
Abstract
The delivery of genes or viruses via liposomes is a common approach used to enhance delivery efficiency. In the current study, to enhance delivery efficiency, proteoliposomes (PLs) containing adenovirus (Ad) were synthesized with dimyristoylphosphatidylcholine (DMPC), cholesterol, and apolipoprotein A-I (apoA-I). Wildtype apoA-I (WT) or V156K-apoA-I (V156K) was then used as an apolipoprotein to compare the structural and functional differences of the PLs. The particle diameter of V156K-PL-Ad was slightly larger than that of WT-PL-Ad, based on native gel electrophoresis. V156K showed more rapid phospholipid bilayer formation than did the WT, based on DMPC clearance. In addition, V156K exhibited maximal fluorescence that was more blue than that of WT in the PL state. Moreover, isothermal denaturation in response to the addition of guanidine hydrochloride (Gnd-HCl) revealed that V156K was more resistant, with no denaturation until 3 M Gnd-HCl was added. In addition, electron microscopy revealed that the viral particles were well associated with PL particles, which had a discoidal structure and were shaped like rouleaux. In addition, treatment of Ad in the PL state showed enhanced green fluorescent protein (GFP) expression when compared with treatment with Ad alone or with DMPC-Ad in hepatoma and brain glioma cells. Cells treated with WT-PL-Ad and V156K-PL-Ad showed approximately 50% more GFP expression than cells treated with Ad alone or with DMPC-Ad after 24 hr of incubation at 37°C, indicating that viral stability was highly increased in the PL state. Furthermore, V156K-PL-Ad showed the highest expression of GFP in adult zebrafish (9 weeks old) at 5 days postinjection (10.5- and 3.8-fold more GFP expressed than by Ad only and DMPC-Ad, respectively). In conclusion, the efficiency of viral delivery and the stability of the virus were significantly enhanced when PLs containing apoA-I were used in cellular and zebrafish models.
Introduction
To determine whether the advantages of viral and nonviral vectors could be combined, Ad and liposome were combined in the current study by creating a proteoliposome (PL) containing Ad as a new gene therapy. PL, which is also known as reconstituted high-density lipoprotein (rHDL), has been developed for the delivery of small molecules, including hydrophobic biomolecules, such as those used to produce anticancer drugs (Lacko et al, 2002; Lou et al., 2005) and antifungal drugs (Oda et al., 2006). The results of studies evaluating the use of rHDLs have indicated that they may be efficient drug delivery vehicles for therapeutic agents because of the ability of peripheral cells to acquire HDL core components.
In the current study, we created a new PL comprising phospholipid, cholesterol, wildtype (WT) apolipoprotein A-I (apoA-I) or its variant (V156K), and Ad in an attempt to maximize viral stability, infection ability, and gene delivery efficiency, while minimizing unwanted side effects. WT human apoA-I is composed of a single polypeptide chain of 243 amino acids and a principal protein component of HDL that is known to perform a crucial role in the reverse cholesterol transport pathway through its antioxidant and antiinflammatory activities (Barter et al., 2004; Ansell et al., 2005). A point mutant of apoA-I, V156K, showed unique structural and functional properties in vitro (Han et al., 2005), and V156K-rHDL has been shown to exert antioxidant effects in hypercholesterolemic C57BL/6 mice (Cho et al., 2006, 2007) and antiatherosclerotic activity in apoE-deficient mice (Cho and Kim, 2009).
The efficiency of delivery and expression was tested in human cells (hepatoma and brain glioma cells) and in a vertebrate animal model using zebrafish (Danio rerio).
Materials and Methods
Materials
Dimyristoylphosphatidylcholine (DMPC, 1 g; cat. no. 850345C) and cholesterol (cat. no. C8667) were purchased from Avanti Polar Lipids (Alabaster, AL) and Sigma-Aldrich (St. Louis, MO), respectively. Guanidine hydrochloride (Gnd-HCl) was purchased from Sigma-Aldrich (cat. no. 177253).
Purification of wild-type and V156K apoA-I
WT and V156K apoA-I were expressed in an Escherichia coli expression system, and then purified by Ni2+ column chromatography as previously described (Han et al., 2005; Cho et al., 2006). Protein purity was assessed by sodium dodecyl sulfate–20% polyacrylamide gel electrophoresis (SDS–PAGE), using a PhastSystem (GE Healthcare Bio-Sciences, Uppsala, Sweden).
Generation of Ad expressing green fluorescent protein
Replication-incompetent Ad encoding green fluorescence protein (GFP) was propagated in 293 cells, purified by CsCl density purification, dissolved in storage buffer (10 mM Tris, 4% sucrose, and 2 mM MgCl2), and stored at −80°C until use. The number of viral particles was calculated from measurements of optical density at 260 nm (OD260), where 1 absorbency unit is equivalent to 1012 viral particles (VPs) per milliliter.
Synthesis of proteoliposome with Ad
PLs containing DMPC, free cholesterol (FC), and apoA-I were synthesized by sonication, using an initial DMPC:FC:apoA-I molar ratio of 200:9:1. DMPC and FC, which had been dissolved in organic solvent, were dried under N2 gas at ambient temperature. The dried lipid mixture was dispersed into 0.5 ml of phosphate-buffered saline (PBS) and vortexed at 25°C to allow the formation of multilamellar vesicles (MLVs). To produce single unilamellar vesicles (SUVs), MLVs were sonicated three times at level 3 for 10 min, using a probe-type sonicator with a 10-min interval between treatments (model vcx130; Sonics & Materials, Newton, CT). The samples were then incubated at 25°C for 30 min, after which the SUV solution was centrifuged at 5000 × g for 5 min to remove any titanium or other precipitated by-products. Apolipoprotein (final concentration, 1 mg/ml in PBS), WT or V156K, was then added to the supernatant, which was subsequently incubated for an additional 1 hr to allow PL formation. After incubation, 10 μl of Ad stock solution (final concentration, 4.2 × 1010 particles/ml of PL) was added to the PL, which was then incubated at room temperature for 24 hr to allow association of the PL and Ad. The association of PL and Ad was monitored by observing the absorbance at 325 nm, using the method described by Pownall and colleagues (1978), with slight modifications. Measurements were initiated after the addition of DMPC and then monitored at 325 nm every 2 min, using a DU800 spectrophotometer (Beckman Coulter, Fullerton, CA) equipped with a thermo-controlled cuvette holder adjusted to 24.5°C (MultiTemp III; GE Healthcare Bio-Sciences).
Characterization of PL-Ad
The size distributions of PL-Ad were determined by 8–25% native polyacrylamide gradient gel electrophoresis (PAGGE; PhastSystem), and comparing the results with those of standard globular proteins (GE Healthcare Bio-Sciences) as described previously (Jonas et al., 1990). The number of apoA-I molecules per PL particle, as well as the self-association properties of the lipid-free proteins, were determined by cross-linking with bis(sulfosuccinimidyl)suberate (BS3), using the method described by Staros (1982). The products of the reaction were then analyzed by SDS–PAGE on precast 8–25% gradient gels (GE Healthcare Bio-Sciences).
Characterization of tryptophan fluorescence and isothermal denaturation
Wavelengths of the maximal fluorescence (WMF) of tryptophan residues in WT and V156K apoA-I were determined from uncorrected spectra obtained with an LS 55 spectrofluorometer (PerkinElmer Life and Analytical Sciences, Waltham, MA) in conjunction with WinLab software package 4.00 (PerkinElmer Life and Analytical Sciences) and a 1-cm path-length Suprasil quartz cuvette (Fisher Scientific, Pittsburgh, PA). Briefly, samples were excited at 295 nm to avoid tyrosine fluorescence, and the emission spectra were then scanned from 305 to 400 nm at room temperature. The effects of the addition of Gnd-HCl on the secondary structure of apoA-I in the lipid-bound state were then monitored by observing the WMF.
Electron microscopy
Transmission electron microscopy (TEM) was performed with a model H-7600 electron microscope (Hitachi, Ibaraki, Japan) operating at 80 kV. Briefly, PL-Ad was negatively stained with 1% sodium phosphotungstate (PTA, pH 7.4), with a final apolipoprotein concentration of 0.3 mg/ml. Five microliters of the PL-Ad suspension was then blotted with filter paper and immediately replaced with 5 μl of 1% PTA. After a few seconds, the stained PL fraction was blotted onto a Formvar carbon-coated 300 mesh copper grid and air-dried. The shape and size of the PLs were then determined on the basis of a TEM photograph taken at a magnification of × 40,000.
Cell cultures
A human embryonic kidney cell line that expresses the adenoviral E1 region (HEK293), a hepatocellular carcinoma cell line (Hep3B), and a brain glioma cell line (U343) were purchased from the American Type Culture Collection (ATCC, Manassas, VA). Hep3B cells were cultured in modified Eagle's medium (MEM; GIBCO-BRL, Grand Island, NY) supplemented with 10% fetal bovine serum (FBS; GIBCO-BRL), glutamine (2 mM), penicillin (100 IU/ml), and streptomycin (50 μg/ml). U343 and 293 cells were cultured in Dulbecco's modified Eagle's medium (DMEM; GIBCO-BRL) supplemented with 10% FBS, glutamine (2 mM), penicillin (100 IU/ml), and streptomycin (50 μg/ml). All cell lines were maintained at 37°C in a humidified atmosphere with 5% CO2.
Flow cytometric analysis for GFP expression
Hep3B cells (1 × 105 cells per well) and U343 cells (5 × 104 cells per well) were seeded in 24-well plates. After 12 hr, medium containing 10% FBS was removed and the cells were washed three times with PBS. Serum-free medium containing PL-Ad at 1 × 108 VP (Hep3B) and 5 × 107 VP (U343) was then added to the cells, which were subsequently incubated for 8 hr. Next, the medium was replaced with 10% FBS–medium and the cells were incubated for an additional 40 hr in the CO2 incubator. The cells were then observed by fluorescence microscopy (BX51; Olympus, Tokyo, Japan), using the MetaMorph imaging system (Molecular Devices, Sunnyvale, CA). To quantify the level of GFP expression in the cells that had been infected with PL-Ad, the cells were raised, using cell dissociation solution (Sigma-Aldrich), and then washed three times with PBS. After washing, 400 μl of PBS was added and flow cytometric analysis was conducted with a FACScan equipped with CellQuest software (BD Biosciences, San Jose, CA).
Zebrafish
Zebrafish and embryos were maintained according to standard protocols. The maintenance of zebrafish and procedures using zebrafish were approved by the Committee of Animal Care and Use of Yeungnam University (Gyeongsan, Korea). The fish were maintained in a system cage at 28°C during treatment under a 10:14 hr light cycle.
Microinjection of Ad into zebrafish
Nine-week-old zebrafish (n = 5 in each group) were individually microinjected, using a pneumatic PicoPump (PV820; World Precision Instruments, Sarasota, FL) equipped with a magnetic manipulator (MM33; Kantec, Bensenville, IL) with a pulled microcapillary pipette-using device (PC-10; Narishige, Tokyo, Japan). To minimize bias, injection was done at the same position on the tails of the zebrafish. Filter-sterilized solutions containing the same number of Ad (1.8 × 108 VP in 100 nl) in each vehicle were injected into zebrafish after anesthetization with tricaine (final concentration, 0.002%) (A5040; Sigma-Aldrich). After injection, live zebrafish were observed under a stereomicroscope (SMZ 168; Motic, Hong Kong) and photographed with a Motic cam2300 CCD camera. The fluorescence signal of GFP was detected with an Eclipse TE2000 microscope (Nikon, Tokyo, Japan). The mean size of the GFP-expressing area was calculated and quantified by computer-assisted morphometry, using Image-Pro Plus software (version 4.5.1.22; Media Cybernetics, Bethesda, MD).
Miscellaneous
Protein concentration was determined by Lowry protein assay, as modified by Markwell and colleagues (1978), or with Bradford assay reagent (Bio-Rad, Seoul, Korea) with bovine serum albumin as a standard. Determination of phosphorus was carried out by the method described by Chen and colleagues (1956) using a phosphate standard (24-2495; Sigma-Aldrich) based on the reduction of phosphomolybdate. FC content was determined by an enzymatic and fluorometric method (Heider and Boyett, 1978) with cholesterol oxidase and horseradish peroxidase.
Statistical analysis
All data are expressed as means ± SD, based on at least three independent experiments with duplicate samples. In Figs. 5 and 6, data comparisons between the two groups were assessed by Student t test, using SPSS (version 14.0; SPSS, Chicago, IL).
Results
Characterization of PL-Ad
WT or V156K apoA-I was expressed and purified to at least 95% purity by densitometric analysis and SDS–PAGE (Fig. 1A). Even though no major difference was observed between WT-PL-Ad and V156K-PL-Ad after synthesis, V156K-PL-Ad had a slightly larger particle size and a more homogeneous particle population than did WT-PL-Ad (Fig. 1B). Furthermore, the main particle diameter distribution of V156K-PL-Ad was >17 nm, as indicated by the arrowhead on the 4–15% native polyacrylamide gradient gel (Fig. 1B), whereas WT-PL-Ad showed two major bands at 17 and 12 nm. Because apoA-I in the lipid-bound state results in multimerization via a cross-linking reaction with a cross-linker, such as BS3 (Han et al., 2005), the apoA-I in PLs was cross-linked with BS3 and then electrophoresed (SDS–PAGE). As shown in Fig. 1C, WT and V156K apoA-I existed as a trimer or tetramer in PLs from the 8–25% SDS–polyacrylamide gradient gel, indicating that three or four molecules of apoA-I protein were required to form PLs. Although the same molar ratio was used for synthesis, the final molar composition of the two PLs was different, with the ratio of V156K-PL-Ad being 230 ± 18:9:1 (DMPC:cholesterol:apoA-I) and that of WT-PL-virus being 174 ± 15:8:1 (DMPC:cholesterol:apoA-I), as shown in Table 1. This finding indicates that V156K-PL-Ad had a greater affinity for DMPC and was therefore able to form larger particles.

Electrophoresis patterns of proapoA-I in proteoliposomes (PLs). (
Abbreviations: Ad, adenovirus; chol, cholesterol; DMPC, dimyristoylphosphatidylcholine; GGE, gradient gel electrophoresis; PL, proteoliposome; V156K, V156K apolipoprotein A-I (apoA-I); WMF, wavelength maximal fluorescence; WT, wild-type apoA-I.
Value in parentheses indicates WMF in the lipid-free state.
Ability to clear DMPC
To compare the phospholipid-binding affinity of WT and V156K in the presence of Ad particles (4.2 × 1010 particles/ml), each apolipoprotein and Ad stock solution were added to DMPC MLVs and the absorbance (325 nm) was then monitored. As shown in Fig. 2, V156K-PL-Ad had better phospholipid clearance activity, with 25% of the DMPC cleared by 60 min, whereas with WT-PL-Ad only 18% of the phospholipid was cleared during the same incubation time. The enhanced clearance of DMPC in the presence of V156K and Ad indicates better solubility of V156K-PL-Ad.

Kinetics of the interaction of proapoA-I and V156K with DMPC single unilamellar vesicles. The absorbance at 325 nm was monitored at 24.5°C at 2-min intervals. Circles, DMPC alone; triangles, DMPC plus Ad; squares, DMPC plus WT apoA-I plus Ad; DMPC plus V156K apoA-I plus Ad.
Wavelength maximal fluorescence
Among the four tryptophan residues in the sequence of apoA-I, Trp-108 is responsible for the fluorescence intensity. As shown in Table 1, the maximal fluorescence of both WT and V156K in the lipid-free state was observed at approximately 349 nm (WMF), which is in good agreement with the results of a previous study (Han et al., 2005). However, in the lipid-bound state, the WMF of V156K-PL-Ad shifted into the blue range (approximately 344 nm), whereas the WMF of the WT-PL-Ad was approximately 346 nm. This result indicates that Trp-108 is more exposed in nonpolar environments.
To compare the change in secondary structure, an isothermal denaturation method using concentrations of Gnd-HCl that increased from 0 to 6 M was used and the change in WMF was monitored, as shown in Fig. 3. Although no difference in WMF was observed when >3 M Gnd-HCl was added, at lower concentrations (<2 M Gnd-HCl) V156K-PL-Ad exhibited a 1- to 2-nm shift toward the blue range when its WMF was compared with that of WT-PL-Ad. This result indicates that Trp-108 in V156K-PL-Ad was less exposed in the aqueous phase than was Trp-108 in WT-PL-Ad, which might influence the stability of the secondary structure of the two proteins.
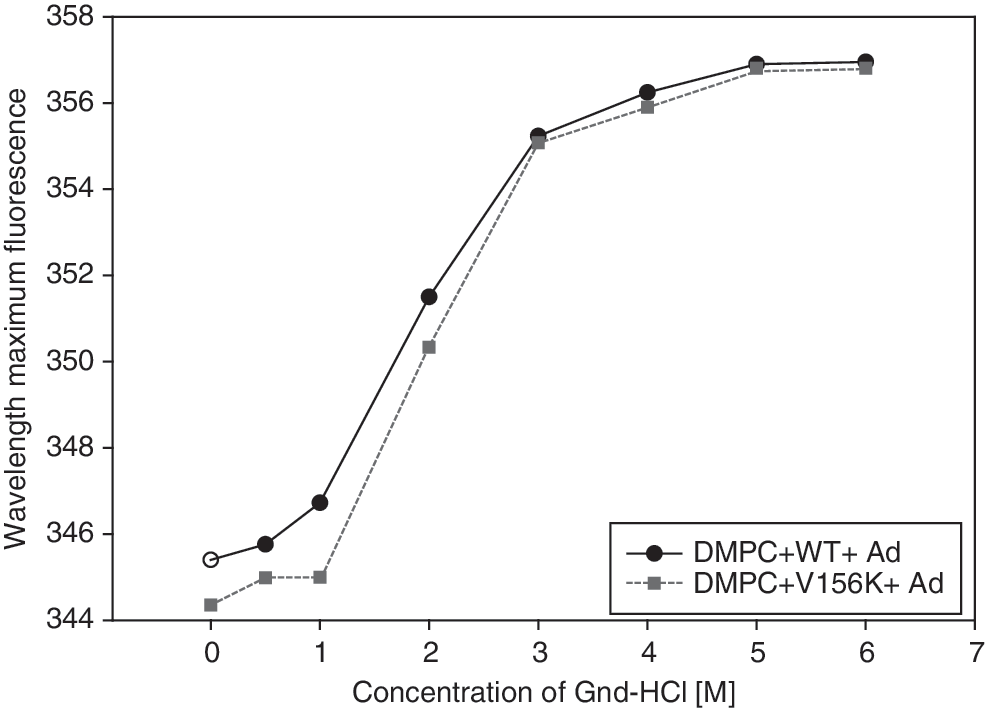
Denaturation of proapoA-I in the lipid-bound state (i.e., the PL state). Denaturation was monitored by observation of the wavelength maximal fluorescence as a function of Gnd-HCl concentration after equilibration for 72 hr at 25°C.
Electron microscopy
As shown in Fig. 4, TEM revealed that the PLs had a discoidal structure and were shaped like rouleaux, and that the adenoviral particles were well associated with the PLs. This association of Ad and PLs may stimulate an increase in the solubility of the virus and its stability. In addition, observation at the same magnification (×40,000) revealed that V156K-PL-Ad had a slightly larger diameter (approximately 17–22 nm) than WT-PL-Ad (12–17 nm). This result is consistent with the size determination based on native gradient gel electrophoresis (Fig. 1B).

Electron microscopy of negatively stained PLs containing adenovirus. Both micrographs are shown at an original magnification of × 40,000.
Enhanced Ad-GFP expression in PL-Ad
After Hep3B cells were treated with PL-Ad at 1000 VP/cell for 48 hr, the fluorescence signal was detected by fluorescence-activated cell sorting (FACS). There was no significant difference in the expression of GFP by cells treated with Ad alone or with DMPC-Ad (Fig. 5A). However, when cells were treated with Ad that was in WT-PLs or V156K-PLs, the expression of GFP was significantly enhanced (38–39%; Fig. 5), indicating that Ad delivery was enhanced by PLs containing either WT or V156K. The results of the FACS analysis were confirmed by fluorescence microscopy, as shown in Fig. 5B.
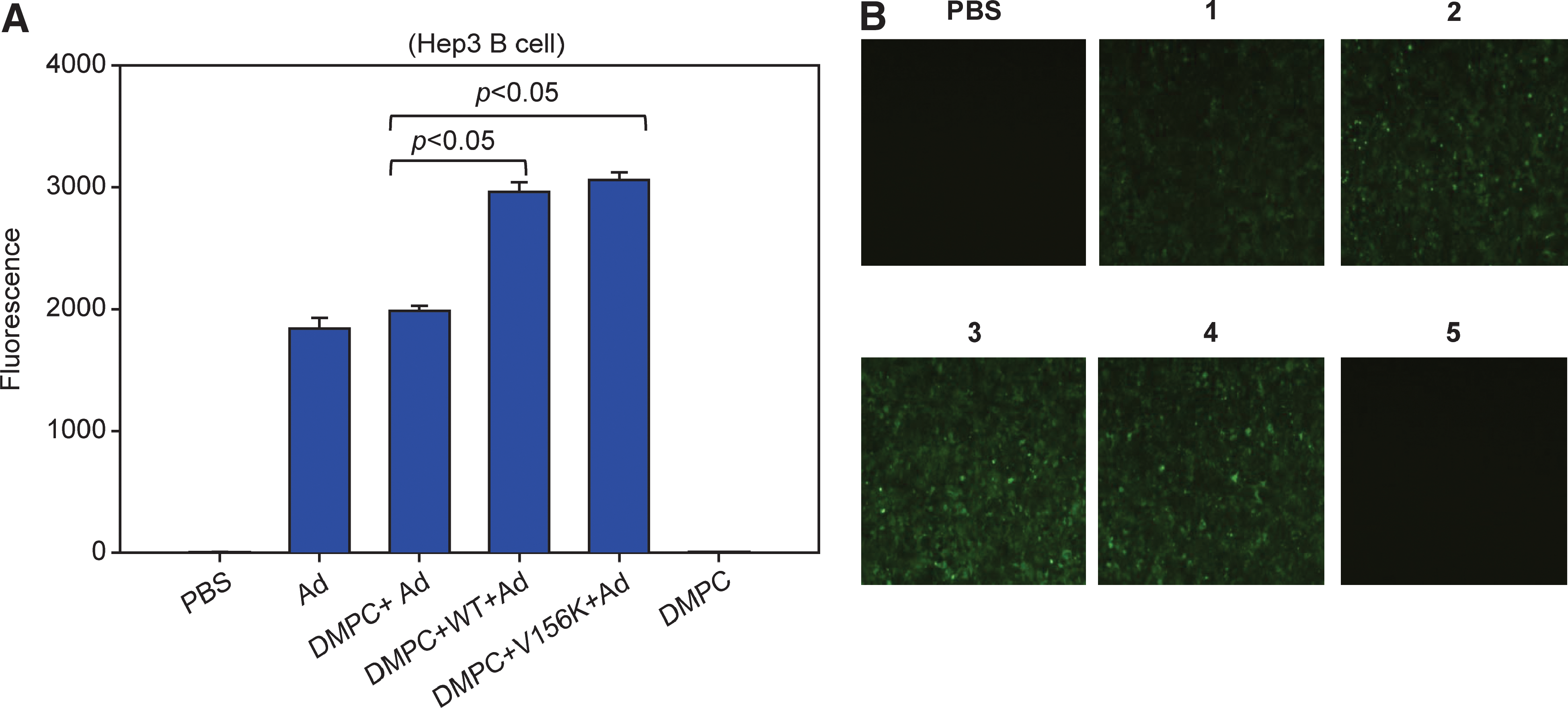
The difference in the level of GFP expression in Hep3B cells depends on adenovirus delivery. (
Similar results were observed when U343 brain glioma cells were treated with the same dose (Fig. 6). When U343 brain glioma cells were used, delivery of Ad by WT-PLs and V156K-PLs resulted in 50 and 48% enhancement of GFP expression, respectively, when compared with cells that were treated with Ad alone. There was no significant difference observed between cells that were treated with Ad alone (Fig. 6, column 1) and those treated with DMPC-cholesterol-Ad (column 2), indicating that apoA-I is an essential component of the enhanced gene delivery.

The difference in the level of GFP expression in U343 cells depends on adenovirus delivery. (
Increased adenoviral stability in PLs
To compare viral stability at ambient temperature, each PL-Ad was incubated for 24 hr at 4, 25, and 37°C. After incubation, U343 cells were treated with PL-Ad (1000 VP/cell) and allowed to incubate for 48 hr at 37°C. As shown in Fig. 7, the infective ability of Ad alone or DMPC-Ad that had been incubated at 25 and 37°C was approximately 2-fold less than that of samples that had been incubated at 4°C (Fig. 7, column groups 1 and 2). When viruses were in the PL state, however, the infective ability was not decreased at 25 and 37°C (Fig. 7, column groups 3 and 4). V156K-PL-Ad showed a slightly higher level of gene expression than WT-PL-Ad; this result suggests that viral stability and infective ability were enhanced by association with V156K-DMPC-cholesterol.
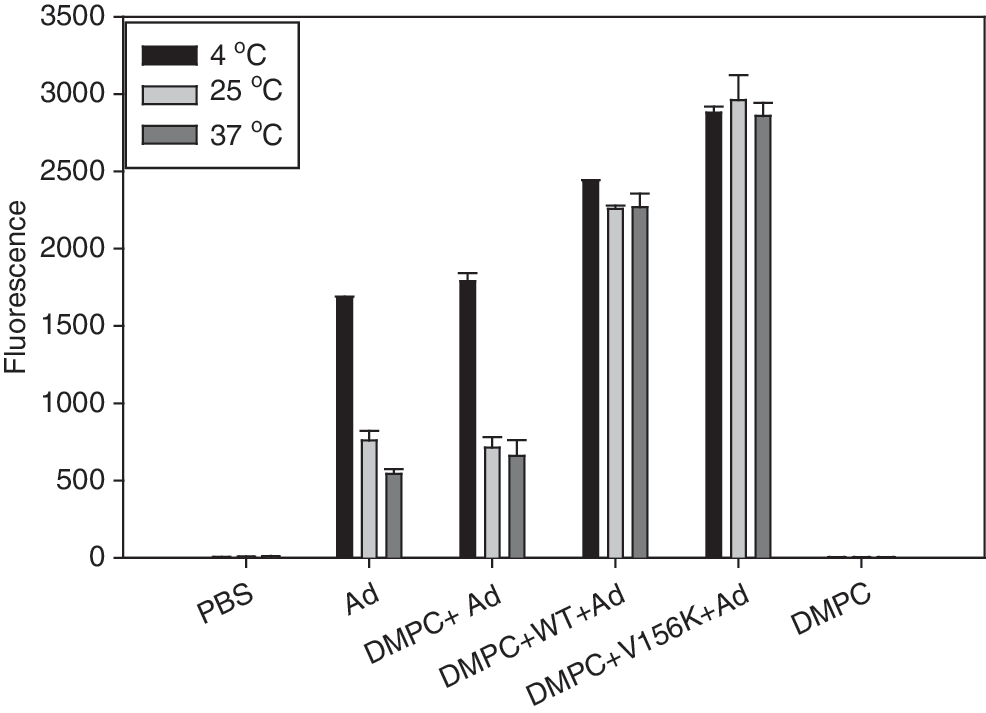
Protective effect of PLs against temperature exposure of adenovirus. After 24 hr of incubation at the designated temperatures, levels of GFP expression in cells were compared by FACS. PBS, treated with phosphate-buffered saline; 1, Ad alone; 2, DMPC + Ad; 3, DMPC + WT + Ad (WT-PL-Ad); 4, DMPC + V156K + Ad (V156K-PL-Ad); 5, DMPC alone.
GFP expression in zebrafish
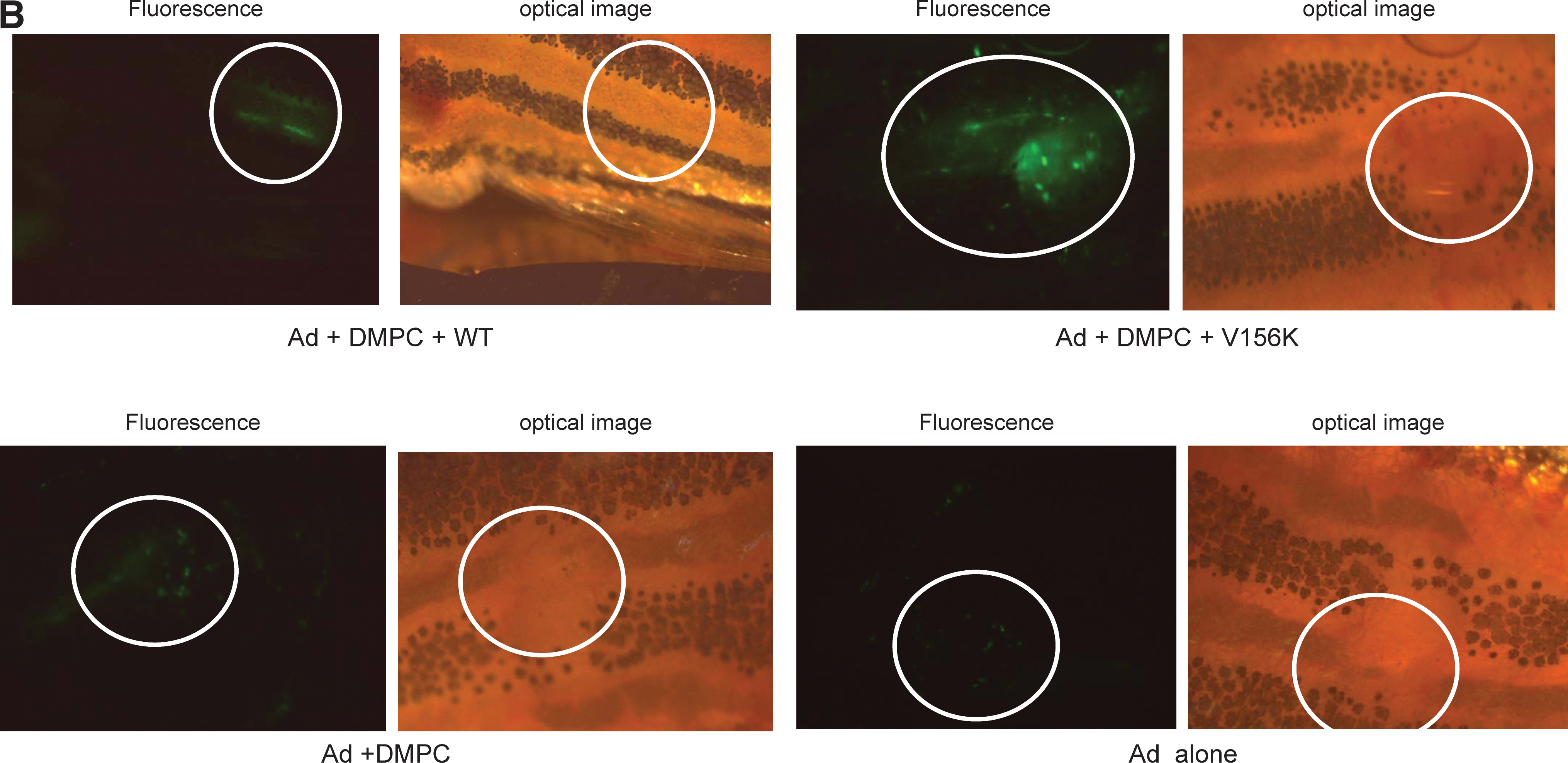
All zebrafish that received adenoviral injection tolerated the injection and survived. At 2 days postinjection, WT-PL-Ad and V156K-PL-Ad showed similar levels of expression, that is, 2.9- and 4.5-fold higher than with DMPC-Ad and Ad alone, respectively (Fig. 8A). In all cases of adenoviral injection, the efficiency of expression reached its maximal level at 5 days postinjection (Fig. 8A), indicating that viral gene expression in zebrafish is slower than in human cell lines (Figs. 5 and 6). Regardless of the type of protein, PL-Ad had a much stronger intensity of GFP expression than DMPC-Ad or Ad only (Fig. 8). Of the two types of PL-Ad tested, V156K-PL-Ad showed the highest level of expression (1.9-fold higher than WT-PL-Ad). At 7 days postinjection, the intensity of expression was diminished in all zebrafish.
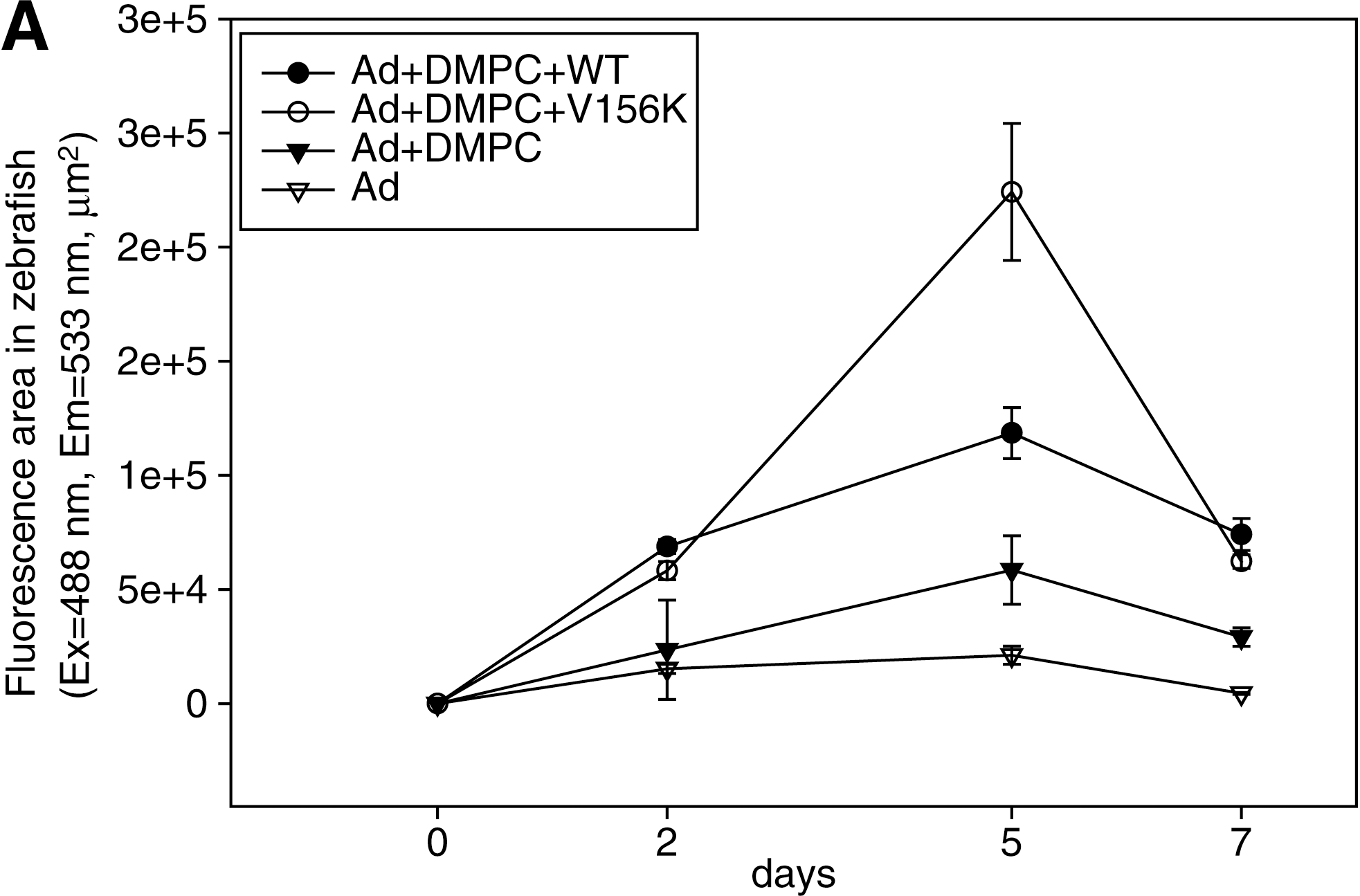
Comparison of the level of GFP expression in adult zebrafish. The same number of Ad particles (1.8 × 108 VP in 100 nl) in each vehicle was injected into zebrafish (n = 5 in each group). (
Discussion
The results of the current study demonstrate that Ad can be formulated in PLs as a novel tool for targeted gene delivery, which has a variety of therapeutic applications. The use of adenoviral gene therapy has been limited by many factors, such as toxicity and antigenicity, which may cause inflammation. Such side effects result in a need to lower the dose to maximize the efficiency of gene delivery. Although there have been several reports of liposomes being used as a delivery vehicle for Ad, this study has focused on minimizing the side effects of Ad (Steel et al., 2005). The current study is the first report involving the use of PLs as an adenoviral delivery vehicle. When apoA-I or its variants were not present, the mixture of DMPC-Ad lost its enhanced delivery efficiency in vitro and in vivo (Figs. 5 –8). ApoA-I is the principal protein of HDL in the blood, and it has been reported that WT apoA-I and V156K apoA-I are strong antioxidants (Cho et al., 2006, 2007). Cho and Jonas (2000) reported that Val-156 was a key amino acid regarding the structure and functions of apoA-I. Recombinant V156K mutant behaved differently from the wild type in both lipid-free and lipid-bound states in vitro and in vivo (Cho et al., 2006, 2007). V156K exhibited enhanced potent antioxidant and antiinflammatory activity, and an antiatherosclerotic effect (Cho and Kim, 2009) with different movement of the hinge domain of apoA-I as compared with wild type (Han et al., 2005). In addition, numerous clinical observations have shown that higher apoA-I serum concentrations result in lower cardiovascular risk (Walldius and Jungner, 2007). Furthermore, DMPC has been found to have profound antiatherosclerotic activity in mice, even though it is a synthetic phospholipid (Navab et al., 2003). In addition, Navab and colleagues (2003) reported that the oral administration of DMPC raises the HDL-cholesterol and apoA-I levels, improves HDL function, and causes regression in apoE-deficient mice. The addition of cholesterol brings order to phospholipids, thereby making the membranes of cells more rigid. Therefore, the addition of cholesterol may also strengthen the structure of the PL bilayer. Because of the beneficial properties of each component, PL-Ad is expected to have antioxidant and antiatherosclerotic activities, as well as anticancer activity. Although 1,2-dioleoyl-3-trimethylammonium propane (DOTAP), didecyldimethylammonium bromide (DDAB), and 1,2-dioleoylphosphatidylethanolamine (DOPE) have been widely used to prepare complexes of cationic liposomes/plasmid DNA or adenoviral vectors (Steel et al., 2007), the application has been limited because such complexes are less physiological phospholipids.
It is important that DMPC and adenovirus be solubilized in PLs without precipitation. However, in the current study, the solubility of Ad was enhanced in the PL solution containing WT or V156K when compared with DMPC liposomes alone. Without sonication, the DMPC in PBS existed primarily in the MLV state. This indicates that apoA-I in the PLs contributes to the association of Ad and phospholipid by inducing increased solubility of the Ad particles in PLs, which provides a micellar environment for the viral particles.
There have been many attempts to introduce an exogenous gene into various target cells, using adenovirus. Kawasaki and colleagues (2009) reported the successful infection of zebrafish embryos with adenovirus. Zebrafish are a good model to test gene delivery efficiency because a zebrafish homolog of the coxsackievirus and adenovirus receptor protein has been identified (Petrella et al., 2002). Furthermore, zebrafish have a well-developed innate and acquired immune system that is similar to the mammalian immune system (Trede et al., 2001). To our knowledge, this is the first report to show that Ad can be infected into adult zebrafish, especially in PLs containing apoA-I (Fig. 8). V156K-PL-Ad showed nearly 2-fold enhanced expression of GFP compared with WT-PL-Ad. This result might correlate well with the following improved properties of V156K: enhanced DMPC binding ability (Fig. 2), greater protein stability in the presence of a denaturant (Fig 3), and enhanced viral stability after incubation at 25 and 37°C in the PL state (Fig. 7).
In conclusion, adenoviral gene delivery and the stability of Ad in vitro and in vivo were enhanced by formulation with PLs containing WT or V156K apoA-I, virus, DMPC, and cholesterol, which indicate that this mode of treatment would allow use of a lower dose and a longer storage time. Furthermore, the enhanced adenoviral gene delivery efficiency, as well as the antiinflammatory and antioxidant activities of apoA-I administered in this fashion, may be used to treat cancer in a synergistic manner.
Footnotes
Acknowledgments
This work was supported by grants from the Korea Science and Engineering Foundation (R15-2004-024-02001-0, C-O. Yun), and supported by NRF (National Research Foundation of Korea) through the Aging-associated Vascular Disease Research Center at Yeungnam University (R13-2005-005-01003-0 [2009], K-H. Cho).
Author Disclosure Statement
No competing financial interests exist.