
Abstract
Asthma, a chronic helper T cell type 2-mediated inflammatory disease, is characterized by airway hyperresponsiveness and inflammation. Growing evidence suggests that increased expression of acidic mammalian chitinase (AMCase) may play a role in the pathogenesis of asthma. In the present study, we sought to develop an RNA interference approach to suppress allergic asthma in mice through silencing of AMCase expression. Mice sensitized with ovalbumin (OVA) were intratracheally administered a recombinant adeno-associated virus expressing short hairpin RNA (rAAV-shRNA) against AMCase. In OVA-sensitized mice, the development of allergic symptoms was significantly associated with elevated AMCase expression. After administration of rAAV-shRNA, there was a significant reduction of AMCase expression in the lung and in bronchoalveolar lavage fluid (BALF) cells of sensitized mice. Sensitized mice receiving rAAV-shRNA showed a significant improvement in allergic symptoms, including airway hyperresponsiveness (AHR), eosinophil infiltration, eotaxin, interleukin-13 secretion in BALF, and serum OVA-specific IgE level. Our data suggest the hyperexpression of AMCase in asthma can be suppressed by rAAV-mediated shRNA. Silencing AMCase expression by shRNA may be a promising therapeutic strategy in asthma.
Introduction
RNA interference (Fire et al., 1998) has become a powerful tool for modulating gene expression in cells (Cullen, 2006). Because short hairpin RNA (shRNA) has been shown to be efficiently processed into small interfering RNA (siRNA) in mammalian cells, strategies for producing siRNAs have been focused on transcription of shRNA, using RNA polymerase III promoters such as U6 and H1 (Storvold et al., 2007; Lee et al., 2008b). Moreover, viral vectors appear to have the highest efficiency in delivering siRNA into mammalian cells (Li et al., 2006; ter Brake et al., 2008). Despite potential immunogenicity problems generated by some viral vectors (Zaiss and Muruve, 2008), adeno-associated viral (AAV) vectors offer a compromise between an adequate level of transduction and an acceptable safety profile (Leung et al., 2007). Hence, several reports have successfully used an AAV-mediated shRNA therapeutic system to control viral infections (Moore et al., 2005; Fechner et al., 2008) and genetic disorders (Franich et al., 2008). In the present study, we hypothesized that specific suppression of elevated AMCase can be achieved by AAV-mediated shRNA production, which would therefore result in reduced eosinophilia and Th2-mediated airway inflammation in a mouse model of asthma. We further investigated whether the inhibition of AMCase may be associated with reduced expression of IL-13, eotaxin, and other proinflammatory molecules.
Materials and Methods
Cell culture
3T3 and 293T cell lines were cultured and maintained in Dulbecco's modified Eagle's medium (DMEM; GIBCO Invitrogen, Carlsbad, CA) containing 10% fetal bovine serum (FBS; HyClone, Logan, UT) with 5% CO2 in humidified air at 37°C.
Cloning of AMCase and setup of AMCase-overexpressing cell line
RNA was extracted from mouse lung tissues, using the TRIzol reagent (Invitrogen) and reverse-transcribed (RT) with a two-step RT-polymerase chain reaction (PCR) kit (Invitrogen). The full-length AMCase sequence was amplified with the following primers: forward, 5′-ATCAGAATTCTATGGCCAAGCTACTTCTC-3′; and reverse, 5′-TTTCTGCGGCCGCATGGCATTAGGTTCATGGC-3′. The AMCase-overexpressing cell line was established by transfection into 3T3 cells of a pTriEx-neo vector carrying the AMCase sequence. A dilution series under G418 selection pressure was applied.
Real-time PCR
Expression of AMCase, humanized Renilla green fluorescent protein (hrGFP), and actin was analyzed by real-time PCR, using a LightCycler PCR system (Roche, Indianapolis, IN). Actin 300 (forward primer, 5′-GAAACTACATTCAATTCCATC-3′; reverse primer, 5′-CTAGAAGCACTTGCGGTGCAC-3′) was used as a housekeeping gene. The reaction parameters for actin 300 and hrGFP amplification (forward primer, 5′-ATGGTGAGCAAGCAGATCCTG-3′; reverse primer, 5′-GGTGCGCTCGTACACGAAGCC-3′) were as follows: initial denaturation at 95°C for 10 min, followed by 35 cycles at 95°C for 10 sec, 50°C for 10 sec, and 72°C for 10 sec. AMCase was amplified using the following primers: forward, 5′-TGGACACACCTTCATCCTGA-3′; and reverse, 5′-AACAAGCCCTGCTTGACAAT-3′. After initial denaturation at 95°C for 10 min, 45 cycles were performed at 95°C for 10 sec, 50°C for 10 sec, and 72°C for 10 sec.
Design and construction of shRNA
Three sequences specifically targeting murine AMCase were designated as siRNA185, siRNA897, and siRNA1336 (siRNA185: top, GCAGAACAAUGAGAUCACCACCAUA; bottom, UAUGGUGGUGAUCUCAUUGUUCUGC; siRNA897: top, GGGUUCUGGGCCUACUAUGAGAUUU; bottom, AAAUCUCAUAGUAGGCCCAGAACCC; siRNA1336: top, GGCAGUGCAUCAAUGGAAUCACAUA; bottom, UAUGUGAUUCCAUUGAUGCACUGCC). shRNA molecules with the same sequences as siRNA, but containing a random eight-nucleotide (CAAGCTTC) loop structure and a four-nucleotide terminator (3′-TTTT) overhanging the 3′ end, were inserted into a pCI-neo plasmid vector with an mU6 promoter. Here an shRNA-targeting luciferase (Luciferase: top, 5′-TCCAATTCAGCGGGAGCCTTCAAGAGAGGCTCCCGCTGAATTGGA-3′; bottom, 5′-GGATTCCAATTCAGCGGGAGCCTCTCTTGAAGGCTCCCGCTGAATTGGA-3′) served as the shRNA control.
Cloning and generation of recombinant adeno-associated virus
The shRNA1336 and shRNA controls were subcloned into adeno-associated viral vector pAAV2-IRES-hrGFP. Virus production was performed with the AAV2 helper system (Stratagene, La Jolla, CA). Briefly, plasmid DNA (rAAV-shRNA1336-IRES-hrGFP plasmid plus the pRC vector encoding Rep and Cap proteins and the pHelper vector encoding adenoviral gene products) was used to transfect 293T cells at the 80% confluence stage. Cell lysates were collected 48 hr posttransfection and purified by CsCl density gradient centrifugation. Titers of rAAV-shRNA1336-IRES-hrGFP were determined by RT-PCR analysis by calculating the viral genome copy number.
5′-RACE analysis
5′-RACE (rapid amplification of cDNA ends) analysis was performed with a GeneRacer kit (Invitrogen). Briefly, total RNA (5 μg) was extracted from 3T3/AMCase cells treated with or without siRNA185 and siRNA1336 and ligated to GeneRacer Oligo. The resultant product was reverse-transcribed into cDNA, using a random primer. To detect cleavage products, PCR was performed with a 5′-nested primer and an AMCase-specific primer (5′-TTTCTGCGGCCGCATGGCATTAGGTTCATGGC-3′ and 5′-AACAAGCCCTGCTTGACAAT-3′, respectively) and then analyzed by electrophoresis and gene sequencing.
Animal immunization protocol
Animal studies were approved by the Institutional Animal Care and Use Committee of Chang Gung University (Taoyuan, Taiwan). Pathogen-free wild-type female BALB/cByJNarl mice from the National Laboratory Animal Center (Taipei, Taiwan) were used. Mice were 6–8 weeks old at the beginning of the experiment. Mice were injected intraperitoneally with chicken OVA (20 μg) complexed to alum or with normal saline alone. The process was repeated 3 days later. Thirteen days after the first immunization, animals were reinjected intraperitoneally with chicken OVA. Moreover, mice received by inhalation either an aerosol of OVA (2%, w/v) in normal saline or normal saline solution alone. Mice were thereafter challenged four times with OVA on days 16, 20, 23, and 27. On day 25, 1 × 1011 genome copies of rAAV-shRNA1336-IRES-hrGFP, rAAV-shRNAcontrol-IRES-hrGFP, rAAV-IRES-hrGFP (rAAV-mock), or normal saline solution was administered intratracheally. Mice were sacrificed on day 28.
Measurement of airway hyperresponsiveness
On day 28, 24 hr after the last challenge, airway responsiveness was measured in unrestrained animals by whole body plethysmography (Buxco Electronics, Wilmington, NC) (Lee et al., 2008a). Briefly, mice were administered aerosolized normal saline or methacholine at increasing concentrations (0, 25, and 50 mg/ml) for 3 min. Recordings were taken and averaged for 3 min after each nebulization. Airway reactivity was expressed as the mean enhanced pause (Penh).
Determination of OVA-specific IgE
Serum levels of OVA-specific IgE were measured by enzyme-linked immunosorbent assay (ELISA). Briefly, 96-well plates (Nunc MaxiSorp; Thermo Fisher Scientific, Waltham, MA) were coated with OVA (10 μg/ml) and blocked. After addition of serum samples at proper dilutions, biotin-conjugated rat anti-mouse IgE was added to the wells. The reaction was developed with streptavidin–horseradish peroxidase (HRP). Substrate solution was then added to each well and the plates were incubated for 30 min at room temperature in a dark room. After the addition of stop solution, the absorbance was read with an ELISA plate reader at 450 nm.
Assay of eotaxin and IL-13 levels
Eotaxin and IL-13 levels in BALF were assayed with commercially available ELISA kits (R&D Systems, Minneapolis, MN).
Western blot analysis
Cell and tissue samples for AMCase analysis were prepared with lysis buffer. After separation by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE), the samples were identified by incubation with rabbit anti-mouse AMCase antibodies and developed with an enhanced chemiluminescence (ECL) detection kit (GE Healthcare Life Sciences, Piscataway, NJ). A monoclonal antibody against β-actin was used as control.
AMCase activity
Chitinase bioactivity in BALF was measured with a fluorogenic substrate according to previous methodology (Zhu et al., 2004). Briefly, samples were incubated with 4-methylumbelliferyl β-
Lung histology
Lung tissues were removed and fixed with 4% formaldehyde. Paraffin sections, 10 μm thick, were stained with hematoxylin and eosin.
Immunohistochemistry
AMCase, hrGFP, and Ki67 expression in lung tissues was analyzed by immunohistochemistry (IHC). Lung tissue samples were prepared as 10-μm-thick paraffin sections and incubated with rabbit anti-AMCase, anti-hrGFP, or anti-Ki67 as the primary antibody. Sections were then counterstained with hematoxylin.
Statistical analysis
Results are presented as means ± SEM. Data were analyzed by nonparametric Mann–Whitney U test. A two-tailed p value less than 0.05 was considered statistically significant.
Results
Reduction of AMCase expression levels by RNA interference
RNA interference (RNAi) technology was used to evaluate whether the reduction of AMCase expression may affect asthma symptomatology in an OVA-sensitized mouse model. First, three shRNA expression vectors containing the mouse U6 promoter were constructed (pCI-shRNA185, pCI-shRNA897, and pCI-shRNA1336). Their ability to suppress AMCase expression was examined in a stable cell line overexpressing AMCase. All three constructs were able to reduce AMCase expression to various extents (Fig. 1a). AMCase expression was reduced by 85% with shRNA1336, by 50% with shRNA185, and by 20% with shRNA 897. Second, rAAV vectors encoding luciferase and shRNA1336, rAAV-shRNAcontrol-hrGFP and rAAV-shRNA1336-hrGFP (rAAV-shRNAcontrol and rAAV-shRNA1336, in brief) were cloned and investigated for their inhibitory effects on AMCase expression. Figure 1b shows that infection with rAAV-shRNA1336 significantly suppressed AMCase expression in the stable cell line between days 2 and 8. These findings indicate that rAAV-shRNA1336 had the better inhibitory effect on the expression of AMCase. To confirm that reduction of AMCase expression was mediated by specific cleavage of AMCase mRNA after transduction with rAAV-sh1336, 5′-RACE PCR was performed with stable cell lines overexpressing AMCase (see Supplementary Fig. 3 at
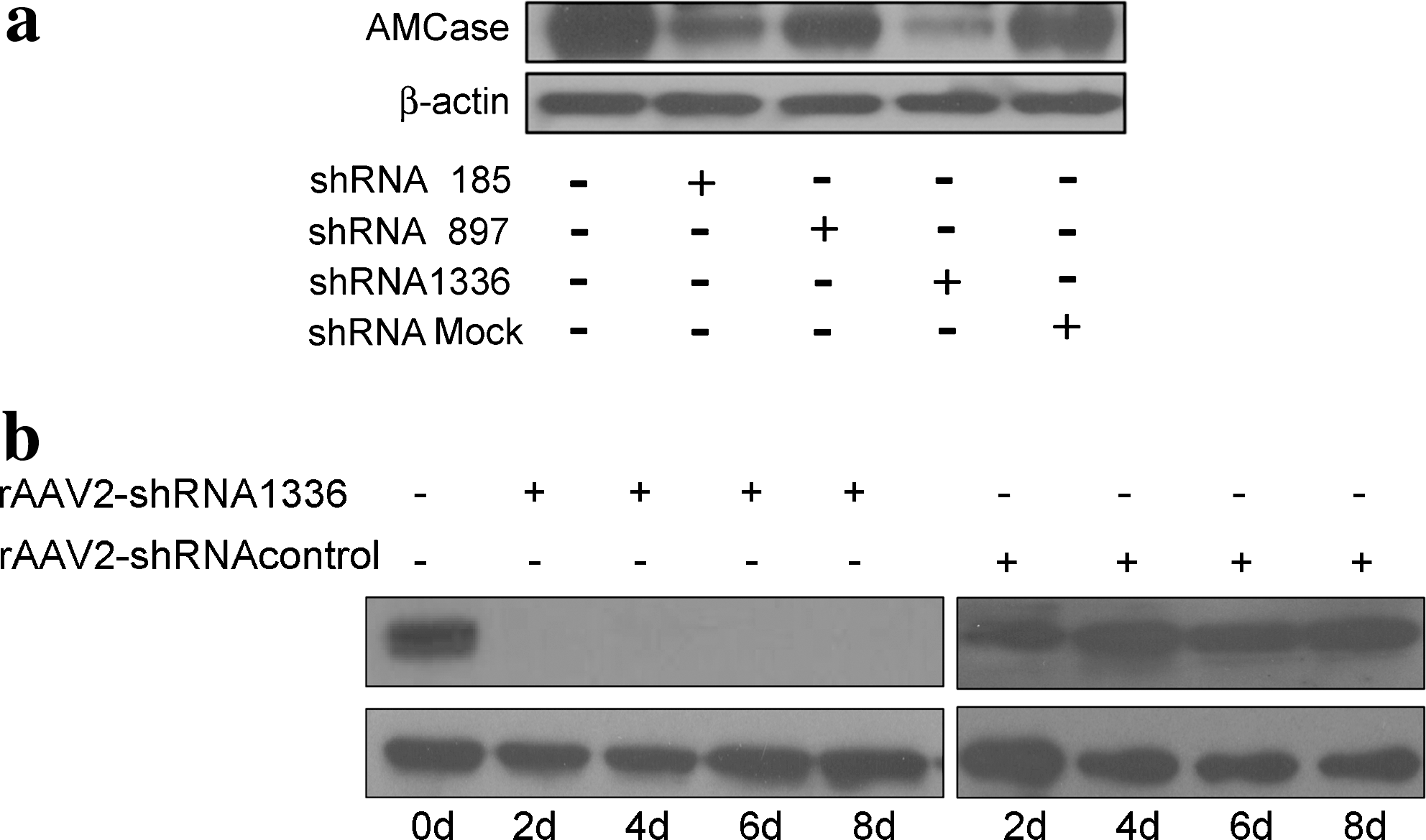
Inhibitory effects of pCI-shRNA and recombinant adeno-associated virus (rAAV)-mediated short hairpin RNA (shRNA) on acidic mammalian chitinase (AMCase) expression. (
Levels of AMCase in BALF and lungs of OVA-sensitized mice
To investigate whether elevated levels of AMCase expression were specifically located in the airways of OVA-sensitized mice, we harvested and analyzed lung, BALF, and peritoneal cells 24 hr after the last OVA challenge. Notably, 17- and 37-fold increases in AMCase mRNA level were found in the lung tissues and BALF of OVA-sensitized mice (Fig. 2a), respectively. No major differences in AMCase expression were seen in peritoneal cells from sensitized animal compared with control mice treated with normal saline. Similar findings were obtained when examining AMCase protein levels (Fig. 2b). Hence, hyperexpression of AMCase was confined chiefly to the airways. We also investigated whether the clinical symptoms in mice were associated with AMCase expression in the airways. As shown in Fig. 2c, airway hyperresponsiveness (AHR) was positively correlated with AMCase mRNA in lung cells as determined by real-time PCR. On the basis of using the 4-MU substrate system to measure AMCase activity in the airways, an association between AMCase activity and eosinophil infiltration was identified (Fig. 2d). In addition, we also found a positive correlation between AMCase expression and the total number of eosinophils in BALF (data not shown).
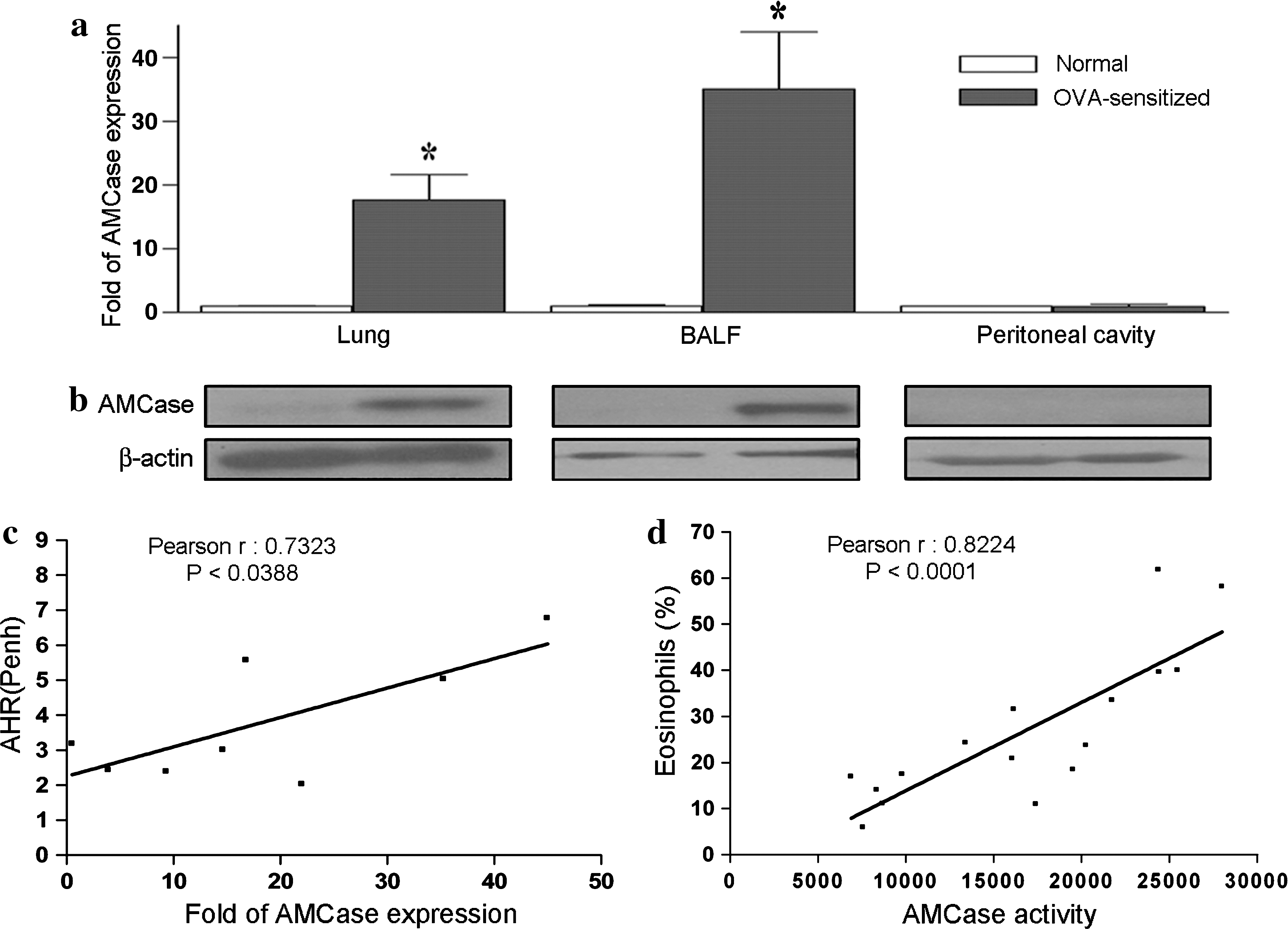
Quantification of AMCase expression and its correlation with asthmatic symptoms. Cell samples from lung tissues, bronchoalveolar lavage fluid (BALF), and the peritoneal cavity were harvested from normal saline controls (n = 5) and OVA-sensitized mice (n = 9), and their AMCase expression was quantified via (
Reduction of AHR and AMCase expression in the airways of OVA-sensitized mice after rAAV-shRNA1336 administration
We then examined whether rAAV-shRNA was able to suppress airway hyperresponsiveness and AMCase hyperexpression in our mouse model. Figure 3a depicts a schematic representation of the rAAV treatment protocols. Mice were sensitized with a mixture of OVA and alum, and then challenged by hyperexposure to aerosolized OVA. Three days after the intratracheal delivery of 1011 rAAV encoding LacZ (rAAV-LacZ), rAAV infection was evident in lung and BALF cells (data not shown). At this time, airway hyperresponsiveness was measured by methacholine challenge. Figure 3b shows that methacholine induced a significant increase in AHR in OVA-sensitized mice either with or without control viral infection (rAAV-mock and rAAV-shRNAcontrol). In contrast, sensitized mice receiving rAAV-shRNA1336 against AMCase showed significantly decreased Penh values in response to methacholine.
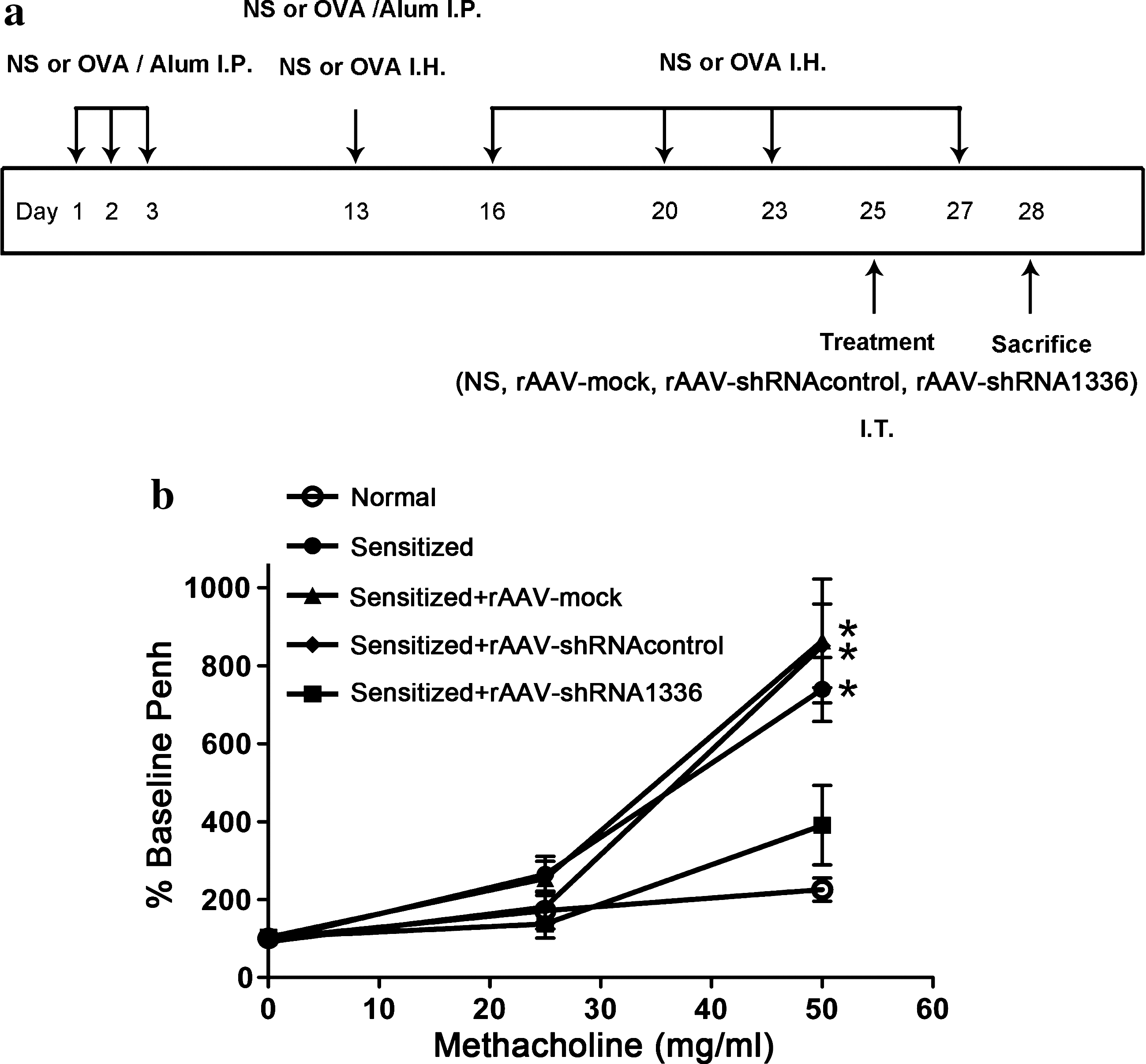
Reduction of airway hyperresponsiveness by treatment with rAAV-shRNA. (
We subsequently investigated whether such treatment resulted in reduced expression of AMCase. Figure 4a shows that AMCase expression was reduced by 2-fold in lungs and by 3-fold in BALF cells of mice treated with rAAV-shRNA1336 compared with mice receiving rAAV-shRNAcontrol or rAAV-mock. We also measured AMCase protein levels and activity in lung and BALF cells by immunohistochemical or enzymatic assay, using a commercial fluorescence system (4-MU). Figure 4b and c shows elevated expression of AMCase in the airway epithelial cells and BALF cells of OVA-sensitized animals receiving rAAV-shRNAcontrol or vehicle. In contrast, markedly reduced AMCase expression was seen in rAAV-shRNA1336-treated animals. Immunohistochemical analysis of the adjunct sections revealed that most of the airway cells such as epithelial cells expressed high levels of AMCase in sensitized animals (Fig. 4d). With the coordinated detection of hrGFP expression, these epithelial cells appeared to be the main target of AAV. Furthermore, by analyzing rAAV-shRNA1336-infected specimens, we actually found that high hrGFP-staining regions had lower levels of AMCase expression, indicating that administration of rAAV-shRNA1336 indeed reduced AMCase expression in lung tissues of OVA-sensitized mice. Moreover, 5′-RACE PCR was performed to confirm that reduction of AMCase was mediated by specific cleavage of AMCase mRNA after transduction with rAAV-shRNA1336, using RNA samples from the lungs of OVA-sensitized mice treated with rAAV-shRNA (Supplementary Fig. 3).

Suppression of AMCase in lung tissues and BALF cells of rAAV-shRNA-treated mice. In (
rAAV-shRNA1336 reduces immunopathological reactions in OVA-sensitized mice
Because treatment with rAAV-shRNA1336 significantly reduced AHR of OVA-sensitized mice, we further examined whether the other essential features of immunopathological allergic responses among these treated OVA-sensitized mice were affected. The OVA-specific IgE serum level was significantly lowered in sensitized mice treated with rAAV-shRNA1336 compared with animals treated with rAAV-shRNAcontrol (Fig. 5a). Moreover, infection with rAAV-shRNA1336 not only reduced eosinophil infiltration in BALF (Fig. 5b), but also decreased the infiltration of inflammatory cells into the lung tissues of sensitized mice (Fig. 5c). Altogether, these results indicate that asthmatic mice receiving rAAV-shRNA1336 showed a significant improvement in allergic inflammatory responses.

Reduction of immunopathological responses in rAAV-shRNA-treated mice. In (
Because leukocyte infiltration was found to be reduced after treatment with rAAV-shRNA, we further investigated whether reduction of AMCase activity could affect the expression of Th2-associated cytokines and chemokines. Specifically, we focused on IL-13, as its expression has been associated with AMCase expression, and eotaxin, which is an eosinophil-specific chemoattractant. As shown in Fig. 6a and b, concentrations of IL-13 and eotaxin were reduced in OVA-sensitized mice that received rAAV-shRNA1336 treatment. We subsequently focused on the expression of other Th2-associated cytokines (IL-4 and IL-5) and inflammatory chemokines (MCP-1 and MIP-1β) according to the various rAAV treatments. As shown in Fig. 6c, treatment with rAAV-shRNA1336 caused a reduction in expression of cytokines and chemokines in the lung, although the inhibition of IL-4 and IL-13 was less prominent compared with that of chemokines (eotaxin, MCP-1, and MIP-1β).

Expression of cytokines and chemokines in rAAV-shRNA-treated mice. (
Discussion
Experimental studies have suggested the potential use of RNAi for targeting asthma (Lee et al., 2008a; Lively et al., 2008). Our current work indicates that AMCase is a potential target. Our findings revealed that AMCase expression was positively associated with asthmatic symptoms, AHR, and eosinophil infiltration. rAAV-mediated shRNA production could effectively reduce AMCase expression in OVA-sensitized mice. Notably, reduction of AMCase expression correlated with a significant improvement of airway inflammation. Altogether, our findings imply that RNA interference targeting AMCase may lead to the development of a potential therapeutic strategy to ameliorate asthmatic symptoms.
AMCase has been hypothesized to associate with Th2-mediated inflammation through an IL-13-dependent mechanism (Zhu et al., 2004), as Zhu and colleagues found significant hyperexpression of AMCase in IL-13-transgenic mice. In fact, IL-13-null mice failed to express AMCase after an allergen challenge (Zhu et al., 2004). Zhu and colleagues showed that inhibition of AMCase activity with specific antibodies reduced the inflammatory response in BALF and lung tissues. However, these authors reported that reduction of AMCase activity did not lead to consequent changes in IL-4 and IL-13 expression. Here we adapted RNAi technology to further knock down AMCase expression, which led to the significant reduction of allergic responses. Importantly, we found that downregulation of AMCase expression was accompanied by suppression of IL-13 in BALF cells. Although lung IL-13 is not entirely suppressed by downregulation of AMCase activity, the existence of other molecular pathways controlling AMCase expression can be estimated.
Chemokines such as eotaxin, MCP-1, and MIP-1β have been implicated in airway inflammation (Zimmermann et al., 2003). Eotaxin acts as a chemoattractant for eosinophils in the lung after an allergen challenge (Pope et al., 2005b), whereas expression of MCP-1 and MIP-1β is enhanced in subjects with asthma. In the current study, we have shown that inhibition of AMCase in OVA-sensitized mice led to a significant reduction of eotaxin in BALF and to massive suppression of eotaxin, MCP-1, and MIP-1β in lung tissues. Inhibition of eotaxin-mediated AMCase suppression in turn decreased eosinophil infiltration. An overall improvement in the asthmatic response was achieved by rAAV-shRNA-mediated downregulation of AMCase. Whether this effect may be ascribed to an IL-13-dependent pathway is currently unclear; further studies are needed to shed more light on this issue. Accordingly, it has been demonstrated that neutralization of IL-13 abrogates the majority of eotaxin activity (Pope et al., 2005a).
Moreover, one study focusing on elaboration of the molecular mechanisms controlling AMCase and its effector responses showed that AMCase physically interacts with epidermal growth factor receptor (EGFR) (Hartl et al., 2008), which has been strongly implicated in the pathogenesis of Th2 tissue inflammation (Davies et al., 1999). In fact, it was demonstrated that EGFR signaling mediated allergen-induced CCL17 chemokine (CCL17/TARC, thymus and activation regulated chemokine) release by lung epithelial cells (Heijink et al., 2007). Indeed, recombinant AMCase was able to induce epithelial cells to produce CCL17, CCL2 (MCP-1), and CXCL8 (IL-8, KC) (Hartl et al., 2008). Indeed, in the current study, we found that reduction of MCP-1 in lung tissues was associated with inhibition of AMCase expression. This result indicates that AMCase plays a positive role in stimulating pulmonary epithelial cell chemokine production.
It is of key importance to determine whether AMCase is a potential therapeutic target, because AMCase is abundantly expressed not only in the airways but also in the gastrointestinal tract (Boot et al., 2001; Paoletti et al., 2007). IL-13 and GATA-3 are known to play a crucial role in the development of asthma (Huebner et al., 2008; Sel et al., 2008), and studies have shown that RNAi targeting IL-13 (Lively et al., 2008) and GATA-3 (Lee et al., 2008a) might serve as a novel therapeutic approach in the treatment of this condition. In the current study, we have shown that AMCase hyperexpression in sensitized mice is likely to be airway specific, being limited to the lung and alveolar macrophages. Local AMCase silencing in the airways could be achieved via an intratracheal administration of rAAV-shRNA.
Further investigation may be needed to determine whether the current strategy for targeting asthma delivers prophylactic and/or therapeutic effects; it could be beneficial if shRNA-mediated reduction of AMCase has prophylactic effects on asthma. Basically, in the current study, allergic responses including BALF eosinophilia as well as the elicitation of serum OVA-IgE appeared on day 18 after inhalation challenge with allergen OVA (see Supplementary Fig. 1 at
To overcome poor transfection rates in vivo, we used the AAV vector as a delivery tool for shRNA, in keeping with similar studies on bronchial diseases (Huang et al., 2008; Lee et al., 2008a). Zavorotinskaya and coworkers have previously compared the efficacy of a single intramuscular injection versus intratracheal administration of an rAAV vector carrying the IL-4 receptor antagonist in OVA-sensitized mice (Zavorotinskaya et al., 2003). The authors showed that intratracheal administration resulted in significant local effects in the airways, with no systemic or local adverse effects (Zavorotinskaya et al., 2003). Accordingly, the AAV vector achieved a better transfection rate not only in vivo, but also in vitro. In the present study, liposome-based transfection with pCI-shRNA resulted in approximately 85% AMCase inhibition, whereas almost total suppression was achieved with rAAV-shRNA. These data clearly indicate that rAAV may be regarded as an optimal transfection vector. To investigate the inhibitory effects of RNAi against AMCase, we have used stable cell lines overexpressing AMCase. It may be argued that RNA interference could exert only minor effects on AMCase expression in alveolar macrophages and lung epithelial cells, where AMCase expression is low. To address that issue, we have tested the inhibitory effects of rAAV-shRNA1336 on AMCase expression in lung epithelial cells (MLE-12), and a remarkable reduction of AMCase expression was also observed (data not shown). In addition, we also addressed whether the inhibitory effects of rAAV-shRNA on AMCase expression could be affected by cell division. As shown in Supplementary Fig. 2 (at
Gene transfer has emerged as a novel powerful therapeutic strategy and many reports have compared the advantages and limitations of various gene delivery vectors. In this study, we have used the rAAV vector as a tool for targeting asthma. Notably, adeno-associated viruses have been investigated in human gene therapy trials. Several studies have pointed out that long-term expression of transgenes may be achieved with little inflammatory response (McPhee et al., 2006; Moss et al., 2007), indicating that gene delivery via rAAV may represent the better option.
In summary, this is the first study to show the potential usefulness of silencing AMCase expression in asthma therapy. Further investigations are warranted to further understand the exact role of AMCase in the pathogenesis of this disabling allergic disorder.
Footnotes
Acknowledgments
This study was financially supported by Chang Gung University grant EMRPD140041 (Ministry of Education, Taiwan) and Chang Gung Memorial Hospital grant CMRPD150381 to C.R.S., and by National Science Council grants 96-2221-E-131-008 to C.L.L. and 95-2311-B-001-057-MY3 to C.N.S. The authors gratefully acknowledge the staff of the RNAi core laboratory at Chang Gung University for providing the siRNA backbone and construct instructions.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.