
Abstract
Despite some advances, patients with advanced renal cell carcinoma (RCC) cannot usually be cured. Alteration of the natural tropism of adenoviruses may permit more specific gene transfer to target tissues. The aim of this study was to use novel targeting moieties for adenoviral gene therapy of RCC. Previous work in rats suggested that use of Ad5/19p (Ad5 capsid with Ad19p fiber) with kidney vascular targeting moieties HTTHREP (HTT), HITSLLS (HIT), and APASLYN (APA) placed into the fiber knob might be useful for targeting kidney vasculature. Therefore, we sought to investigate the utility of Ad5/19p variants for gene delivery to human RCC cell lines, clinical samples, and orthotopic murine models of metastatic RCC. Six different human RCC cell lines were infected but only Ad5/19p-HIT showed increased transduction, and only in one cell line. Thus, we analyzed human normal and cancerous kidney specimens fresh from patients, which might better mimic the three-dimensional architecture of clinical tumors and found that Ad5/19p-HIT showed transduction levels similar to Ad5. In mice, we found that intraperitoneal and intravenous Ad5/19p-HIT transduced tumors at levels comparable to Ad5, and that intratumoral Ad5/19p-HIT was superior to Ad5. Liver tropism was significantly reduced in comparison with Ad5. Improvements in tumor-to-liver transduction ratios suggested that Ad5/19p-HIT may be promising for systemic gene delivery to kidney tumors.
Introduction
The success of gene therapy is dependent on the use of an efficient gene transfer system to allow the expression of the therapeutic gene in a specific organ, tissue, or cell. Unfortunately, intravenous administration of adenoviruses results in rapid blood clearance and liver transduction, which may be less than optimal for many gene therapy approaches (Alemany and Curiel, 2001; Isaka, 2006). Therefore, modification of the virus may be required for optimal systemic delivery.
Recombinant adenovirus serotype 5 (Ad5) is the most commonly used vector for gene therapy. Many viral modifications have been employed to increase the specificity and/or efficacy of Ad5 in vitro. Two main strategies have been used for targeting adenoviruses: (1) modification of the knob domain by adding different peptides into the C terminus or HI loop, and (2) pseudotyping with fibers derived from other adenoviral serotypes for binding receptors other than the coxsackievirus–adenovirus receptor (CAR) (Kanerva et al., 2002a,b, 2003; Bauerschmitz et al., 2006; Kangasniemi et al., 2006; Sarkioja et al., 2006; Ranki et al., 2007a,b). Examples of the former include C-terminal αv integrin-binding RGD motifs and heparan sulfate-binding polylysine residues (Wickham et al., 1997). There have also been reports (reviewed by Mizuguchi and Hayakawa, 2004) of inserting the peptides into the HI loop of the fiber knob, including those discovered by phage display library to show high affinity for vascular endothelial cells (Nicklin et al., 2000; Havenga et al., 2001, 2002), cancer cells (Nicklin et al., 2003), RGD in the HI loop (Dmitriev et al., 1998; Bauerschmitz et al., 2002), transferrin receptors (Xia et al., 2000), and vascular smooth muscle cells (Work et al., 2004).
Examples of the latter approach have featured fibers from all known adenoviral subgroups in the context of an Ad5 capsid. For example, altered vector tropism was reported by substitution of the Ad5 fiber protein into that of Ad3, Ad7, Ad11, Ad16, Ad17, Ad35, and others (reviewed by Mizuguchi and Hayakawa, 2004).
Almost all adenoviral subgroups use CAR-mediated entry into cells in vitro. Because of variable expression of CAR in many tumor types including renal cancer (Haviv et al., 2002; Guse et al., 2007a,b), many of the above-mentioned strategies have attempted to overcome dependency on CAR by switching to other cellular receptors. Some of the more promising approaches include RGD-4C or polylysine modification of the fiber, or serotype chimerism with Ad5/3 or Ad5/35 (Haviv et al., 2002; Kanerva et al., 2002a; Shayakhmetov et al., 2002; Smith et al., 2003; Guse et al., 2007a,b). However, most of the reported studies have used cell lines or primary patient samples in vitro, or intratumoral administration in vivo, while data on systemic delivery has been scarce.
Importantly, several publications suggest that the intravenous biodistribution of adenovirus is not determined by the fiber knob. Therefore, although knob modification may be able to enhance tumor transduction, it has not reduced uptake by nontarget organs such as the liver. Instead, the fiber shaft may play a key role in determining biodistribution (Haviv et al., 2002; Kanerva et al., 2002b; Shayakhmetov et al., 2002; Bayo-Puxan et al., 2006; Sarkioja et al., 2006; Ranki et al., 2007b). Therefore, approaches that have switched the complete fiber (instead of just the knob) or mutated relevant regions of the shaft, may be appealing for influencing the biodistribution of systemic virus. For example, Ad5 pseudotyped with the Ad35 fiber (including shaft) or interaction of Ad5 hexon with blood factors demonstrated less accumulation of the virus into the liver compared with Ad5 after intravenous administration (Shayakhmetov et al., 2002; Waddington et al., 2008). Also, mutation of the KKTK region of the fiber shaft has been reported to alter virus biodistribution (Alemany and Curiel, 2001; Bayo-Puxan et al., 2006).
Several gene transfer techniques have attempted to target specific renal structures, and promising approaches for treating renal diseases have been reported (Haviv et al., 2002; Isaka, 2006; Guse et al., 2007a,b). However, systemic delivery to the kidney or kidney tumors, while reducing delivery to nontarget organs, has not been reported heretofore.
In an earlier study in rats, we found that Ad19p results in less liver transduction than Ad5 (Denby et al., 2004). Ad19p belongs to subgroup D, which contains 32 different serotypes, most of which are rarely isolated from humans (Burmeister et al., 2004). Ad19p was reported to use sialic acids as the primary receptor (Arnberg et al., 2000a,b; Burmeister et al., 2004).
Using in vivo phage display in rats, we previously identified peptides HTTHREP, HITSLLS, and APASLYN that homed to the kidneys after intravenous injection (Denby et al., 2007). Furthermore, we engineered chimeric adenoviruses by inserting these peptides into the HI loop of Ad19p to create Ad5/19p-HTTHREP (Ad5/19p-HTT); Ad5/19p-HITSLLS (Ad5/19p-HIT), and Ad5/19p-APASLYN (Ad5/19p-APA). In vitro and in vivo experiments in rats followed, showing selective delivery to kidneys by Ad5/19p-HIT. However, the utility of these moieties for tumor transduction has not been studied previously. Also, other animals besides rats have not been studied thus far.
In this study, we hypothesized that Ad5/19p-HIT would result in effective transduction of human renal cancer substrates, including fresh patient samples, while retaining reduced hepatic transduction in the context of systemic delivery in three different mouse models of human cancer.
Materials and Methods
Kidney-derived cell lines and fresh tissues
Human renal clear cell carcinoma lines ACHN, 786-O, 769-P, Caki-2, angiomyolipoma Sv7tert, and leiomyosarcoma SK-UT-1B were obtained from the American Type Culture Collection (ATCC, Manassas, VA). All cell lines were maintained in the conditions recommended by the manufacturer.
Fresh normal and cancerous tumor samples were obtained with signed informed consent and ethics committee permission from a patient undergoing surgery at Helsinki University Central Hospital (Helsinki, Finland). She was a 62-year-old female with clear cell carcinoma of the kidney (histopathologic grade G3, primary tumor type pT3a). The complete kidney was removed and therefore normal kidney cells could be obtained from the same patient. For transportation, the tissues were kept on ice in RPMI 1640 with 20% fetal calf serum (FCS), penicillin (200 IU/ml), streptomycin (200 μl/ml), and
Adenoviruses
Generation and characterization of the replication-deficient adenoviruses used here (Table 1) have been reported (Denby et al., 2004, 2007).
From Denby et al. (2007).
Gene transfer assays
Cell line cells were plated in 24-well plates in triplicate and infected with viruses for 30 min in 200 μl of growth medium with 2% FCS. Cells were washed once and complete medium was added. After a 24-hr incubation at 37°C, β-galactosidase (β-Gal) (Tropix Galacto-Light Plus; Applied Biosystems, Foster City, CA) assays were done.
Human normal and tumor tissue samples were cut into 200-mg pieces, estimated to contain 50,000 cells each, and homogenized. However, the size of human explants (tumor and normal tissue) was not sufficient to test all of the differently modified adenoviruses, and hence we tested the capsid modification that showed the highest transduction in cell lines. Samples were washed twice with RPMI 1640 containing 2% FCS and infected with 5000 viral particles per cell in 500 μl of growth medium. After 1 hr of infection at 37°C, the medium was replaced with 2 ml of fresh growth medium. Tissue homogenates were lysed by adding 200 μl of lysis buffer for the luminescent β-Gal reporter system. Samples were freeze–thawed three times and supernatant was analyzed for β-Gal as described above.
Immunohistochemistry
Tissues were fixed in 4% formalin and paraffin blocks were made. Tissue sections (thickness, 4 μm) were prepared and incubated with rabbit anti-β-galactosidase polyclonal antibody at a dilution of 1:200 for 1 hr at room temperature (AB1211; Chemicon International/Millipore, Bedford, MA). Further, sections were incubated according to the manufacturer's instructions with LSAB2 system–HRP system (labeled streptavidin–biotin horseradish peroxidase) (K0673; Dako, Carpinteria, CA). Sections were counterstained with hematoxylin and dehydrated in ethanol, clarified in xylene, and sealed with Canada balsam. Pictures at magnifications of ×10, ×20, ×40, ×63, and ×100 were taken with an Axioplan2 microscope (Carl Zeiss, Thornwood, NY) equipped with an Axiocam (Carl Zeiss).
LacZ staining
Whole-mount tissues were fixed in fixing solution (25% glutaraldehyde, 100 mM EGTA [pH 7.3], 1 M MgCl2, 0.1 M phosphate buffer [pH 7.3]; Sigma-Aldrich, St. Louis, MO) and stained with 5-bromo-4-chloro-3-indolyl-β-
Quantitative real-time polymerase chain reaction
DNA was extracted from samples, using a QIAamp DNA mini kit (Qiagen, Valencia, CA). A LacZ quantification standard curve was generated from serial dilutions of each adenovirus preparation, using Power SYBR green (Applied Biosystems) with lacZ primers at 200 nmol/L (sense, 5′-ATC TGA CCA CCA GCG AAA TGG-3′; antisense, 5′-CAT CAG CAG GTG TAT CTG CCG-3′). Total DNA (100 ng) was amplified with the lacZ primers and products were quantified with a TaqMan 7900HT and SDS 2.3 analysis program (Applied Biosystems). The following reaction conditions were used: denaturation, 95°C for 10 min; amplification, 95°C for 15 sec, 54°C for 15 sec, 72°C for 30 sec (45 cycles); dissociation, 95°C for 15 sec, 60°C for 15 sec.
Animal experiments
All animal protocols were reviewed and approved by the Experimental Animal Committee of the University of Helsinki and the Provincial Government of Southern Finland. Mice were obtained from Taconic (Ejby, Denmark) at 4 to 5 weeks of age and quarantined for at least 1 week before the study. The health status of the mice was frequently monitored and as soon as any sign of pain or distress was evident they were killed.
For the subcutaneous tumor model, 5 × 106 ACHN cells were injected into nude and nonobese/severe combined immunodeficient (NOD/SCID) mice via both flanks. When tumors reached a volume of about 80 mm3 the mice were randomized into groups and injected with virus. Virus injections were performed on three consecutive days (days 1–3) intratumorally with 109 viral particles. Day 0 was the day before the first virus injection. Because some of the models were analyzed in parallel, we decided to do intravenous injection of 5 × 1010 viral particles on day 2 so that there would be an identical average time (48 hr) from virus injection to collection of tumors and organs in all models. Mice were killed 24 hr after the last intratumoral injection or 48 hr after the intravenous injection and selected organs including tumors were collected and stored at −80°C. The tissue samples were homogenized and lysed with cell culture lysis buffer, freeze–thawed, and supernatant was analyzed for LacZ activity with Tropix Galacto-Light Plus (Applied Biosystems) as described earlier (Kangasniemi et al., 2006).
Peritoneally metastatic disease was established in SCID mice by intraperitoneal injection of 107 ACHN cells per mouse. After 28 days mice were randomized and allocated into groups for intraperitoneal injection of 5 × 109 viral particles. Mice were killed 48 hr later. Selected organs and tumors were collected, stored at −80°C, and analyzed as described previously.
Results were normalized to protein content of the organs by DC protein assay (Bio-Rad, Hercules, CA).
SCID mice bearing intraperitoneal or subcutaneous tumors were injected intravenously at a dose of 5 × 1010 viral particles. Forty-eight hours after the intravenous injection selected organs including tumors were collected and stained for LacZ activity either by X-Gal staining or by immunohistochemistry.
Statistical analysis
To analyze variance in the quantitative real-time polymerase chain reaction (qPCR), the standard deviation was calculated on the basis of differences in the number of cycles needed for the fluorescence cutoff value, as reported (Denby et al., 2007). Variation in LacZ analysis was calculated as the standard error as reported (Kangasniemi et al., 2006). Mean tumor-to-liver ratios were first calculated for each animal and subsequently averaged. Variance is displayed as the standard error. To compare differences between groups, the Mann–Whitney test (SPSS 14.0; SPSS, Chicago, IL) was used and p values less than 0.05 were considered significant.
Results
Gene delivery of kidney-targeting adenoviruses in human kidney cancer cell lines
Our previous data suggested that Ad5/19p-HTT, Ad5/19p-HIT, and Ad5/19p-APA have kidney tropism in rats (Denby et al., 2007). However, a possible caveat is that the peptides were identified by in vivo phage display and enrichment in the same strain of rats and therefore the generalizability of the findings was unknown. Also, tropism to kidney tumors had not been assessed. Here we tested six different human renal cancer cell lines (ACHN, Sv7tert, Caki-2, 786-O, 769-P, and SK-UT-1B) representing three different histologies. Interestingly, only in the ACHN cell line did Ad5/19p or any of its capsid-modified variants achieve transduction levels comparable to that of Ad5 (Fig. 1).
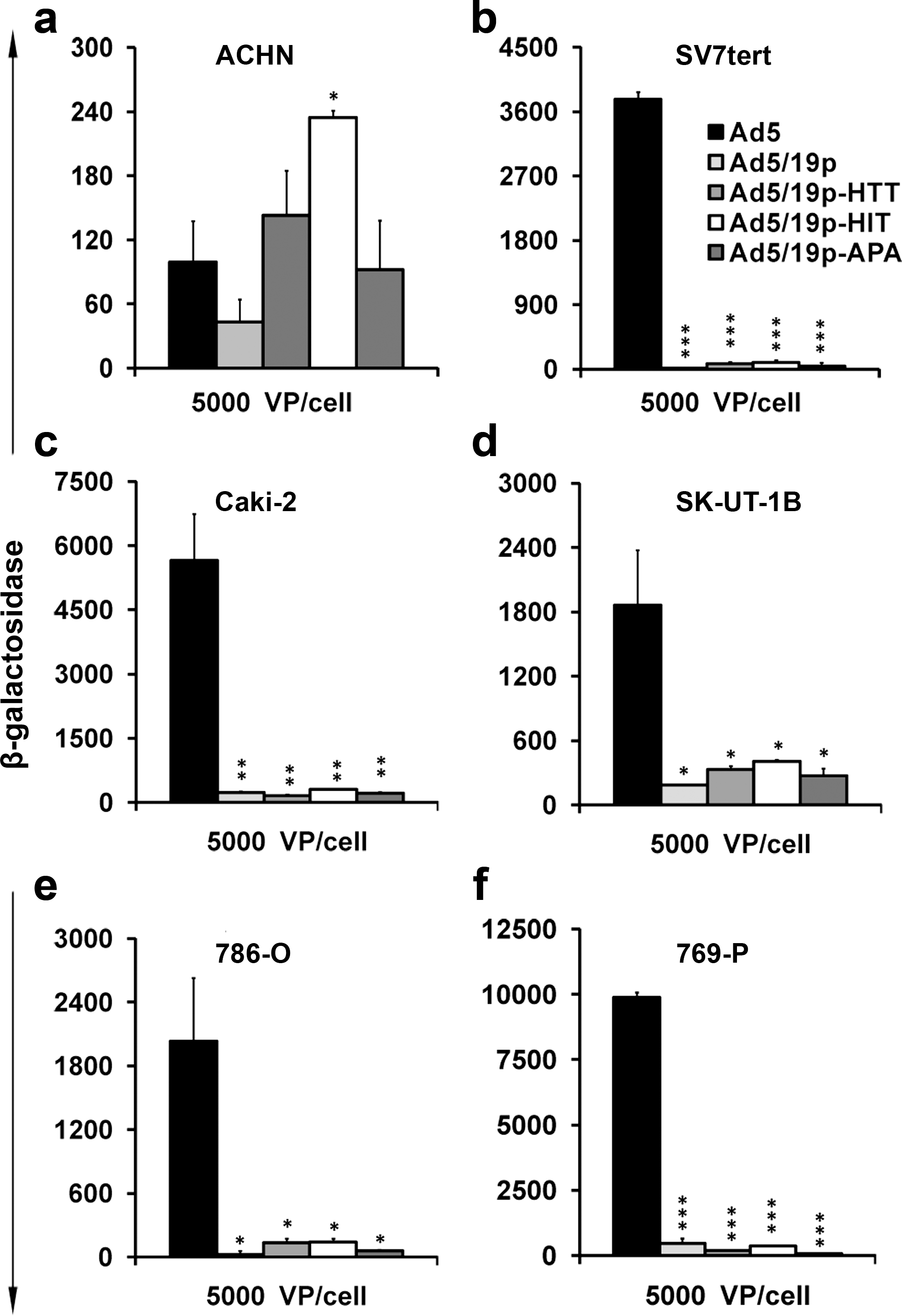
Transduction of human kidney cancer cell lines with capsid-modified viruses. Six renal carcinoma cell lines (ACHN, 786-O, 769-P, Caki-2, Sv7tert, and SK-UT-1B) were infected with replication-deficient β-galactosidase-expressing adenoviruses Ad5/19p-HTT, Ad5/19p-HIT, Ad5/19p-APA, isogenic control Ad5/19p, and Ad5. LacZ activity was measured as relative light units (RLU) 72 hr after infection. * p < 0.05; ** p < 0.01; *** p < 0.001. VP, viral particles.
Gene delivery of kidney-targeting adenoviruses in fresh human normal and tumor specimens
Cell lines are grown two-dimensionally and therefore might lack the epitopes that the peptides (originally identified in systemic administration in vivo) recognize. Therefore, we analyzed fresh human samples, which might retain some of the three-dimensional architecture present in vivo. Ad5/19p-HIT showed transduction similar to Ad5 in both tumor tissue (Fig. 2a) and normal tissue (Fig. 2b). To exclude interference due to transgene expression, the findings were confirmed by qPCR in tumor tissue (Fig. 2c) and normal kidney tissue (Fig. 2d).
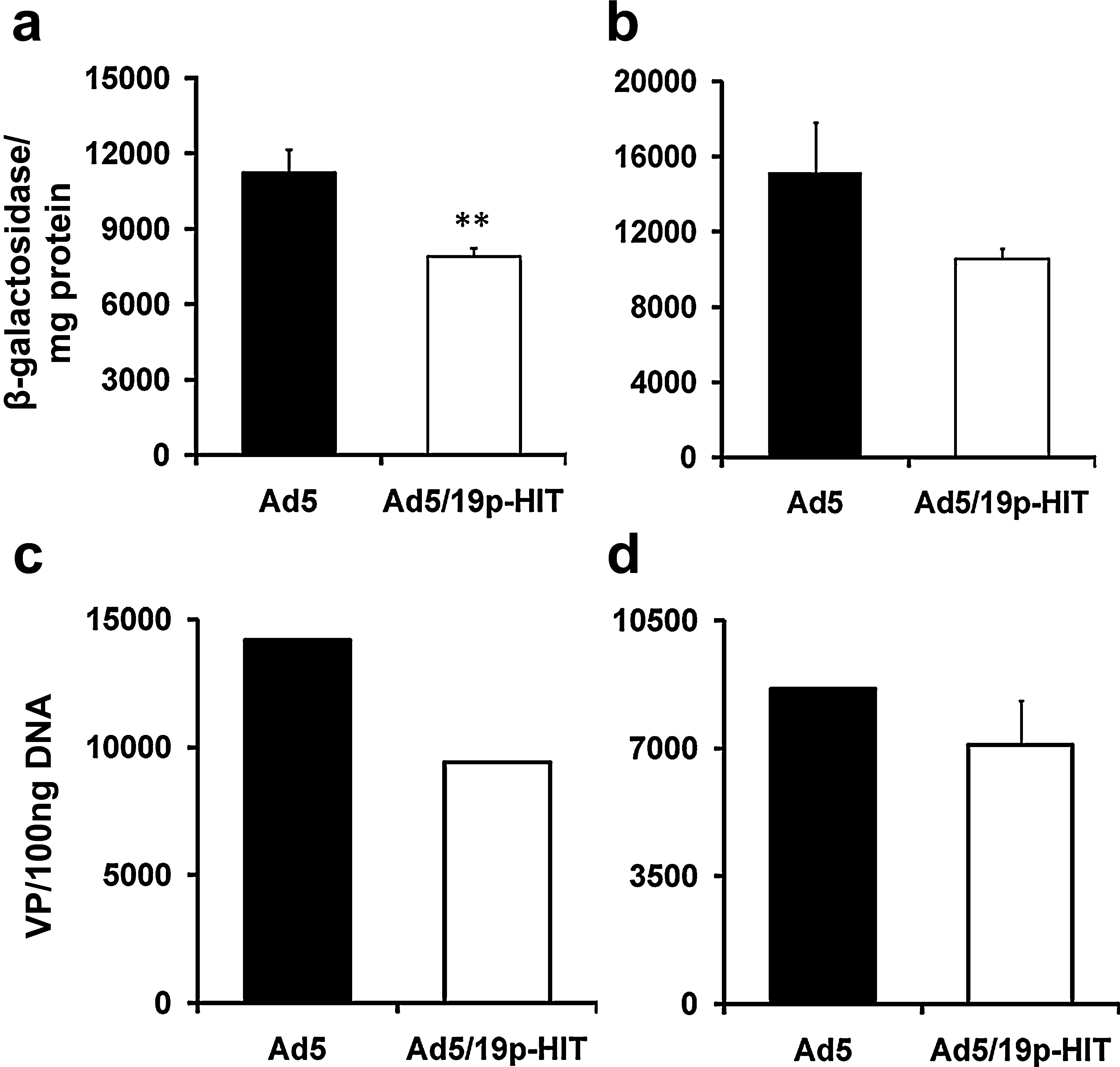
Transduction of fresh human tumor and normal tissues with capsid-modified viruses. Fresh human renal cell cancer (
Tumor transduction through various routes of virus administration in three murine models of human renal cell cancer
In vitro analyses may not fully capture the three-dimensional architecture of tumors, or their vascular supply. Thus, tumors were induced in immunodeficient mice and virus was given intravenously or intraperitoneally. In intravenously injected nude mice with subcutaneous tumors, the levels of LacZ activity with Ad5/19p-HIT and also other Ad5/19p-derived viruses were comparable to that of Ad5 (Fig. 3a). Similar results were observed after intraperitoneal administration to SCID mice bearing intraperitoneal tumors (Fig. 3b). When tumor cells were grown subcutaneously in NOD/SCID mice and virus was given intravenously, similar data were seen (Fig. 3d). Interestingly, when viruses were given intratumorally, Ad5/19p-HIT showed a 1000-fold increase in infectivity compared with Ad5 (Fig. 3c). To assess tumor transduction after intravenous administration, intraperitoneal tumors from SCID mice were collected 48 hr after virus administration and immunohistochemistry for LacZ (Fig. 3e) and qPCR (Fig. 3f) were performed. Significantly higher transduction (Fig. 3f) of the tumor was observed in the group of mice treated with Ad5/19p-HIT compared with Ad5 vector, supporting the hypothesis that virus bearing kidney-specific peptide enhances kidney-derived tumor transduction (Fig. 3e and f).
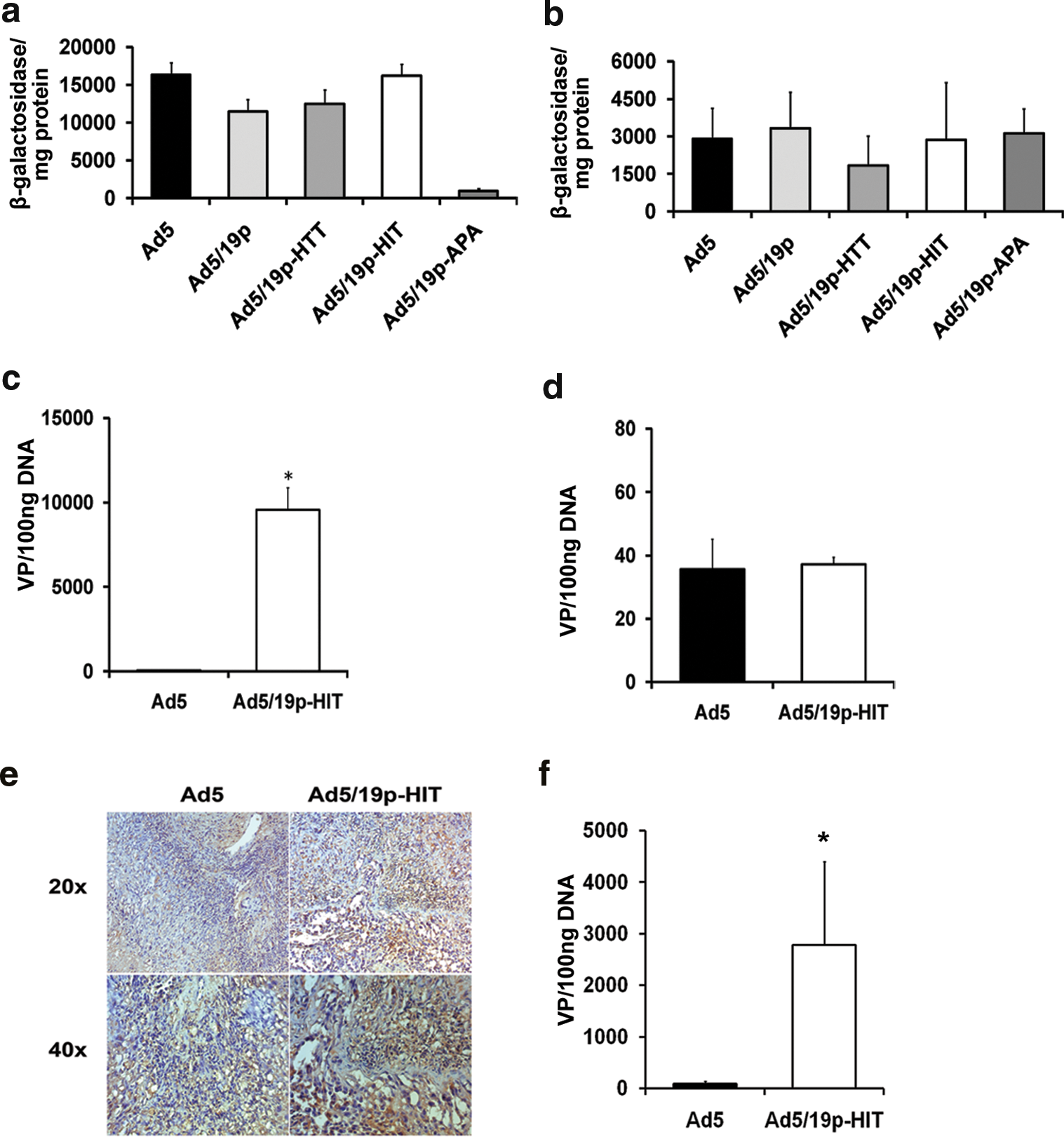
Tumor transduction in mouse models of human kidney cancer. (
Ad5/19p-HIT enhances kidney transduction after intravenous administration
Given that Ad5/19p-modified viruses have enhanced tropism to kidney in rats (Denby et al., 2004), we sought to investigate the biodistribution of such vectors after intravenous administration in mice. To this end, five groups of nude mice were injected intravenously with viruses bearing kidney-specific peptide and compared with Ad5-derived virus. Kidney was collected 48 hr later and β-Gal activity was measured. Although the virus bearing 19p-HIT peptide (Ad5/19p-HIT) showed higher kidney tropism in this mouse model, the overall expression from all the tested viruses was comparable to that of Ad5 (Fig. 4a). These data correlate with the results showed in Fig. 2. β-Gal expression, however, measures expression of the transgene in the analyzed tissue; it does not provide information at the cellular level. To assess whether viruses bearing kidney peptide had different tropism to different kidney microstructures, an additional experiment was performed. Two groups of SCID mice were injected intravenously with either Ad5 or Ad5/19p-HIT and 48 hr later the kidneys were collected and stained for LacZ. Interestingly, Ad5/19p-HIT showed more efficient penetration into the subcortical area compared with Ad5, suggesting a stronger affinity for proximal tubules (Fig. 4b). Moreover, these data suggest that transgene expression-associated postentry steps are more efficient with Ad5/19p-HIT than with Ad5.

Kidney transduction after intravenous injection of virus. (
In vivo liver detargeting with Ad5/19p-HIT in mice
In accordance with previous work in rats and in a human hepatoma line (Denby et al., 2004), we found significantly reduced liver uptake with Ad5/19p-HIT in comparison with Ad5 (Fig. 5). Comparable data were consistently observed among all of the various mouse models tested, suggesting the generalizability of the result in the context of mouse liver, regardless of route of administration. In nude mice (Fig. 5a) and NOD/SCID mice (Fig. 5c) injected intravenously, Ad5/19p-HIT resulted in significantly less liver uptake than Ad5 (p = 0.049 for nude mice and p = 0.014 for NOD/SCID mice). In SCID mice (Fig. 5b) injected intraperitoneally, Ad5/19p-HIT showed a significant decrease in liver uptake compared with Ad5 (p = 0.014). To complement the qPCR data, we performed an additional experiment in which SCID mice bearing intraperitoneal tumors were injected intravenously with Ad5 and Ad5/19p-HIT. Forty-eight hours after virus administration livers were collected and immunohistochemistry for LacZ was performed. Consistent with the qPCR data, Ad5/19p-HIT showed a significant reduction in liver transduction compared with Ad5 (Fig. 5d). Tumors were also analyzed, showing results consistent with previously described experiments in this paper (Fig. 3e).
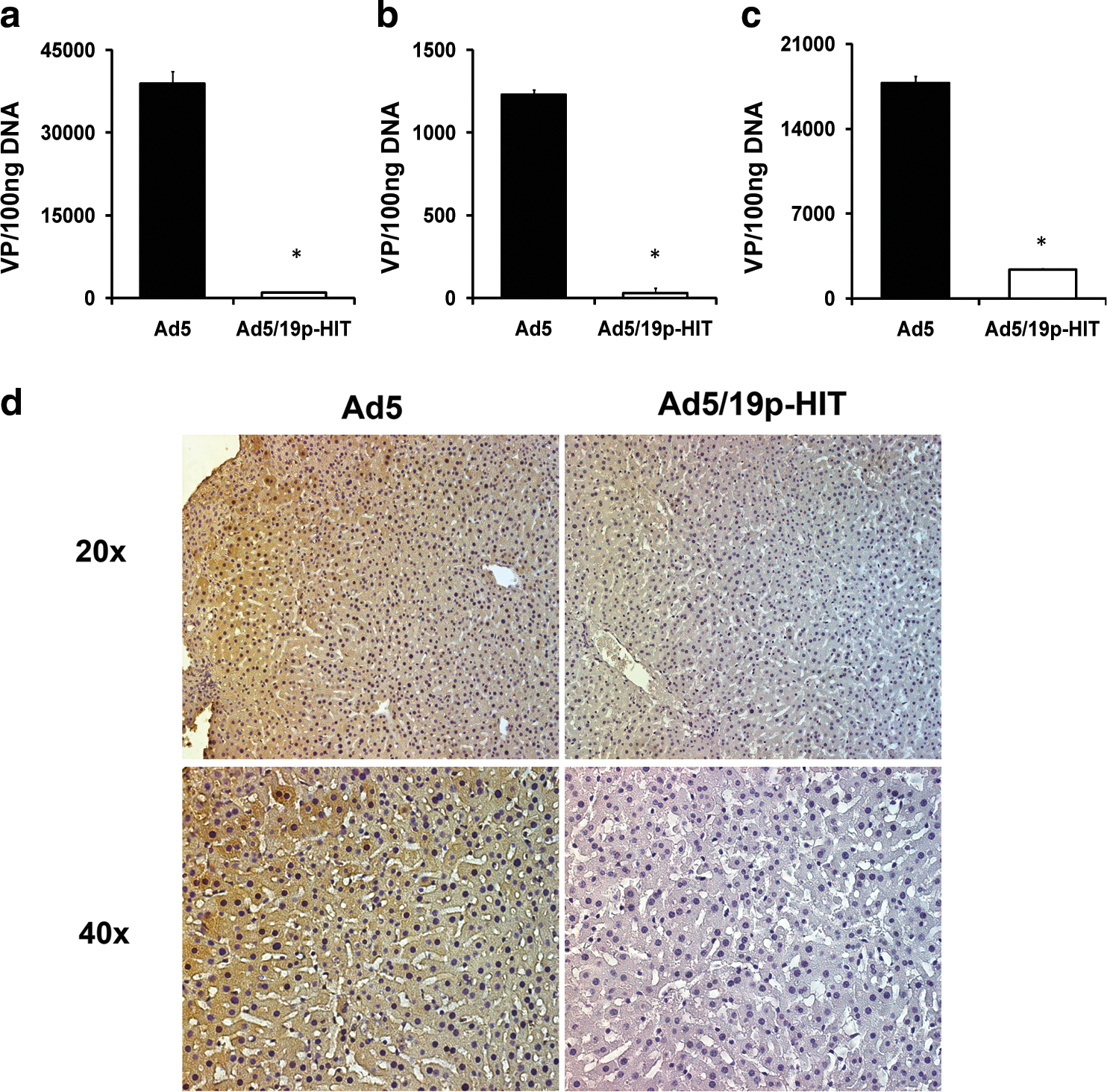
Liver transduction with Ad5 and Ad5/19p-HIT after intravenous or intraperitoneal injection. (
Tumor versus liver gene delivery in mice
Given the key role of the liver in determining adenoviral toxicity in both mice and humans (Alemany and Curiel, 2001; Isaka, 2006), the key attribute of systemic delivery of adenoviruses is the ratio of tumor-to-liver transduction. Intraperitoneal administration of Ad5/19p-HIT to mice bearing intraperitoneal tumors resulted in a similar rate of gene delivery to tumor versus liver (p = 0.286). Intratumoral administration of Ad5/19p-HIT in nude mice (p = 0.011) and NOD/SCID mice (p = 0.009) significantly increased the tumor-to-liver ratio of gene delivery when compared with Ad5 (Fig. 6a). Intravenous administration to tumor-bearing NOD/SCID mice resulted in higher transduction with Ad5/19p-HIT, although the difference was not statistically significant (Fig. 6a). Intravenous administration was also performed in SCID mice bearing intraperitoneal (Fig. 6b) and subcutaneous (Fig. 6c) tumors and the tumor-to-liver ratio was analyzed by immunohistochemistry. Ad5/19p-HIT showed significantly higher transduction (Fig. 6e) of the tumor coupled with lower transduction of the liver (Fig. 6d and e).
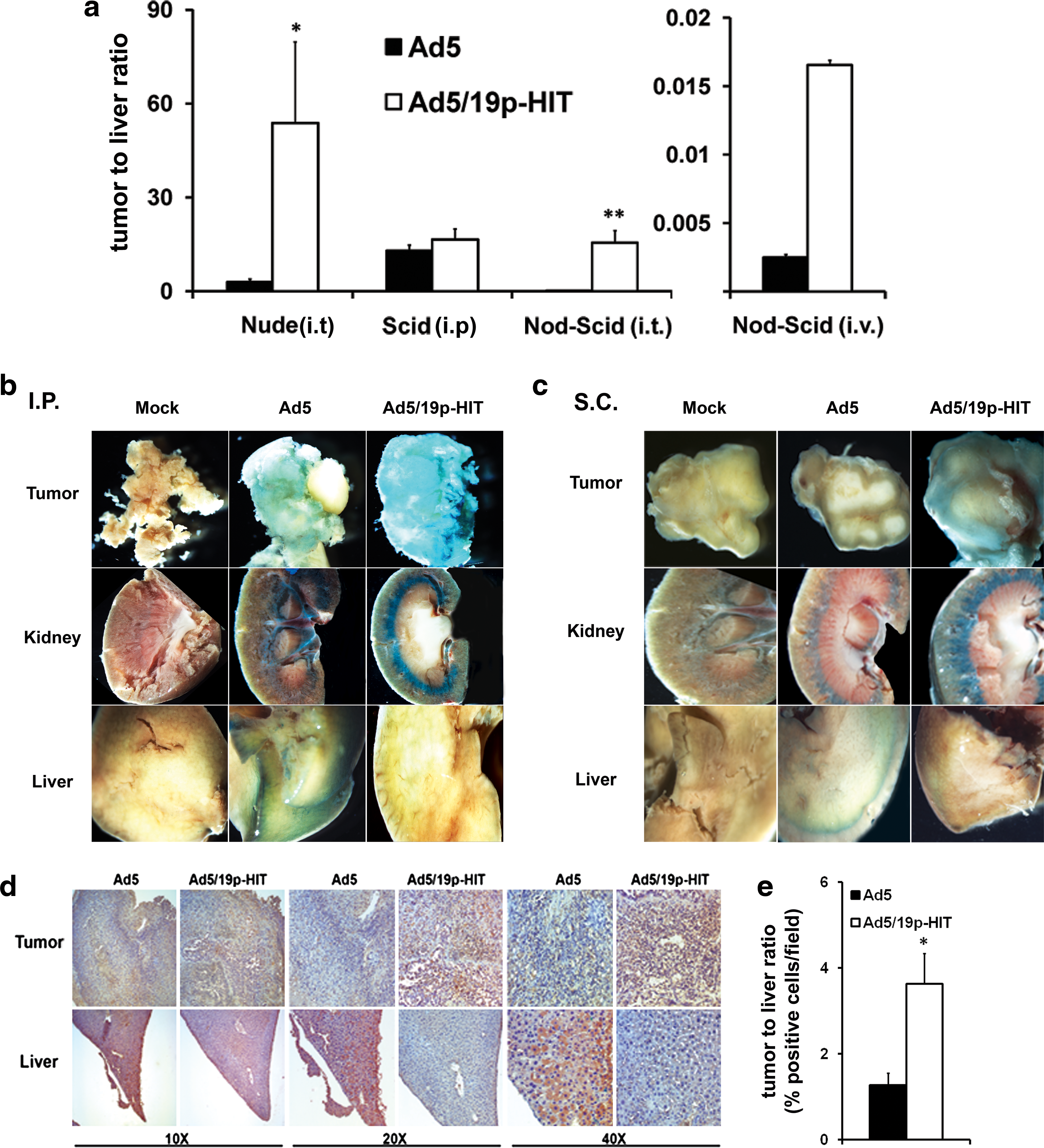
Ad5/19p-HIT increases the ratio of tumor-to-liver transduction in comparison with Ad5. (
Discussion
Patients with metastatic kidney cancers are in need of new treatment approaches. One developmental modality is gene transfer for a therapeutic purpose. When several different nonviral and viral approaches for kidney targeting were compared, the most efficient vector was adenovirus (Isaka, 2006). Subsequently, capsid-modified viruses and serotype-chimeric viruses have been shown to be more effective than Ad5 (Haviv et al., 2002; Guse et al., 2007a,b). We previously identified Ad19p as a useful approach for rat kidneys, and the virus was further improved by ligand incorporation into the HI loop (Denby et al., 2007). However, only one species of rat was studied, and the same rats had been used for discovery of the ligands. Therefore, the generalizability of the findings was not clear. Also, neither Ad19p nor its modified versions had been studied in the context of cancer.
With kidney cancer cell lines, Ad5/19p and Ad5/19p-HIT were useful only in one out of six lines. In contrast, in fresh normal and cancerous kidney-derived patient specimens, the infectivity of the fiber-modified viruses was similar to that of Ad5. One reason for these differences may relate to three-dimensional structure, which might be better retained in fresh clinical specimens in comparison with established cell lines. Of the Ad5/19p-related viruses, Ad5/19p-HIT emerged as the most appealing virus with regard to kidney or kidney tumor transduction.
Despite advances in engineering vectors for detargeting the liver, this nevertheless remains the main obstacle for systemic gene therapy. Systemic delivery, however, is of key importance in the treatment of metastatic disease, as most patients who die of cancer have widely disseminated disease. Previous work has shown that RGD or Ad5/3 fiber knob-modified adenoviruses can transduce kidney tumor cells more efficiently than Ad5 without increased liver toxicity (Haviv et al., 2002; Kanerva et al., 2002b; Guse et al., 2007a,b). Also, the systemic safety and utility of Ad5/3 have been demonstrated in some other tumor types (Kangasniemi et al., 2006; Sarkioja et al., 2006; Ranki et al., 2007a,b). Nevertheless, Ad5/3 does not result in reduced liver transduction.
Interestingly, we found significant liver detargeting with Ad5/19p-HIT in all mouse models reported. Our results are not in contrast with those reported by others regarding different capsid modifications for liver detargeting. Shayakhmetov et al. (2002) have reported that Ad5 pseudotyped with the Ad35 fiber showed decreased accumulation in the liver compared with Ad5 wild-type fiber after intravenous administration; reduced adenoviral interaction with CAR, coagulation factor IX, and complement protein C4 was speculated to play a role in the detargeting effect in the case of Ad5/35. Moreover, Denby et al. (2007) showed reduced factor X binding for Ad5/19p-derived adenoviruses compared with Ad5. Complete clarification of the reasons why Ad5/19p-derived adenoviruses show reduced liver tropism may require further studies. Nevertheless, one possibility is that the fiber modification might have modified the interaction with these proteins.
Haviv et al. (2002) have reported that adenovirus-based gene therapy for renal cancer requires retargeting to alternative cellular receptors. In their report they identified αvβ3 and αvβ5 as targets for renal cancer infection. In the same report, they focused on kidney tumors rather than liver detargeting and kidney targeting. Here, we found effective transduction of kidney tumors in three different mouse models and in addition to that we report an efficient adenovirus-based vector with tropism for the normal kidney. Interestingly, despite the comparable β-galactosidase expression between kidneys from mice treated with Ad5 or Ad5/19p-HIT, we observed different biodistributions for the tested vectors within the tissue. Further and more specific investigations are required to elucidate completely the biodistribution of Ad5/19p-derived adenoviruses within the kidney, and eventually these vectors might be used in future specifically for kidney targeting in the context of nonmalignant kidney diseases. Nevertheless, it may be potentially important for cancer therapy that we found higher tumor-to-liver transduction ratios with Ad5/19p-HIT than with Ad5.
In conclusion, Ad5/19p-HIT is promising for systemic and local delivery to human kidney tumors fresh from patients, and grown as xenografts in mice. Low liver transduction resulted in a favorable tumor-to-liver transduction ratio. Further work is needed to more intricately characterize the toxicity profile in animals as a prelude to possible clinical testing in humans. Eventually, increases in the rate of tumor-to-liver gene delivery might improve the safety and efficacy of adenoviral cancer gene therapy in patients.
Footnotes
Acknowledgments
The authors thank Eerika Karli, Aila Karioja-Kallio, and Roxana Ola for expert assistance. This study was supported by the European Research Council, Finnish Cancer Society, EU FP6 APOTHERAPY and THERADPOX, Helsinki Biomedical Graduate School, HUCH Research Funds (EVO), Sigrid Juselius Foundation, Academy of Finland, Biocentrum Helsinki and Emil Aaltonen Foundation, Finnish Cultural Foundation, and Kidney Research UK. Akseli Hemminki is K. Albin Johansson Research Professor of the Foundation for the Finnish Cancer Institute.
Author Disclosure Statement
There are no competing financial interests.