
Abstract
Tian, Lu, Guiqin Liu, Qin Zhao, Junjun Han, Yue Lin, Qian Wang, Qiangqiang Jia, Delong Duo, Duan Yabin, Zhu Junbo, and Li Xiangyang. Pharmacokinetics of midazolam in plasma and brain tissue of rats after exposure to acute and chronic high altitude hypoxia. High Alt Med Biol. 26:273–282, 2025.
Background:
Midazolam effectively improves sleep quality under high altitude hypoxia by reducing central nervous system excitability.
Methods:
Field modeling and sample collection were performed at an altitude of 4,300 m in a high altitude hypoxic environment with a pressure of inspired oxygen of 107 mmHg. Pharmacokinetic alterations of midazolam in high altitude hypoxic rats are determined by high performance liquid chromatography–mass spectrometry. Quantitative real-time polymerase chain reaction and Western blot were used to confirm the connection with drug metabolism and alterations in hypoxia CYP3A4 and P-glycoprotein (P-gp) expression.
Results:
This study demonstrated that high altitude hypoxia increased blood–brain barrier permeability in rats, caused brain tissue damage, and altered the expression of inflammatory cytokines in the brain. In the acute high altitude group and the chronic high altitude group, the area under the curve and Tmax of plasma midazolam revealed substantial increases of 88.6% and 283% and 28.6% and 85.3%, respectively. The clearance rate reduced by 47.3% and 90.0%, while the brain–blood drug concentration ratio (Cbrain/Cplasma) diminished by 11.4% and 82.1%, respectively. The relative expression of CYP3A1 mRNA in the brain tissue of high altitude rats decreased by 42.4% and 66.8%, respectively, and the protein expression was downregulated, while the relative expression of P-gp mRNA increased by 61.3% and 91.2%, respectively (p < 0.05 for all parameters), and the protein expression was upregulated. High altitude hypoxia altered CYP3A1 and P-gp expression and activity, causing alterations in midazolam metabolism.
Conclusions:
This research provided a new reference for the rational use of midazolam in highland areas.
Introduction
Sleep is a vital component of an organism’s health, essential for immunological defense, memory consolidation, recuperation, and control of the endocrine system and body metabolism. Poor sleep quality may cause a significant impact on an individual’s life, increase the chances of developing mental health diseases, and result in significant economic and social burdens (Varghese et al., 2020; Robbins et al., 2022; Tu et al., 2022). There is a substantial association between high altitude and sleep quality, and the body’s rapid eye movement (REM) sleep and non-rapid eye movement (NREM) sleep are altered in the hypoxic environment. The study indicated that mountaineers at an altitude of 3,150 m exhibited a noticeably higher level of alertness, a significant decrease in sleep efficiency, and a modest drop in REM sleep (Tseng et al., 2015). As the altitude climbed from 490 to 4,559 m, the climbers’ sleep efficiency dropped from 91% to 65%, and the quantity of N3 and N4 in NREM was notably diminished (Nussbaumer-Ochsner et al., 2012). Sleeping in high altitude areas, the body’s capacity to adjust to prolonged periods of low oxygen levels gradually enhances. However, frequent awakenings, hypoxemia, and similar symptoms cannot be alleviated. Therefore, the majority of patients opt to use sedative-hypnotic drugs to intervene in their therapy (Beaumont et al., 2004; Tanner et al., 2013; Xiang et al., 2021). In high altitude hypoxia, the use of drugs should be combined with pharmacokinetic alterations, and the dosage and time interval ought to be adjusted according to the actual situation in order to improve the efficacy of the treatments and reduce the toxic side effects. The metabolism of the majority of medications is decelerated in high altitude hypoxia, increasing the mean resident time (MRT), half-life (t1/2), and area under the curve (AUC), while the clearance (CL) is reduced (Zhu et al., 2021). Currently, research on the impact of hypoxia on drug metabolism has mostly centered on strategies to prevent and treat common illnesses in the hypoxic environment. Conversely, there is a dearth of academic research investigating alterations in the pharmacokinetic of sedative-hypnotic drugs in hypoxia. A comprehensive evaluation of the pharmacokinetic changes of these medications in high altitude hypoxia should be conducted to guarantee optimal effectiveness for patients.
Drug-metabolizing enzymes and drug transporters are crucial in the process of drug metabolism. Hypoxia affects the pharmacokinetic characteristics and effectiveness of drugs through altering the expression of metabolic enzymes and transporters (Zhou et al., 2018). Midazolam is a sedative-hypnotic drug primarily metabolized in the liver of the CYP3A4 enzyme (Denisov et al., 2021; Venkatapura, DuBois, Mehvar, 2021). Transport across the blood–brain barrier is a crucial determinant in the penetration of central nervous system (CNS) treatments into the brain to produce their effects. P-glycoprotein (P-gp) is a crucial efflux transporter protein situated at the blood–brain barrier. If the expression of P-gp is increased in high altitude hypoxia, drugs will be transported back to the bloodstream by transporter proteins before entering the brain. This will impact the drug’s delivery to the CNS. Conversely, when the expression of P-gp is decreased, drugs can readily penetrate the brain and induce severe adverse responses. Transportation of midazolam across the blood–brain barrier is crucial for establishing the optimal dosage to provide and attaining the intended therapeutic outcome in high altitude hypoxia. This experiment aimed to examine the impact of high altitude hypoxia on CYP3A4 and P-gp expression. We indicated the alterations in pharmacokinetic parameters of midazolam in the brain and plasma of rats in each group and assessed the impact of hypoxia on drug metabolism, considering the ratio of brain drug concentration to plasma drug concentration, as well as the permeability of the blood–brain barrier. The results of this study provided new guidelines for the appropriate use of midazolam in high altitude areas.
Materials and Methods
Reagents and instruments
Midazolam injection was obtained from Enhua Pharmaceutical Co., Ltd. (Jiangsu, China, Lot: TMZ23A12). Carbamazepine was provided by the China Institute for Food and Drug Control (Lot: 100142 - 202207). Evans blue dye was purchased from Macklin (Shanghai, China, Lot: E2129). Enzyme - linked immunosorbent assay kits, which were used to evaluate IL - 1β, IL - 10, and TNF - α, were from Kexing Trading Co., Ltd. (Shanghai, China, Lot: 24040424N, 24040415N, 24040417N). An RNA extraction kit was sourced from TransGen (Beijing, China, Lot: Q41206). TransScript® One - Step gDNA Removal, cDNA Synthesis SuperMix, and PerfectStart® Green qPCR SuperMix kits were products of TransGen Biotech Co., Ltd. (Beijing, China). The primers used in quantitative real-time polymerase chain reaction were synthesized by Takara (Kyoto, Japan). A Bicinchoninic acid (BCA) protein concentration determination kit was obtained from Solarbio (Beijing, China, Lot: PC0020). A rat anti - CYP3A1 primary antibody was purchased from Abcam (USA, Lot: AB22724), a rat anti - P-gp primary antibody was from Abcam (USA, Lot: AB170904), a rat anti - β - actin primary antibody was from Immunoway (Texas, USA, Lot: YT0099), and a goat anti - rabbit Immunoglobulin G - Horseradish Peroxidase (IgG-HRP) secondary antibody was from PhD Bioengineering Co., Ltd. (Wuhan, China, Lot: AS070). An Ultimate 3,000 Ultra-High Performance Liquid Chromatograph (Thermo Fisher Scientific, USA), a Q Exactive Focus Electrostatic Field Orbitrap High Resolution Mass Spectrometer (Thermo Fisher Scientific, USA), and a Sorvall ST4 Plus centrifuge (Thermo Fisher Scientific, USA) were used in the experiment.
Grouping and modeling of experimental animals
Male Sprague - Dawley (SD) rats weighing between 180 and 220 g were purchased from the Laboratory Animal Centre of the Department of Medicine at Xi’an Jiaotong University (animal certificate number: 2,019-0008, Xi’an, China). During a 12-hour period, sunlight and darkness alternated, with a temperature of (22 ± 2)°C and a humidity of (55 ± 10%). All rats were individually housed in separate cages and provided ad libitum access to water and food pellets. In this experiment, SD rats were randomly divided into three groups by adopting high altitude hypoxic field modeling: the low altitude control group (LC, Xi’an, Shanxi province, altitude: 390 m, pressure of inspired oxygen [PIO2]: 154 mmHg), the acute high altitude hypoxia group (AHH, Maduo County, Qinghai Province, low altitude rats lived for 3 days after entering Maduo County, altitude: 4,300 m, PIO2: 107 mmHg), and the chronic high altitude hypoxia group (CHH, Maduo County, Qinghai Province, low altitude rats lived for 30 days after entering Maduo County, altitude: 4,300 m, PIO2: 107 mmHg). Animal research strictly adheres to the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and the protocol was approved by the Animal Ethics Committee of Qinghai University (Permit NO. 2,022-24).
Hematoxylin and eosin stain
The rats were intraabdominally injected with (0.7 ± 0.1) of a 10% solution of chloral hydrate and kept under anesthesia. The heart was infused with warm saline, and the brain tissue was extracted and placed in a 4% paraformaldehyde solution. Brain tissue sections were dehydrated using anhydrous ethanol and rendered transparent with xylene, immersed in hematoxylin solution for 15 minutes, rinsed in running water, stained in eosin solution for 5 minutes, rinsed again in running water, further dehydrated and made transparent, and finally sealed with neutral gum. The morphological alterations in the brain tissue of rats from each group were observed using a light microscope, and images were captured.
Evans blue staining
The fluorescence intensity of Evans blue in brain slices was examined using the Evans blue staining method in combination with confocal laser scanning microscopy. Rats were administered a 2% solution of Evans blue dye (2 ml/kg) via injection into the tail vein. After 45 minutes, the rats were sacrificed by cervical vertebra dislocation, and brain tissues were promptly collected. Fluorescence images of the Evans blue dye in brain slices were captured using confocal imaging.
Enzyme-linked immunosorbent assay
Take the IL-10, IL-1β, and TNF-α kits and prepare for backup. Set up the standard and sample wells, adding 50 μl of different concentrations of standards into each standard well; subsequently, add 10 μl of the sample for analysis, followed by 40 μl of sample diluent to the sample wells. Then, dispense 100 μl of the enzyme reagent into each well, ensuring that the sample wells are pre-warmed and thoroughly rinsed. Each well was supplemented with 50 μl of chromogen A and chromogen B, mixed delicately, and the absorbance optical density (OD) value was quantified at 450 nm using an enzyme marker. Standard linear regression curves were plotted, and concentration values were calculated for each sample. The standard curve equations for IL-10, IL-1β, and TNF-α were: Y = 29.3x − 1.69, R2 = 0.999, Y = 271x − 4.60, R2 = 0.998, Y = 212x − 8.15, R2 = 0.996.
Pharmacokinetics
Biological sample collection
Midazolam was administered in each group of rats at a dosage of 50 mg/kg in a single gavage after a 12-hour fasting period and water restriction. Blood samples of 0.5 ml were taken from the jugular plexus at distinct time intervals: 0.08 hour, 0.17 hour, 0.5 hour, 1 hour, 1.5 hours, 2 hours, 4 hours, 6 hours, 8 hours, and 12 hours following drug administration. The samples were placed into a centrifuge tube filled with sodium heparin anticoagulant and subjected to centrifugation at 120,000 rpm at 4°C for 10 minutes. The plasma was then separated and kept at −20°C. Following the blood sample from each rat, brain tissue was immediately dissected and kept in liquid nitrogen.
Biological sample preparation
The plasma sample was pipetted into a 2 ml centrifuge tube. Then, 400 μl of methanol and 100 μl of the internal standard solution were added to the tube. The mixture was shaken for 30 seconds and then centrifuged at 14,000 rpm for 10 minutes. Subsequently, 2 μl of the supernatant was transferred into the sample bottle. The 0.1 g of brain tissue was weighed and placed in a 2 ml centrifuge tube. After that, 1.5 ml of methanol was added to the tube for homogenization. Then, 100 μl of the homogenate was taken and combined with 500 μl of the internal standard solution. The resulting mixture was shaken for 30 seconds and then centrifuged at 14,000 rpm for 10 minutes. Finally, a volume of 2 μl of the supernatant was transferred into the sample bottle.
Chromatography and mass spectrometry conditions
The detection ion pair of midazolam was set at m/z 326, while the detection ion pair of carbamazepine, which was used as an internal standard, was set at m/z 237. Hypersil GOLD™ C8 HPLC Columns (Thermo Fisher Scientific; dimensions: 2.1 × 50 mm, inner diameter of 1.9 μm) were employed, and the column temperature was maintained at a constant 35°C. The mobile phase consisted of formic acid (0.1%) - methanol in a volume ratio of 35:100 (v/v), and the flow rate was kept constant at 0.25 ml/min. The multi-reactive ion monitoring mode was utilized. The ion spray voltage was set at 3,000 V, and the ion transfer tube temperature was set at 350°C.
Cerebral blood ratios and permeability of the blood–brain barrier
A graph was constructed to illustrate the temporal variation of the brain/plasma drug concentration ratio. This ratio was derived based on the drug concentrations determined in plasma samples and brain tissue specimens. The permeability of midazolam across the blood–brain barrier was assessed by determining the brain/plasma partition coefficient KP (AUCbrain/AUCplasma).
Quantitative real-time polymerase chain reaction
One hundred grams of liver and brain were collected, and total RNA was extracted using the kit. The extracted RNA was diluted to the appropriate multiplicity, and the concentration of the RNA solution was determined. Conduct reverse transcription following the guidelines of the TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix kit. Conduct the amplification procedure in accordance with the guidelines for the PerfectStart Green qPCR SuperMix kit. Two-step amplification was performed using a Roche Light Cycler 96 real-time fluorescence quantitative polymerase chain reaction (PCR) instrument. The PCR program settings are as follows: pre-denatured cycle (1 cycle): 94°C for 30 seconds; PCR cycle (45 cycles): 94°C for 5 seconds, 60°C for 30 seconds; and dissolution curve: 60°C–95°C for 30 seconds. Fold induction values were calculated using the value of 2−ΔΔCt and the primer sequences are listed in Table 1.
Rat Primers Used for RT-qPCR Analysis
RT-qPCR, quantitative real-time polymerase chain reaction.
Western blot
Total protein was extracted from 200 mg of liver and brain tissue from each group of rats, respectively, utilizing the BCA protein concentration assay kit for measurement. Polyacrylamide gel electrophoresis was used to separate the protein samples. Following electrophoresis, the protein bands were transferred onto polyvinylidene fluoride membranes using a steady current of 280 mA. The membranes were then immersed in a 5% solution of skimmed milk powder for 2 hours. The membrane was subjected to overnight incubation at 4°C with primary antibodies that specifically target CYP3A1 (1:2,000) and P-gp (1:1,000). Subsequently, the membrane underwent five washes with Tris - Buffered Saline with Tween 20 (TBST) solution, each lasting 3 minutes. Following a 40-minute incubation period with the corresponding secondary antibody (1:10,000), the membrane was subsequently rinsed five times. Subsequent exposures were made in a dark room with a supersensitive luminescent solution, and finally the images were analyzed. Gray value analysis of the bands of the target protein samples was performed on the obtained images using Image J software. The relative expression of CYP3A1 and P-gp proteins was then determined by calculating the relative expression using β-actin as an internal reference.
Statistical analysis
All quantitative data are expressed as mean ± standard error of the mean. The data were analyzed using one-way analysis of variance in the statistical package for social sciences version 27.0 (IBM). Differences between the means of the two groups were compared using least significant difference post hoc tests. The results were considered statistically significant when p < 0.05.
Results
Effects of high altitude hypoxia on brain tissue pathology
Hematoxylin and eosin staining was used to examine the morphology of high altitude hypoxic rat brain tissue. The results are shown in Figure 1A–C. LC group cells exhibited homogeneous staining, complete morphology, distinct borders, and nuclei characterized by a generally rounded shape and even distribution, without any apparent pathological alterations. In both the AHH and CHH groups, dark-colored neurons were sporadically observed. These cells exhibited irregular cell morphology, smaller size, deeper color, blurred internal cell structure, evident structural disorder of the cytosol, and perivascular gaps. However, no notable expansion of glial cells or infiltration of inflammatory cells was seen. The regions shown by yellow arrows in Figure 1B and Figure 1C exhibited unmistakable abnormalities in cell structure and a reduction in the size of nuclei in the cytosol tail, indicating that hypoxia impacts the brain tissue of rats.

Observation of brain tissue in rats at different hypoxic environments.
Effects of high altitude hypoxia on blood–brain barrier integrity in rats
Blood–brain barrier permeability was measured using the Evans blue perfusion method in high altitude hypoxic rats. The results are shown in Figure 2B, C. Both the AHH and CHH groups exhibited notable Evans blue extravasation and modified blood–brain barrier permeability in comparison to the LC group shown in Figure 2A.
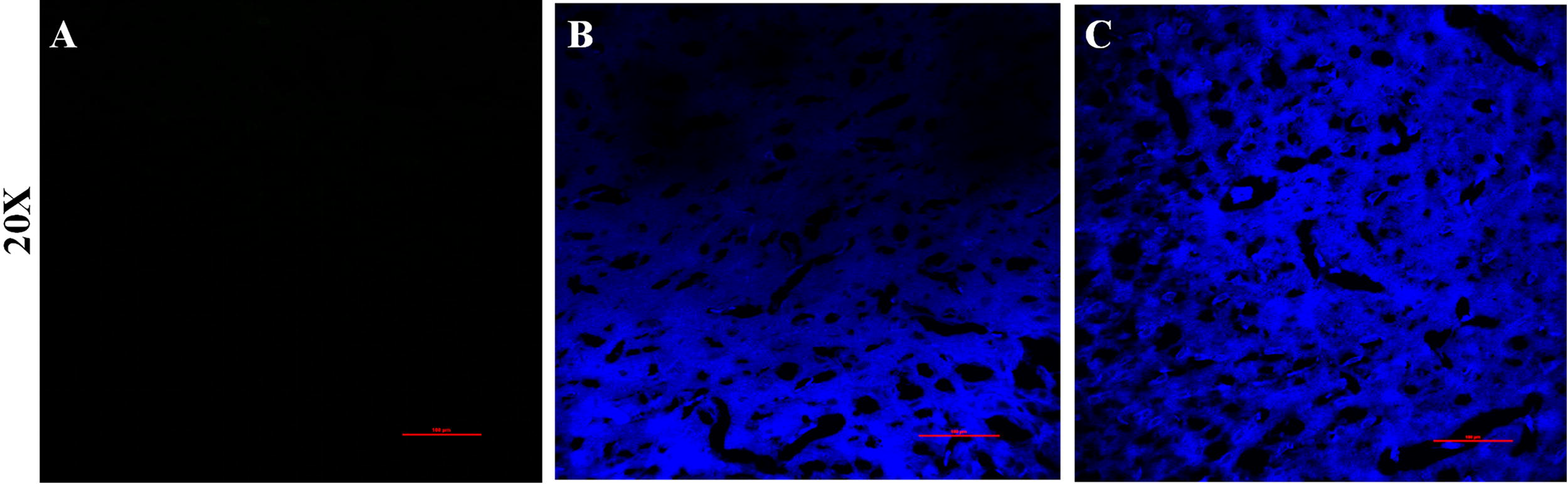
Evans blue staining of brain tissue from rats at different hypoxic environments.
Effects of high altitude hypoxia on inflammatory cytokines in rat brains
Effects of high altitude hypoxia on inflammatory cytokine expression in rat brain tissue were investigated by enzyme-linked immunosorbent assay detection of IL-10, IL-1β, and TNF-α. Compared with the LC group, the AHH and CHH groups showed a significant decrease (p < 0.05) in the concentrations of IL-10 in the brain tissue of rats, as shown in Figure 3A. Conversely, the concentrations of IL-1β and TNF-α were significantly increased (p < 0.05), as shown in Figure 3B, C.
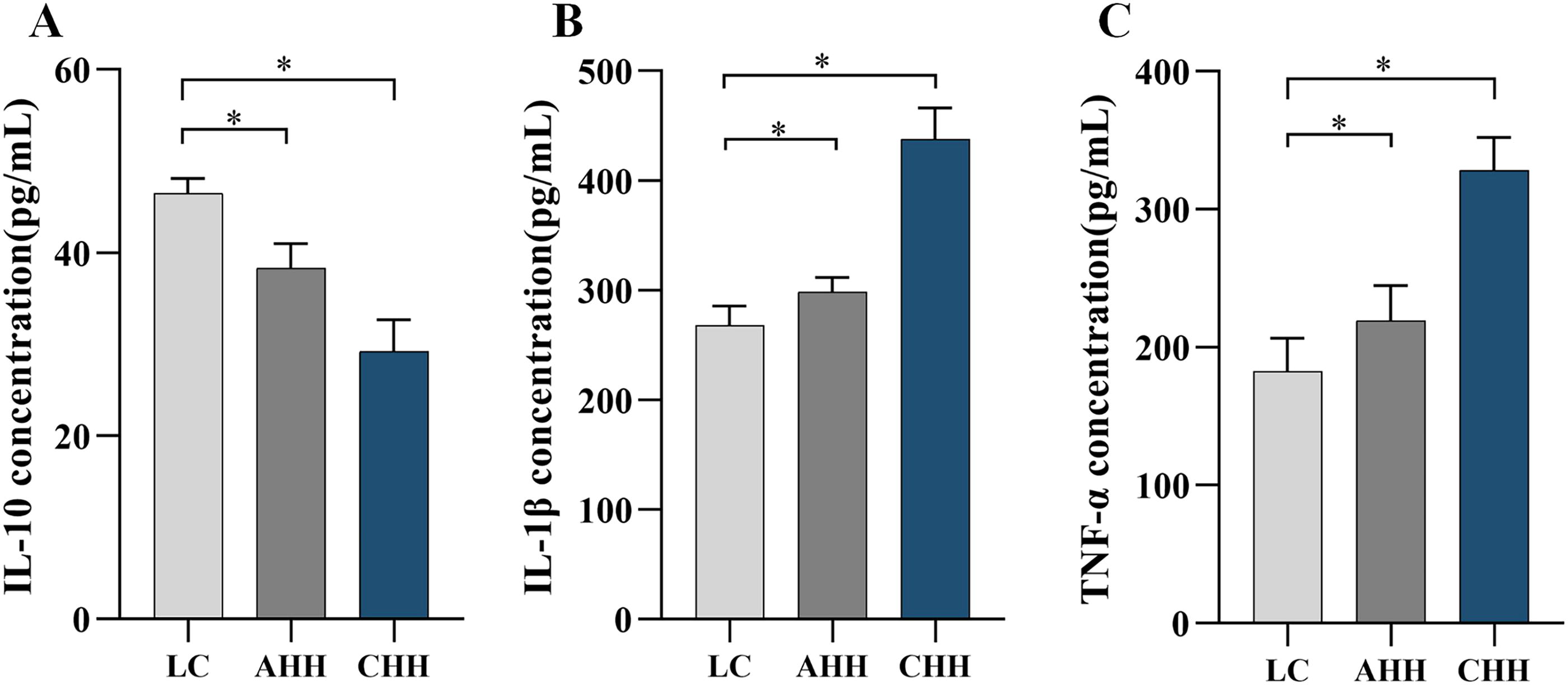
Alterations of inflammatory indexes in rat brain tissue after exposure to hypoxia.
High performance liquid chromatography–mass spectrometry method validation
Based on the biological sample handling methods and test conditions described above, endogenous chemicals in plasma and brain do not disrupt chromatography. The retention period of midazolam in both plasma and brain was 4.96 minutes, as shown in Figure 4. The linear regression equations were as follows: Y = 0.0007X + 0.0002, R2 = 0.999, Y = 0.0004X − 0.0006, R2 = 0.998. Both plasma and brain exhibited strong linearity within the range of 1.17–300 ng/ml, with a minimum quantification limit of 1.17 ng/ml determined in both instances. The intra- and inter-day precision and accuracy satisfied the criteria for determining biological samples. For the substances to be assessed under each assay setting, the impact of the matrix effect was insignificant, the recoveries were acceptable, and the experimental method was repeatable. The stability of the mixed-quality control samples containing low and high doses of midazolam was examined by subjecting them to conditions of being left at room temperature for 8 hours, undergoing three times of freezing and thawing at −20°C, and then being left at −20°C for 30 days. The results indicated that the samples exhibited excellent stability following experimental treatment under various settings.
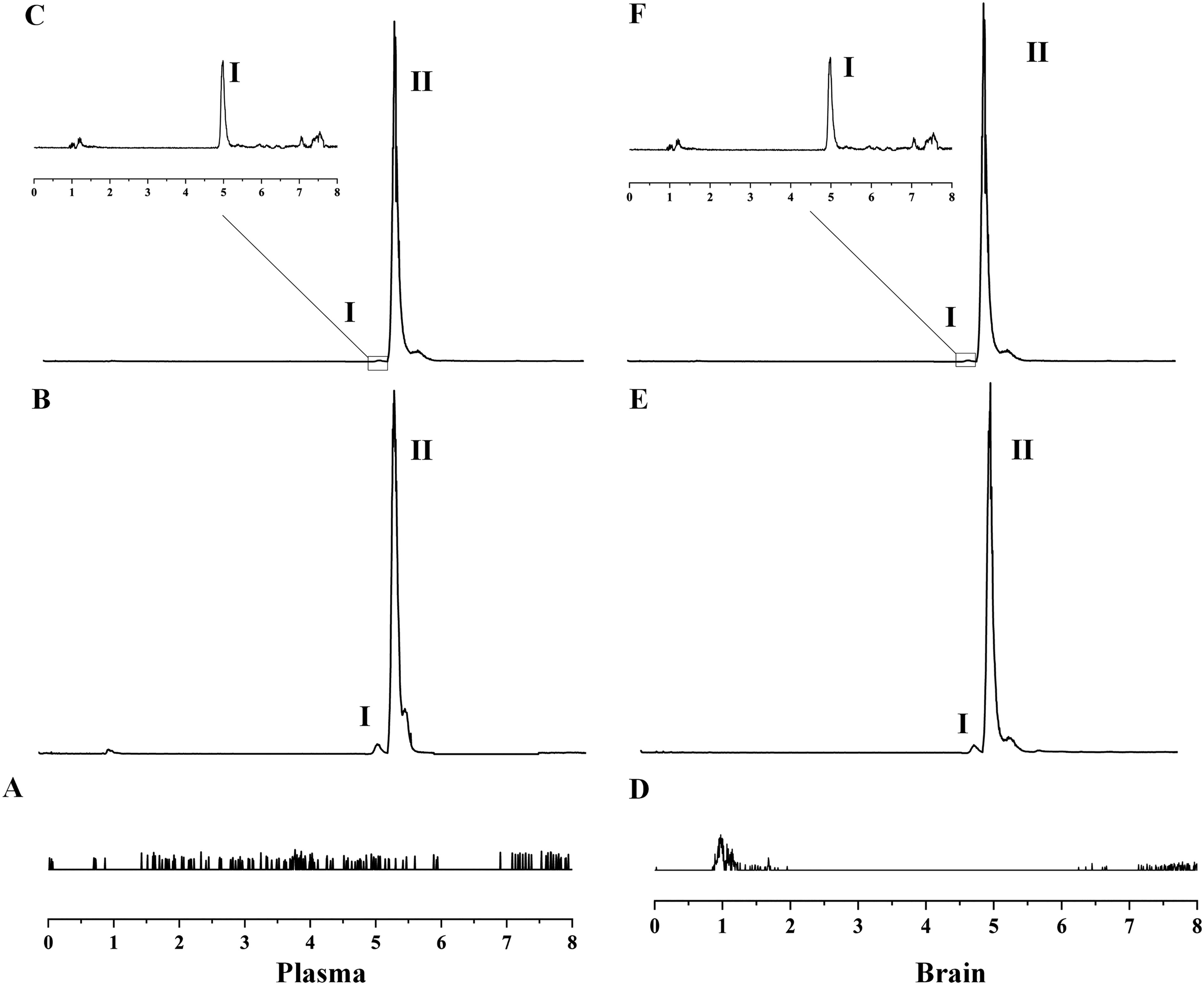
Midazolam (I). Carbamazepine (II).
Effects of high altitude hypoxia on the pharmacokinetics of midazolam in rats
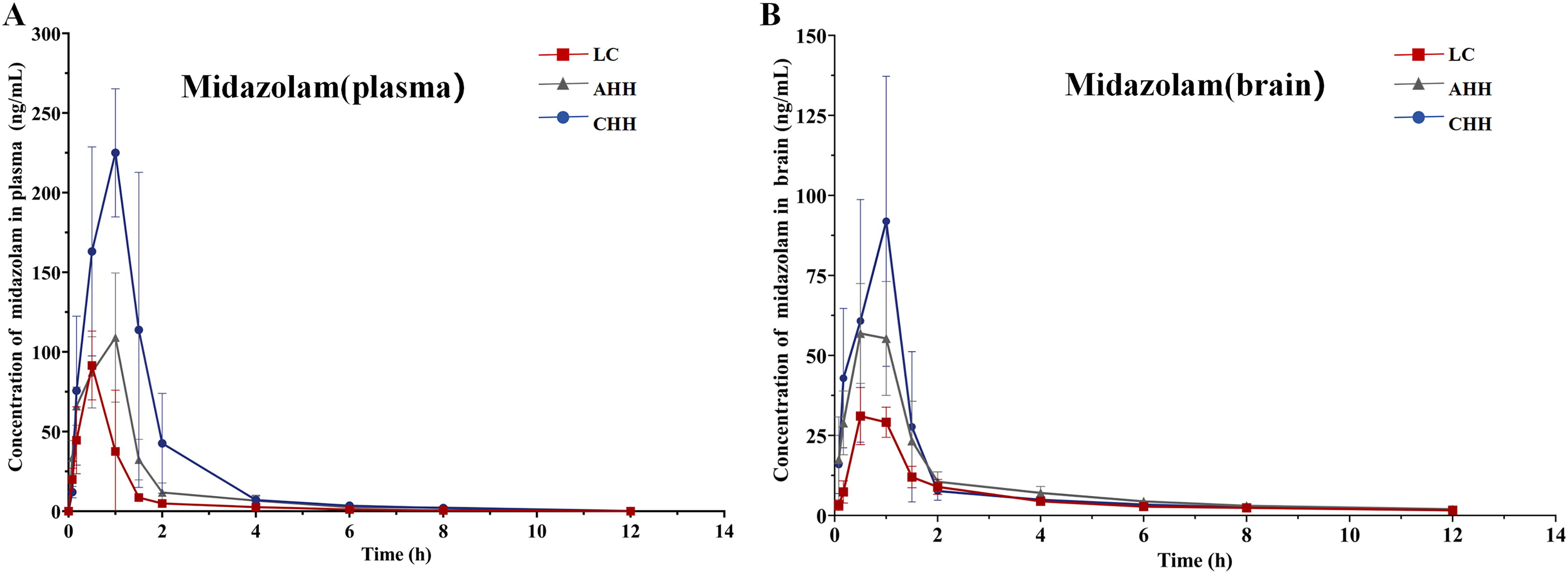
The pharmacokinetic characteristics of midazolam in rat plasma and brain tissue are shown in Tables 2 and 3. Figure 5 illustrated the mean blood concentration time curves of midazolam in plasma and brain tissue.
Pharmacokinetic Parameters of Midazolam in Rat Plasma After Exposure to Hypoxia
*p < 0.05 compared with the LC group.
Data are expressed as the mean ± SD values (n = 6).
AHH, acute high altitude hypoxia group; AUC, area under the concentration time curve; CHH, chronic high altitude hypoxia group; CLz/F, total plasma clearance; Cmax, the maximum plasma concentration; ke, elimination rate constant; LC, low altitude control group; MRT, mean residence time; SD, standard deviation; t1/2, half-life; Vz/F, apparent volume of distribution.
Pharmacokinetic Parameters of Midazolam in Rat Brain After Exposure to Hypoxia
*p < 0.05 compared with the LC group.
Data are expressed as the mean ± SD values (n = 6).
Compared with the LC group of midazolam, the AUC and Tmax of rat plasma in both the AHH and CHH groups were significantly increased, and CL was markedly decreased. Particularly, AUC was increased by 88.6% and 283% (p < 0.05), Tmax was increased by 28.6% and 85.3% (p < 0.05), and CL was decreased by 47.3% and 90.0% (both p < 0.05). In the AHH group, MRT was increased by 37.6% (p < 0.05). In the CHH group, Vz/F was decreased by 58.6% (p < 0.05) and Cmax was increased by 136% (p < 0.05).
Compared with the LC group of midazolam, the AUC and Cmax of rat brain in both AHH and CHH groups were significantly increased, while MRT and CL were significantly decreased. Specifically, AUC was increased by 69.7% and 87.3% (both p < 0.05), Cmax was increased by 103% and 243% (both p < 0.05), MRT was decreased by 15.3% and 33% (both p < 0.05), and CL was decreased by 40.9% and 46.4% (both p < 0.05). In the CHH group, t1/2z was increased by 28.1% (p < 0.05).
Effect of high altitude hypoxia on the cerebral blood concentration ratio of midazolam
The brain–blood drug concentration ratio (Cbrain/Cplasma) was determined by the drug concentrations in brain tissue and plasma in the high altitude hypoxia, as seen in Figure 6A. Compared with the LC group, the AHH and CHH groups (Cbrain/Cplasma) significantly decreased, with the CHH group demonstrating reductions of 168%, 40.5%, 27.8%, 68.6%, and 18.1% after 1.5, 2, 4, 6, and 8 hours post-drug administration, respectively (all p < 0.05). The brain–blood partition coefficient KP (AUCbrain/AUCplasma) was measured as shown in Figure 6B, compared with the LC group, demonstrating reductions of 11.4% and 82.1% (p < 0.05) in the AHH and CHH groups.

Effect of high altitude hypoxia on the expression of CYP3A1 and P-gp mRNA in rat brain and liver
Quantitative real-time polymerase chain reaction results showed that the relative expression of CYP3A1 mRNA in the brains of rats subjected to AHH and CHH groups decreased by 42.4% and 66.8%, compared with the LC group (both p < 0.05) (Fig. 7A), while in the liver, it decreased by 63.5% and 77.0% (both p < 0.05) (Fig. 7B). The relative expression of P-gp mRNA in the brain increased by 61.3% and 91.2% (both p < 0.05) (Fig. 7C), while it decreased by 44.6% and 61.5% in the liver (both p < 0.05) (Fig. 7D).
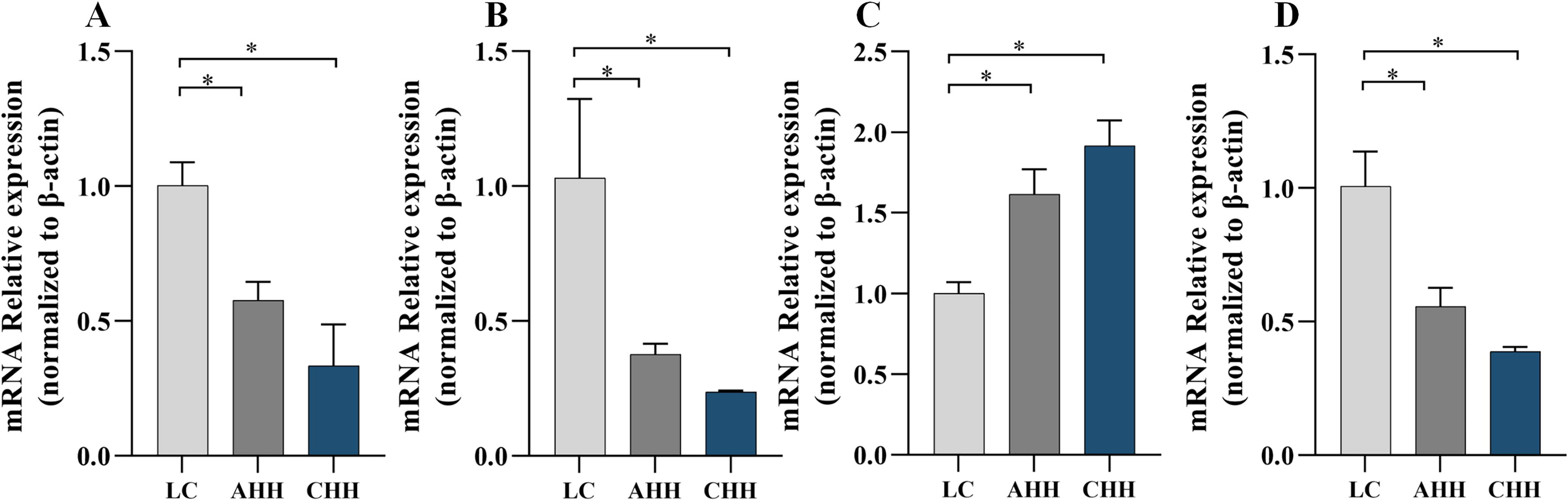
The effect of different hypoxic environments on CYP3A1 and P-gp mRNA expression in rat brain and liver.
Effect of high altitude hypoxia on CYP3A1 and P-gp protein expression in rat brain and liver
The Western blot results showed a decrease in the relative expression of CYP3A1 protein in both the brain (Fig. 8A) and liver (Fig. 8B) of rats subjected to AHH and CHH groups in comparison with the LC group. The relative expression of P-gp in the brain increased by 8.89% and 27.9% (p < 0.05) (Fig. 8C), while the relative expression of P-gp in the liver decreased by 27.5% and 65% (p < 0.05) (Fig. 8D).

The effect of different hypoxic environments on CYP3A1 and P-gp protein expression in rat brain and liver.
Discussion
Midazolam is extensively used for sedation and hypnosis due to its brief half-life, high water solubility, and rapid elimination rate. While midazolam has been subject to thorough investigation as an active indicator of CYP3A, there is a scarcity of information regarding its metabolism in the brain. This study provided a highly sensitive high performance liquid chromatography–mass spectrometry technique specifically designed for investigating the metabolism of midazolam in rats. As midazolam exerts its effects on the CNS and requires access to the brain in order to interact with the target receptors, we also determined the drug concentration in the brain. Figure 5 showed that midazolam had a lower concentration in the brain compared with plasma when it entered the absorption phase. During the clearance phase of the drug, plasma concentration progressively diminished while remaining high in the brain; this may explain the necessity for re-dosing CNS therapeutic drugs with narrow therapeutic indices under hypoxic conditions at high altitude (Luszczki et al., 2018; Zhang et al., 2020). Furthermore, the current research revealed that MRT in the brain was significantly higher than in plasma, possibly resulting in an extended period of pharmaceutical impact on the CNS. According to this study, it is suggested to exercise caution when determining the scheduled time intervals for administering midazolam in highland regions, as it has a significant risk of leading to the accumulation of the medication in the brain. Benzodiazepines are the main representatives of CNS depressants, enhancing γ-aminobutyric acid (GABA)-mediated central inhibition. Investigations have demonstrated that in the low-oxygen conditions of a high altitude, the inhibitory neurotransmitter GABA tends to accumulate (Nilsson and Lutz, 1993). Nevertheless, further study is needed to determine if the buildup of GABA enhances the CNS depressive effects of sedative-hypnotics under hypoxic conditions.
Permeability of CNS medications across the blood–brain barrier is crucial for establishing the appropriate dosage to deliver and delivering the intended therapeutic effect in the high altitude hypoxic environment. The empirical findings of this study indicated that the permeability of the blood–brain barrier rises following exposure to hypoxia, which results in an augmentation of drug concentration within the brain. Nevertheless, when the organism was exposed to hypoxia, the expression of the drug metabolism enzyme CYP3A4 decreased and the expression of the exocytosis transporter protein P-gp increased. Ultimately, this also results in decreased the brain–blood drug concentration ratio, diminished permeability of the blood–brain barrier, and modified drug metabolism. Cytokines have been proven to influence drug-metabolizing enzymes in human hepatocytes, with pro-inflammatory cytokines such as IL-6 and TNF-α demonstrated to downregulate CYP3A mRNA levels (Morgan, 1997). This study indicated a significant elevation in TNF-α levels due to high altitude hypoxia; however, the regulatory role of TNF-α and other cytokines on the expression of drug-metabolizing enzymes under high altitude hypoxia requires additional examination. Besides the liver, many CYP enzymes are discovered in the brain and play an essential part in drug metabolism (Pai et al., 2002; Ferguson and Tyndale, 2011; McMillan and Tyndale, 2018; Fanni et al., 2021; Kuban and Daniel, 2021). The study revealed that high altitude hypoxia decreased the expression of CYP3A4 mRNA and protein in the rat brain and liver. However, P-gp expression significantly increased in the brain but decreased in the liver. This may be due to variations in the distribution and function of P-gp in different tissues. Drug-metabolizing enzymes and drug transporters are intricately associated with drug metabolism in the high altitude hypoxic conditions. Exploring the regulation of drug-metabolizing enzymes and transporter proteins under low oxygen conditions may provide novel approaches for investigating intracerebral drug delivery in the CNS.
Conclusion
In this experiment, high altitude hypoxia was integrated with drug metabolism. The effects of high altitude hypoxia on the histopathological morphology of brain tissue, the permeability of the blood-brain barrier, and the expression of inflammatory cytokines in the brain were investigated. Additionally, the changes in the pharmacokinetic parameters of midazolam in plasma and brain under high altitude hypoxic conditions were determined, along with the trend of the cerebral blood drug concentration ratio. The involvement of relevant drug-metabolizing enzymes or drug transporters in the modulation of drug metabolism was verified. This study demonstrated that high altitude hypoxia increased blood–brain barrier permeability in rats, caused brain tissue damage, and altered the expression of inflammatory cytokines in the brain. The hypoxic environment affects the expression and activity of relevant drug-metabolizing enzymes and transporters, leading to modified metabolism of midazolam and a diminished ratio of intracerebral drug concentration to plasma drug concentration. This research provided innovative insights into the treatment of CNS disorders in highland regions and offers a new reference for the rational use of midazolam in highland areas.
Footnotes
Authors’ Contributions
L.T. designed, developed, and optimized the assays and contributed to writing—original draft and writing—review and editing. G.L. analyzed the data and contributed to figure making. Q.Z. analyzed samples. J.H. collected samples. Y.L. collected samples. Q.W. collected samples. Q.J. provided support to the assay development. D.Y. collated data. D.D. analyzed the data and contributed to editing. Z.J. analyzed the data and contributed to editing. L.X. contributed to review and editing, conceptualization, and supervision and provided financial support.
Data Availability
The data that support the findings of this study are available on request from the corresponding author.
Author Disclosure Statement
None of the authors have any conflict of interest, financial or otherwise, to disclose. All authors have approved the contents of this article.
Funding Information
This work was supported by the National Natural Science Foundation of China [No. 82260731] and the Central Government guided Local Science and Technology Development Foundation [No. 2024ZY001].