 1.25 mg/kg of chlorpromazine and promethazine and 2 mg/kg of UAMC00039 intraperitoneally at 2 hours after TBI and then twice each day for 2 days. The TBI group received the same amount of normal saline.
Hematoxylin–eosin staining
Brain samples of each group were collected at 48 hours after TBI and then fixed, dehydrated, permeabilized, waxed, embedded, and sliced (4 μm). After that, the tissue sections were dried, dewaxed, soaked in gradient ethanol, washed, and stained with hematoxylin–eosin (HE) solution. Subsequently, the sections were dehydrated, permeabilized, sealed, and observed under a microscope (Leica).
Immunohistochemistry
Brain samples were collected at 48 hours after TBI, fixed, dehydrated, permeabilized, waxed, embedded, and sliced (4 μm). Sections were dewaxed, hydrated, heated for antigen retrieval, blocked, and incubated with primary antibodies (Caspase-3, Servicebio, Dpp2/7, Affinity, Bcl-2, Servicebio) and secondary antibodies.
Western blot
The cortical tissues were lysed at 48 hours after TBI, quantitated protein concentration, added loading buffer, and boiled. Protein samples were submitted to SDS-PAGE analysis using a mini-Protean II system (Bio-Rad), electroblotted to PVDF membranes, and blocked by milk. The PVDF membranes were probed with primary antibodies: rabbit anti-Caspase-3 (Servicebio, GB11767C, 1:1000), rabbit anti-Dpp2/7 (Affinity, DF14036, 1:1000), mouse anti-Bcl-2 (Servicebio, 1:2000) and mouse anti-β-tubulin (Servicebio, GB122667, 1:1000), and probed with corresponding second antibodies. The PVDF membranes were visualized by enhanced chemiluminescence, and Image J software was applied for gray-scale quantification of protein bands.
Real-time quantitative polymerase chain reaction
Total RNA was extracted from brain tissues using TRIzol reagent (15,596,026, Invitrogen) at 48 hours after TBI, followed by reverse transcription into cDNA following the manuals of a SweScript All-in-One First-Strand cDNA Synthesis SuperMix for qPCR (Servicebio, G3337). The cDNA was subjected to real-time quantitative polymerase chain reaction (RT-qPCR) using the 2 × SYBR Green qPCR Master Mix (Servicebio, G3320) on an ABI PRISM 7300 RT-PCR system (Thermo Fisher). Three replicates were set up for each well. The gene relative expression was calculated using the 2-ΔΔCt method and standardized by GAPDH. The primer sequences are as follows:
DPP2/7, F, TTCTCTGAAGAACTCCGCCAG, R, TTGCTCTGGATCTGTTGGAAGA.
BCL-2, F, GCTACCGTCGTGACTTCGCA, R, CATCCCAGCCTCCGTTATCC.
CASP3, F, TGGAATGTCATCTCGCTCTGGT, R, GAAGAGTTTCGGCTTTCCAGTC.
GAPDH, F, CCTCGTCCCGTAGACAAAATG, R, TGAGGTCAATGAAGGGGTCGT.
Label-free quantitative proteomics
Total protein was extracted from brain tissues, precipitated, and quantitated protein concentration. The protein samples were enzymatically digested by trypsin, labeled by TMT labeling, and separated by high pH reverse phase chromatography. Mass spectrometry data were collected by imsTOF Pro 2 (Bruker) mass spectrometer and NanoElute liquid phase system. The mass spectrometry data were retrieved using MaxQuant (v1.6.15.0) software and the Andromeda algorithm to obtain a proteome reference database. Using the Diamond program of eggNOG mapper software for functional annotation and enrichment analysis. Proteins with ratios greater than 1.50-fold or less than 1/1.5-fold (p < 0.05) were defined as being significantly differentially expressed.
Statistical analysis
SPSS 21.0 software (IBM) was applied for statistical analysis, using a value of p < 0.05 as an equivalent of statistical significance. The homogeneity of variance was checked before using a parametric test. Measurement data were summarized as mean ± standard deviation. The statistical significance of the differences between the two groups was determined using a t-test. The statistical significance of the differences among more than three groups was determined using one-way analysis of variance with Bonferroni’s multiple comparisons of each two groups.
Results
Phenothiazine minimized neurological injury after high altitude TBI
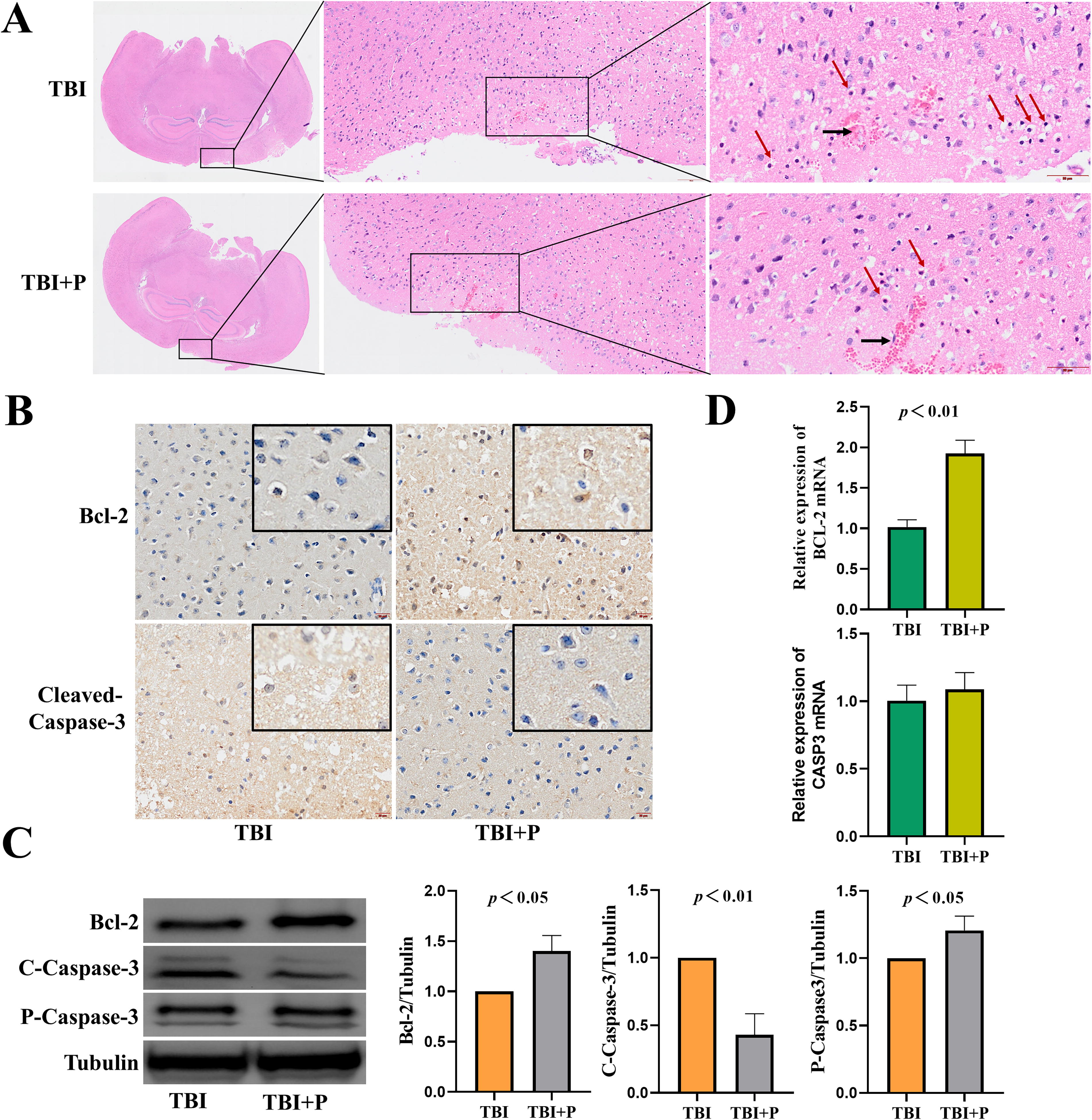
To study the neuroprotective effects of phenothiazine on high altitude TBI mice, we first kept mice in a hypobaric chamber for 7 days under simulated conditions of 5,000 m above sea level, and TBI mice were constructed and treated with chlorpromazine and promethazine subsequently. Finally, the effects of chlorpromazine and promethazine on the cerebral cortex of TBI mice were observed by HE staining and immunohistochemistry (IHC). HE staining demonstrated some micropathological changes in the cortical tissues after 48 hours of TBI. We found that the signs of cytoplasmic vacuolization and nucleus shrinkage (red arrows) were less observed in the cortex of mice treated with chlorpromazine and promethazine than in TBI group (Fig. 1A). As shown in Figure 1B, the IHC assay showed that the chlorpromazine and promethazine treatment increased anti-apoptotic protein Bcl-2 expression and decreased pro-apoptotic protein cleaved-Caspase-3 expression. The results were further confirmed by western blot, and we observed that the chlorpromazine and promethazine treatment group demonstrated a lower level of cleaved-Caspase-3 protein expression than that in the TBI group; however, the level of Bcl-2 protein expression was increased after chlorpromazine and promethazine treatment than that in TBI group (Fig. 1C). In addition, compared with the TBI group, chlorpromazine and promethazine treatment after TBI showed an increase in the level of BCL-2 mRNA (Fig. 1D). There was no significant difference between the TBI group and the chlorpromazine and promethazine treatment group in CASP3 mRNA expression (Fig. 1D). These data indicated that chlorpromazine and promethazine treatment minimized neurological injury after high altitude TBI by suppressing cell apoptosis.
Phenothiazine upregulated Dpp2/7 expression after high altitude TBI
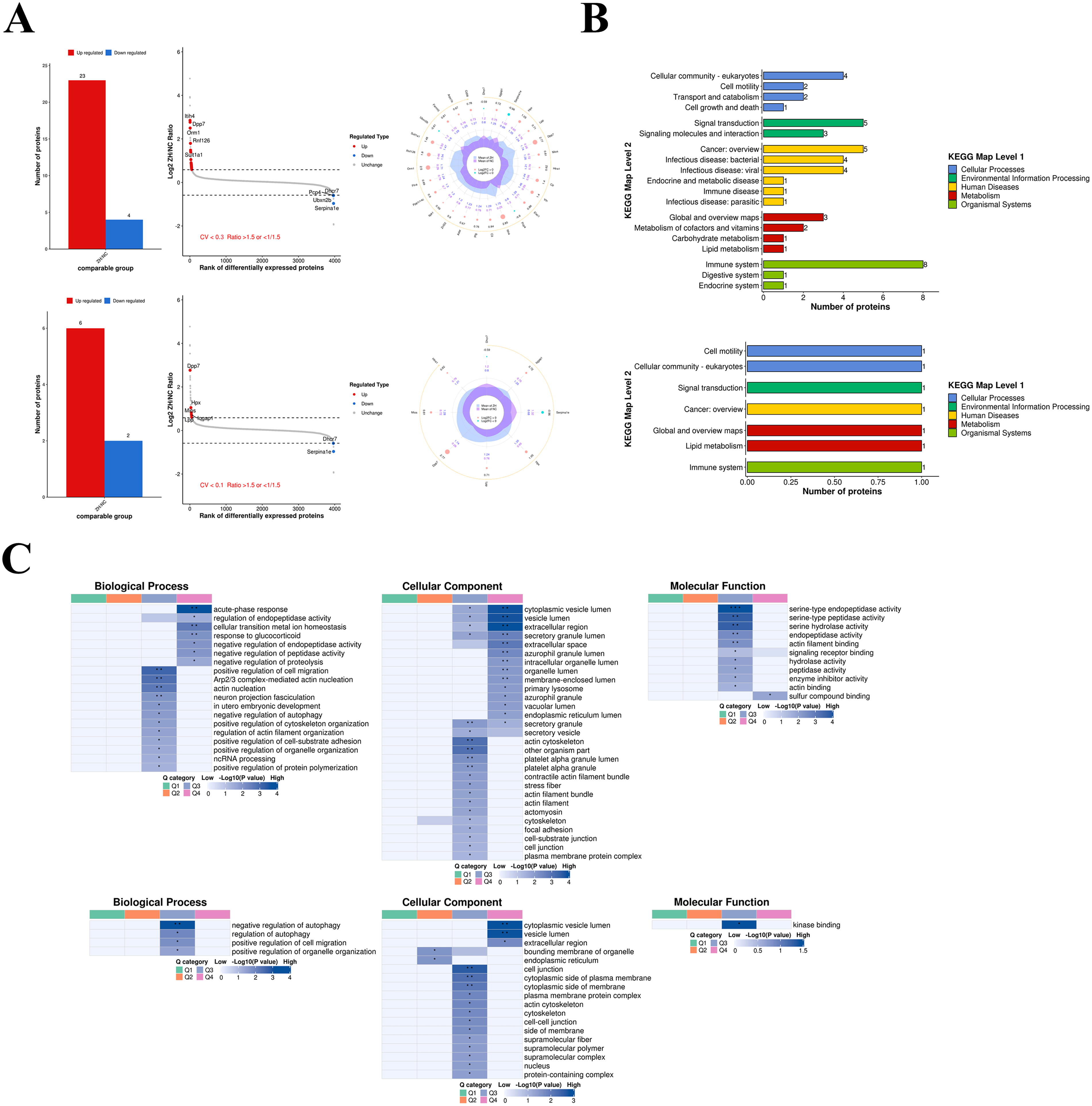
To investigate the changes in the cerebral proteome of phenothiazine treatment after high altitude TBI, we performed label-free quantitative proteomics. We found that the expressions of 27 cerebral proteins were greatly affected by chlorpromazine and promethazine treatment when compared with the TBI group (coefficient of variation [CV] <0.3) (Fig. 2A, upper panel). Furthermore, when CV <0.1, a total of eight cerebral proteins were significantly affected by chlorpromazine and promethazine treatment—six cerebral proteins were upregulated (Dpp2/7, Lpp, Mios, Hpx, Iqgap1, Htra1) and two cerebral proteins were downregulated (Dhcr7, Serpina1e); among them, the most significant upregulated was Dpp2/7 (Fig. 2A, lower panel). Kyoto Encyclopedia of Genes and Genomes pathway database was performed for the functional classification of proteins; a total of 17 proteins were involved in cellular community, cell motility, transport and catabolism, cell growth and death, signal transduction, and signaling molecules and interaction (CV <0.3) (Fig. 2B, upper panel). When CV <0.1, we found only three proteins were involved in cellular community, cell motility, and signal transduction (CV <0.1) (Fig. 2B, lower panel). The cluster analysis was performed to find the functional correlations of differentially expressed proteins, indicating that they were enriched in biological process, cellular component, and molecular function for terms such as regulation of autophagy, positive regulation of cell migration, and positive regulation of organelle organization (Fig. 2C, upper CV <0.3 and lower CV <0.1).
Phenothiazine offered an antiapoptotic effect through Dpp2/7
The above data demonstrated that phenothiazine treatment attenuated neurological injury through decreasing cell apoptosis, and phenothiazine treatment also could upregulate Dpp2/7 expression in high altitude mice. Previous studies have demonstrated that Dpp2/7 is related to cell apoptosis; to verify the Dpp2/7 responsible for the antiapoptotic effect of phenothiazine in high altitude TBI mice, we performed Dpp2/7 inhibitor UAMC00039 to block its biological effect. IHC assay demonstrated that UAMC00039 treatment significantly abrogated the upregulated expression of Bcl-2 protein and downregulated expression of cleaved-Caspase-3 protein induced by chlorpromazine and promethazine treatment in high altitude TBI mice (Fig. 3A). In addition, as shown in Figure 3B, western blot analyses show that UAMC00039 treatment significantly decreased phenothiazine-induced Bcl-2 protein upregulation and restored phenothiazine-induced cleaved-Caspase-3 protein downregulation. Therefore, these data suggested that Dpp2/7 was a key factor in the anti-apoptotic effect of phenothiazine on high altitude TBI mice.

Phenothiazine-induced antiapoptotic effect was attenuated by Dpp2/7 inhibitor in high altitude TBI mice.
Discussion
This study sought to explore phenothiazine-induced neuroprotection effects after TBI at high altitudes. We found that treatment with chlorpromazine and promethazine reduced neurological injury in high altitude TBI mice. This present study further explored the underlying mechanisms and revealed that chlorpromazine and promethazine treatment attenuated cell apoptosis by increasing anti-apoptotic factor Bcl-2 expression as well as suppressing pro-apoptotic protein cleaved-Caspase-3 expression after TBI. Meanwhile, chlorpromazine and promethazine treatment also increased the level of Dpp2/7 protein. Chlorpromazine and promethazine mediated neuroprotection and antiapoptosis effects, Bcl-2 upregulation, and cleaved-Caspase-3 downregulation could be reversed by Dpp2/7 inhibitor UAMC00039. Our study suggested that Dpp2/7/Bcl-2/Caspase-3 was a key pathway by which phenothiazine exerted its neuroprotective effects following TBI at high altitude.
The application of phenothiazine is an efficient therapeutic method in clinics for TBI, ischemic stroke, and hemorrhagic stroke. However, the underlying mechanisms are complex and still not be interpreted completely at present. Previous studies suggest that the hypothermia induced by phenothiazine is key to its potent neuroprotective effects, and pharmacological approach-induced hypothermia has been used worldwide for brain protection after TBI, hypoxic–ischemic encephalopathy, and cardiac resuscitation (Dietrich and Bramlett, 2017). It has been confirmed that therapeutic hypothermia plays a neuroprotective role by reducing the generation of reactive oxygen species and inhibiting the activity of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX) (Han et al., 2021). In addition, phenothiazine-mediated hypothermia provides neuroprotection by reversing ischemic injury-induced hyperglycolysis, gluconeogenesis, and apoptotic cell death (Guan et al., 2022). However, more and more studies have shown that the neuroprotective mechanisms of phenothiazine are only partially dependent on hypothermia. Prior studies have suggested that the neuroprotective effects of phenothiazine are dependent on drug-induced BBB integrity rather than hypothermia; this is achieved by attenuating the increase in aquaporins and matrix metalloproteinases levels after stroke and reversing the decrease in tight junction proteins and basal laminar proteins levels (Li et al., 2019). The neuroprotective effects of phenothiazine are reported to partially depend on attenuating hyperglycolysis and HIF-1-alpha-regulated NOX activation after ischemic stroke (Guo et al., 2021a). Phenothiazine treatment conferred neuroprotection in stroke also by suppressing neuroinflammation and nucleotide-binding oligomerization domain, leucine-rich repeat, and pyrin domain-containing protein 3 inflammasome activation (Guo et al., 2021b). Here, we first found that chlorpromazine and promethazine could provide neuroprotection of high altitude TBI mice by increasing the level of Dpp2/7. Therefore, our study can provide valuable information for the pharmacological mechanisms of phenothiazine in high altitude TBI.
Dpp2/7 is a member of the dipeptidyl peptidase subfamily, lesser known, but nevertheless important (Underwood et al., 1999). Dpp2/7 serves as a quiescent cell proline dipeptidase and is significantly related to mediating the homeostasis of quiescent lymphocytes. The inhibition of Dpp2/7 can induce cell apoptosis in quiescent lymphocytes (Chiravuri et al., 1999). In addition, Dpp2/7 is also essential for maintaining fibroblasts in the G0 phase, and inhibition results in apoptosis that is mediated by the induction of c-Myc and p53 (Mele et al., 2009). Consistent with lymphocytes, inhibition of Dpp2/7 also induces the apoptosis of chronic lymphocytic leukemia B cells in the G0 phase, and this process involves a complex set of proteins, including Syk, p27, p130, and c-Myc (Danilov et al., 2010). The initial finding in our research was that chlorpromazine and promethazine induced the upregulation of Dpp2/7 protein level after TBI, and Dpp2/7 contributed to a reduction in neurological injury via anti-apoptotic effects. Finally, some limitations of our study need to be considered; while we had shown phenothiazine-induced Dpp2/7 upregulation after TBI, it still poorly understood the exact signaling pathways upstream of Dpp2/7. Previous studies have confirmed that the promoter of Dpp2/7 can be activated by Kruppel-like factor 2 and transducer of ERBB2.1 in quiescent lymphocytes (Bista et al., 2008). Therefore, we need to carry out further studies on the regulatory mechanism of phenothiazine on Dpp2/7, and the regulatory mechanism of Dpp2/7 on Bcl-2 and Caspase-3.
Conclusion
In conclusion, the present study suggests that phenothiazine attenuates neurological injury by inducing anti-apoptotic effects in TBI under high altitudes. Mechanistically, phenothiazine promotes Dpp2/7 expression after TBI, and Dpp2/7 suppresses cell apoptosis via the downregulation of the cleaved-Caspase-3 and the upregulation of Bcl-2 expression (Fig. 4). Thus, this study provides a new target with potential application value for the intervention of TBI in high altitude environment.

Schematic diagram of the pathway of neuroprotection from phenothiazine. Phenothiazine suppresses cleaved-Caspase-3 expression while enhancing Bcl-2 expression via upregulating Dpp2/7 expression and ultimately inhibiting cell apoptosis in high altitude TBI mice.
Footnotes
Authors’ Contributions
Q.O. and Y.M. conceived, organized, supervised the work, and wrote and revised the article. W.Z. performed the experiments and prepared and wrote the article. Y.Y. performed the experiments. J.G. and F.H. contributed to the analysis of data. All authors contributed to the article and approved the submitted version.
Data Availability
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
The present study was supported by grants from the Research Project of the General Hospital of Western Theater Command (2021-XZYG-A13 and 2021-XZYG-C37) and the Spark Talent program to Qing Ouyang of General Hospital of the Western Theater Command.