
Abstract
Previous studies showed that stimulation of T cells derived from HIV-1-infected patients with autologous dendritic cells electroporated with mRNA encoding HIV antigens can induce antigen-specific T cell responses in vitro. Linking the antigen to an MHC class II-targeting sequence, such as dendritic cell lysosome-associated membrane protein (DC-LAMP), in the mRNA construct results in presentation of antigenic peptides in both MHC class I and class II molecules and therefore enhances the induced T cell responses. To analyze whether the lumenal domain of DC-LAMP is required for optimal induction of cellular immunity against HIV antigens, we compared fusion constructs with or without the lumenal domain of the DC-LAMP protein. A human codon-optimized consensus Gag sequence and a chimeric cDNA sequence encompassing Tat, Rev, and Nef codons (TaReNef ) were cloned into a vector containing the DC-LAMP sequence with or without its lumenal domain. The Gag protein lacking the DC-LAMP-derived sequence altogether elicited only weak T cell responses. DCs electroporated with Gag or TaReNef linked to DC-LAMP were able to elicit similar levels of antigen-specific CD4+ and CD8+ T cell responses for both Gag and TaReNef, irrespective of the addition of the DC-LAMP lumenal domain. These data show that DC-LAMP-mediated antigen targeting is absolutely required for optimal T cell stimulation, but that in our experimental setup, the lumenal part of DC-LAMP does not improve the overall induction of antigen-specific T cell responses.
Introduction
In this study, we compared fusion constructs with (pSig-Antigen-DC-LAMPLumTMCy) or without (pSig-Antigen-DC-LAMPTMCy) the lumenal domain of DC-LAMP. We stimulated T cells derived from HIV-infected patients with autologous dendritic cells electroporated with various Gag or TaReNef (a chimeric construct encompassing Tat, Rev, and Nef; Allard et al., 2008) mRNA constructs. To our knowledge, this is the first time that mRNA constructs encoding antigen linked to DC-LAMP, with or without its lumenal domain, were compared in vitro, using T cells and dendritic cells derived from HIV-infected patients.
Materials and Methods
Study subjects
Patients were recruited from the AIDS Unit of the Department of Internal Medicine (Universitair Ziekenhuis Brussel, Brussels, Belgium) and the Department of Internal Medicine II (Erasmus Medical Center, Rotterdam, The Netherlands). Patient selection criteria were as follows: HIV-1 seropositivity as determined by Western blot, and highly active antiretroviral therapy (HAART) regimen with CD4+ T cell count: 500/mm3 and viral load <50 copies/ml for at least 3 and 6 months, respectively. Samples were obtained from buffy coats or by leukapheresis.
mRNA constructs
The plasmid pST1, which was used for all in vitro mRNA syntheses, was obtained from U. Sahin (Saarland University Medical School, Homburg, Germany).
For cloning of the pST1-Gag construct, the full open reading frame of Gag was isolated from pCR-Gag as a HinDIII–XhoI fragment and cloned in pST1. The gag gene (B clade) was codon optimized by Geneart (Regensburg, Germany).
The plasmid pST1-Sig-DC-LAMPTMCy was generated by cloning a HinDIII–XhoI Sig-DC-LAMP fragment from pGEM-Sig-DC-LAMP (Bonehill et al., 2004) into pST1. This construct contains a unique BamHI site at the boundary of the signal sequence and the transmembrane–cytoplasmic (TMCy) region of DC-LAMP. The vector pST1-Sig-DC-LAMPLumTMCy was created by replacing the signal sequence in pST1-Sig-DC-LAMP by the lumenal region (Lum) of DC-LAMP. This gene fragment was obtained by PCR of cDNA from mature human dendritic cells. During this amplification a 5′ HinDIII site and a Kozak consensus sequence were introduced. At the 3′ end a BamHI site, in frame with the BamHI site at the 5′ end of the DC-LAMP transmembrane–cytoplasmic region, was generated.
A Gag fragment without start or stop codon was generated by PCR. At both ends a BamHI site was introduced, which allows cloning in frame with the existing site in both pST1-Sig-DC-LAMPTMCy and pST1-Sig-DC-LAMPLumTMCy.
pST1-Sig-TaReNef-DC-LAMPTMCy was generated by cloning the Sig-TaReNef-DC-LAMP fragment from pGEM-Sig-TaReNef-DC-LAMP (Allard et al., 2008) as a HinDIII–XhoI fragment.
pST1-Sig-TaReNef-DC-LAMPLumTMCy was obtained after cloning the HinDIII–BamHI lumenal DC-LAMP fragment in pST1-Sig-TaReNef-DC-LAMPTMCy, digested with the same enzymes, replacing Sig. A schematic representation of all mRNA constructs is shown in Fig. 1.

Schematic representation of the mRNA constructs used in this study. 1a, LAMP signal peptide; 1b, DC-LAMP signal peptide; 2, DC-LAMP lumenal domain; 3, antigen (Gag or TaReNef ); 4, DC-LAMP transmembrane and cytoplasmic domain; 5, 3′ human β-globin; 6, poly(A) tail (A120).
Peptides
HIV-1 clade B consensus Gag, Tat, Rev, and Nef 15-mer peptides overlapping by 11 amino acids were obtained from National Institutes of Health AIDS Research and Reference Reagent Program (Rockville, MD). They were dissolved in 10% dimethyl sulfoxide (DMSO) in 10 mM acetic acid, pooled at 10 peptides per pool, and stored in aliquots at a concentration of 2 mg/ml at −20°C.
Cell culture and reagents
All cultures were performed in a humidified atmosphere containing 5% CO2 at 37°C. Dendritic cells (DCs) were cultured as described earlier (Allard et al., 2008).
T cells were cultured in Iscove's modified Dulbecco's medium (IMDM; Cambrex, Verviers, Belgium), supplemented with 10% heat-inactivated human AB serum (PAA laboratories, Linz, Austria) containing penicillin (100 U/ml), streptomycin (100 μg/ml), and 2 mM
Human recombinant granulocyte-macrophage colony-stimulating factor (GM-CSF), interleukin (IL)-1β, IL-4, and IL-6 were produced in-house; tumor necrosis factor (TNF)-α was purchased from PeproTech (London, UK). Prostaglandin E2 (PGE2) was obtained from Pharmacia (Puurs, Belgium).
Generation and maturation of monocyte-derived dendritic cells
DCs were derived from the adherent cell fraction of buffy coats or by leukapheresis. When using buffy coat material, DCs were generated as described earlier (Allard et al., 2008). When using leukapheresis, DCs were generated in four-tray Cell Factories (Nunc/Thermo Fisher Scientific, Rochester, NY) as previously described (Tuyaerts et al., 2002). DCs were matured by adding a mixture of inflammatory cytokines including IL-1, IL-6, TNF-α, and PGE2 to the DC cultures, as described earlier (Allard et al., 2008).
Electroporation of DCs and EBV-B cells
mRNA electroporation of mDCs and EBV-B cells was performed with an EasyjecT Plus apparatus (EquiBio, Kent, UK), as previously described (Michiels et al., 2005). For each electroporation, 4 million cells were electroporated with 20 μg of mRNA.
T cell culture
On day 0, T cells were thawed and rested overnight in lymphocyte medium supplemented with IL-2 (25 U/ml; Chiron, Emeryville, CA) and DNase (10 U/ml). On day 1, mRNA-electroporated DCs were cocultured with the T cells (DC:T cell ratio, 1:10) in lymphocyte medium supplemented with IL-7 (10 ng/ml; Peprotech) and 400 μM 1-methyl-
Intracellular FACS staining
To evaluate the expression of Gag after its electroporation into DCs, brefeldin A (GolgiPlug; BD Biosciences, San Jose, CA) was added to the DC cultures immediately after electroporation. Using a BD Cytofix/Cytoperm kit (BD Biosciences), intracellular Gag was stained 4 and 24 hr after electroporation with KC57-RD1 (phycoerythrin)-conjugated antibody (Beckman Coulter, Fullerton, CA), in accordance with the manufacturer's instructions. Intracellular cytokine stainings were performed on T cells stimulated overnight with EBV-B cells electroporated with antigen-encoding mRNA in the presence of brefeldin A. Cytokine staining was performed with the BD Cytofix/Cytoperm kit combined with the following antibodies: phycoerythrin–Cy7 (PE–Cy7)-conjugated CD4 (BD Biosciences), allophycocyanin (APC)-conjugated CD8 (BD Biosciences), and PE-conjugated interferon (IFN)-γ and fluorescein isothiocyanate (FITC)-conjugated IL-2 (both from eBioscience, San Diego, CA). Data were analyzed with a FACSCanto flow cytometer using FACSDiva software (BD Biosciences).
IFN-γ ELISPOT
An IFN-γ ELISPOT assay was performed as described earlier (Tuyaerts et al., 2003). In each well, 2 × 105 T cells were cocultured with 2 × 104 electroporated DCs (overnight ELISPOT) or 1 × 105 T cells were cocultured with 1 × 104 EBV-B cells (ELISPOT after restimulation). For screening with peptide libraries, T cells were stimulated with EBV-B cells pulsed with pools of ±10 overlapping 15-mer peptides of Gag, Tat, Rev, or Nef. Spots were counted with an AID EliSpot reader (Autoimmun Diagnostika, Strassberg, Germany). As negative controls, T cells were stimulated with mock-electroporated DCs or EBV-B cells. As a positive control, T cells were stimulated with phorbol myristate acetate (PMA, 10 ng/ml) and ionomycin (10 μg/ml).
Results and Discussion
To evaluate antigen expression on electroporation, DCs were electroporated with 20 μg of Gag, Gag-DC-LAMPTMCy, or Gag-DC-LAMPLumTMCy. At 4 and 24 hr after electroporation, all cells electroporated with one of the Gag mRNA constructs displayed strong intracellular Gag expression. DCs electroporated with Gag mRNA showed slightly lower Gag expression compared with DCs electroporated with DC-LAMP-linked Gag constructs. Histograms representing Gag expression 24 hr after electroporation into DCs are shown in Fig. 2.
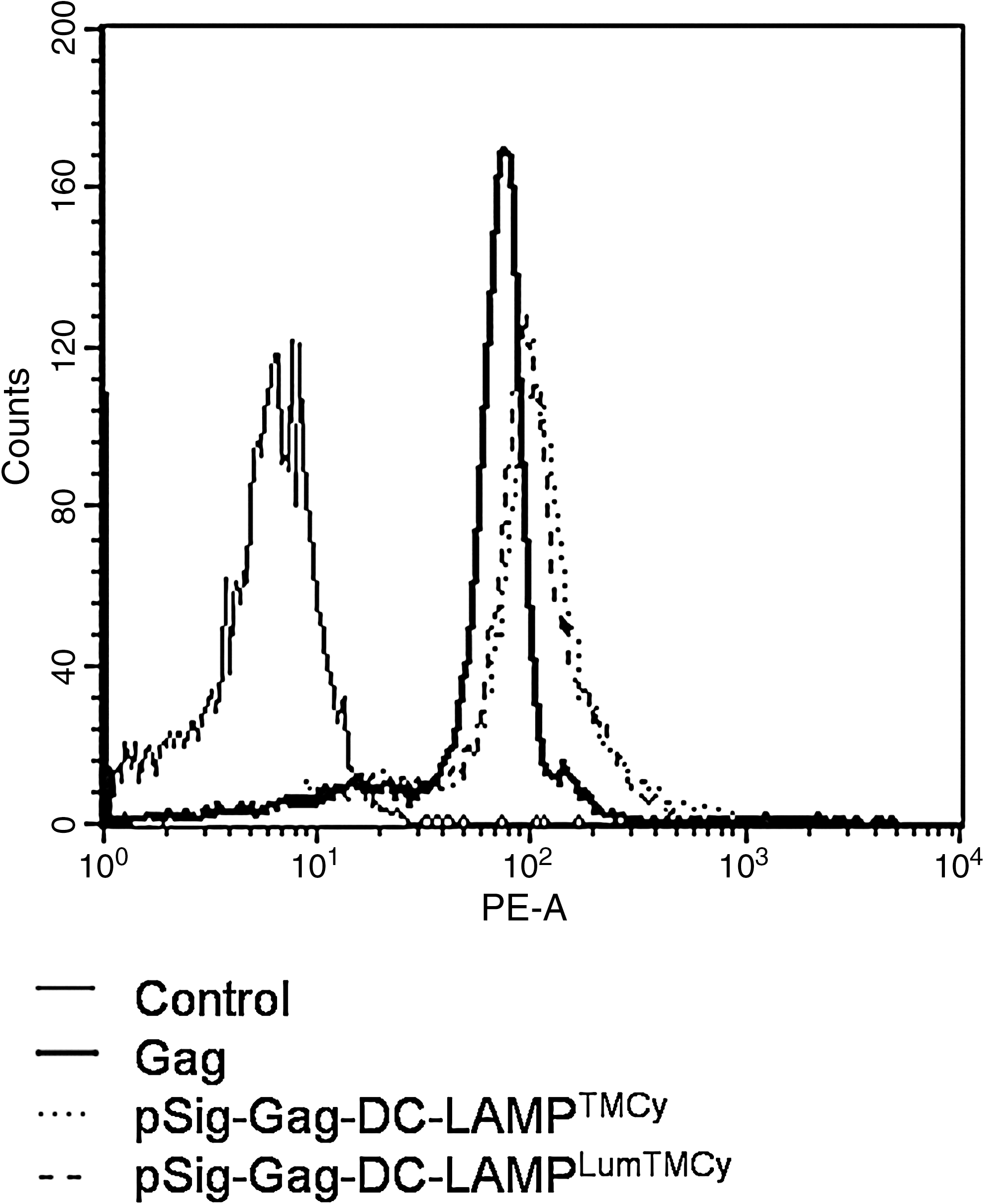
Antigen expression upon electroporation. FACS staining was performed to analyze intracellular Gag expression after electroporation. Shown is Gag expression in DCs 24 hr after electroporation with each of the various Gag mRNA constructs. As a control, mock-electroporated DCs were stained with the Gag antibody. Thin line, control; thick line, Gag; dotted line, pSig-Gag-DC-LAMPTMCy; dashed line, pSig-Gag-DC-LAMPLumTMCy.
To compare the T cell-stimulatory capacity of DCs electroporated with various Gag constructs, an overnight IFN-γ ELISPOT was performed. Both Gag-DC LAMPTMCy and Gag-DC-LAMPLumTMCy constructs caused an increase in the number of IFN-γ spot-forming units (SFU) compared with mock-stimulated T cells. For all four patients analyzed, the number of IFN-γ spot-forming units induced by DCs electroporated with Gag-DC-LAMPTMCy was at least as high as the number of spot-forming units caused by Gag-DC LAMPLumTMCy-electroporated DCs. DCs electroporated with unmodified Gag induced only a marginal increase in the number of IFN-γ spot-forming units compared with T cells cocultured with mock DCs (Fig. 3a). Similar results were obtained for TaReNef; T cells stimulated with pSig-TaReNef-DC-LAMPTMCy- or pSig-TaReNef-DC-LAMPLumTMCy-electroporated DCs induced similar numbers of IFN-γ spot-forming units (Fig. 3c).

Induction of HIV-1 Gag- and TaReNef-specific T cell responses. T cells from chronically HIV-1-infected subjects were stimulated with DCs electroporated with the various Gag or TaReNef mRNA constructs. T cell responses were analyzed after overnight stimulation (left). After 11 or 12 days of stimulation, the T cells were restimulated overnight with Gag or TaReNef mRNA-electroporated EBV-B cells (right). Each graph shows the number of IFN-γ spot-forming units (SFU) per million T cells. Each dot represents the mean number of spot-forming units for one patient (mean of three replicate wells). Each vertical line shows the mean spot-forming units of all patients (n = 4 for Gag stimulations, n = 3 for TaReNef stimulations). Data derived from overnight ELISPOTs are shown in (
To examine the effect of the various DC-LAMP constructs on HIV-specific T cell responses on DC vaccination, immune responses elicited after an 11- or 12-day stimulation with DCs electroporated with the various Gag constructs were determined by IFN-γ ELISPOT as an in vitro approximation. After restimulation with EBV B cells electroporated with the same mRNA construct as the one used for the 11 or 12 days of stimulation (Gag, Gag-DC-LAMPTMCy, or Gag-DC-LAMPLumTMCy), the number of IFN-γ spot-forming units elicited by the Gag-DC-LAMP constructs was always higher compared with the response elicited by the Gag construct, which caused only weak responses. No difference was observed between T cell responses induced by 11 or 12 days of in vitro stimulation with Gag-DC-LAMPTMCy or Gag-DC-LAMPLumTMCy mRNA-electroporated DCs (Fig. 3b). Similar results were also obtained after T cell restimulation with EBV-B cells electroporated with TaReNef- DC-LAMPTMCy or pSig-TaReNef-DC-LAMPLumTMCy (Fig. 3d).
To evaluate whether the different DC-LAMP constructs induced responses against different antigenic regions of Gag, T cells were restimulated with autologous EBV-B cells pulsed with pools of overlapping 15-mer peptides. The number of IFN-γ spot-forming units was comparable for cells primed with Gag-DC-LAMPTMCy- or Gag-DC-LAMPLumTMCy-electro-porated DCs and the same pools were recognized. T cells primed with DCs electroporated with unmodified Gag mRNA showed a weaker response to the Gag peptides (Fig. 4). For some patients, we also analyzed T cell responses after restimulation with peptide libraries derived from Tat, Rev, and Nef in ELISPOT. Again, no clear differences in T cell responses between pSig-TaReNef-DC LAMPTMCy- and pSig-TaReNef-DC-LAMPLumTMCy-primed T cells were found (data not shown).
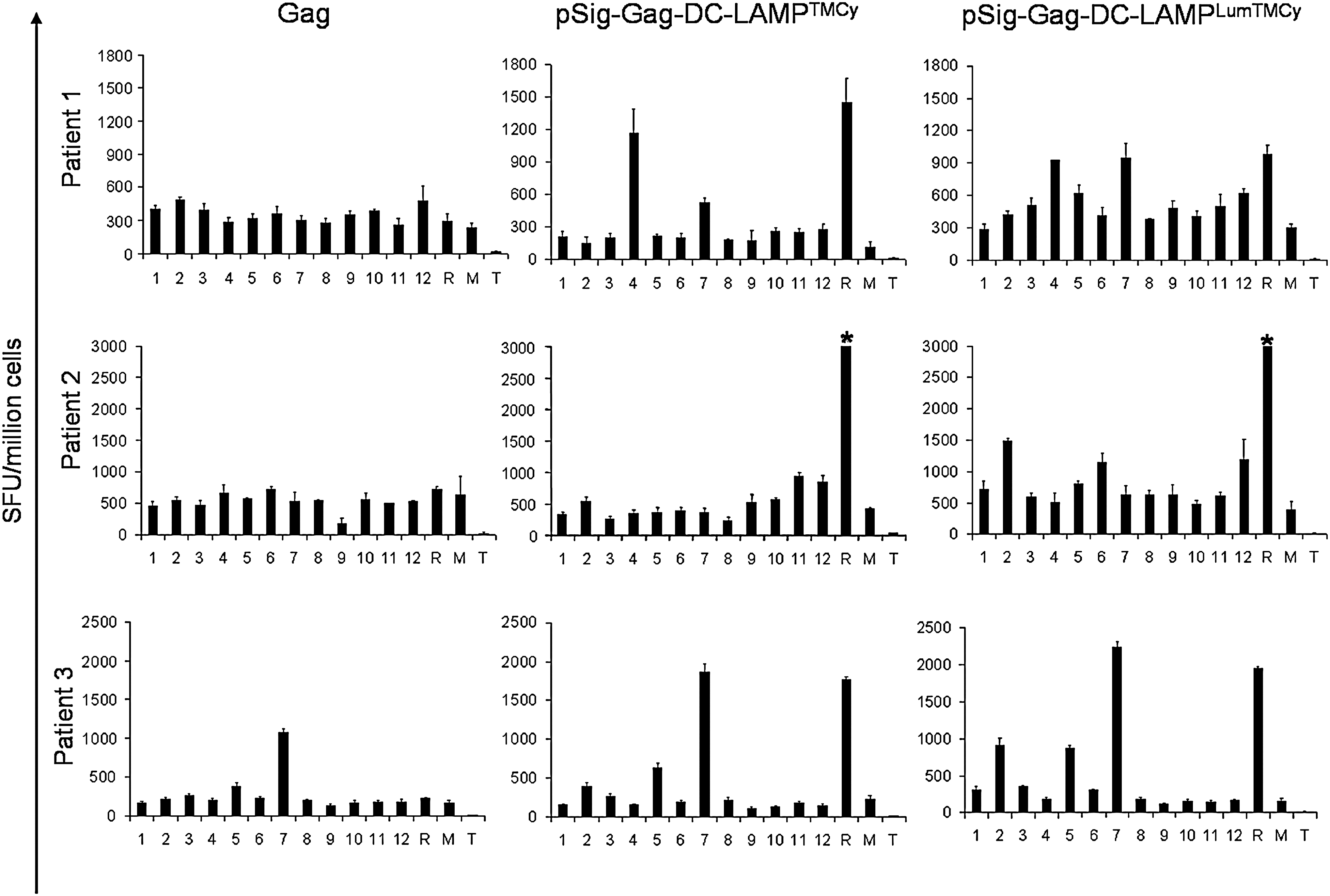
Epitope screening, using libraries of overlapping peptides in an IFN-γ ELISPOT assay. T cells from HIV-infected patients were stimulated in vitro for 11 or 12 days with DCs electroporated with Gag, pSig-Gag-DC-LAMPTMCy, or pSig-Gag-DC-LAMPLumTMCy. After the primary stimulation, T cells were restimulated overnight with EBV-B cells pulsed with pools of Gag peptides, within an ELISPOT plate. Peptide pools are indicated by the number of the pool. As controls, cells were stimulated with mRNA-electroporated EBV-B cells (R), mock-electroporated EBV-B cells (M), or medium (T). Error bars indicate the SEM. An asterisk (*) indicates that the number of spot-forming units exceeded the resolution of the ELISPOT reader.
Because it has been shown that both CD4+ and CD8+ T cell responses are important for protective immune responses against HIV and that addition of the DC-LAMP targeting signal is important for presentation with both MHC class I and class II molecules, we analyzed whether Gag-electroporated DCs were able to enhance both CD4+ and CD8+ T cell responses. We stimulated T cells for 11 or 12 days with DCs electroporated with the various Gag mRNA constructs and restimulated them overnight with electroporated EBV-B cells. After overnight restimulation, the cells were stained intracellularly for IFN-γ and IL-2 and analyzed by flow cytometry. For T cells primed with Gag-DC-LAMPTMCy or Gag-DC-LAMPLumTMCy, Gag-specific responses were found in both the CD4+ (e.g., patient 5) and CD8+ (e.g., patient 4) T cell fractions. Cytokine production by T cells primed with Gag only was much lower. We were able to detect T cells that produced both IFN-γ and IL-2 in only one patient (Fig. 5a). A summary of the percentages of IFN-γ+ CD4+ and CD8+ T cells as measured by the intracytoplasmic cytokine stainings is shown in Fig. 5b.
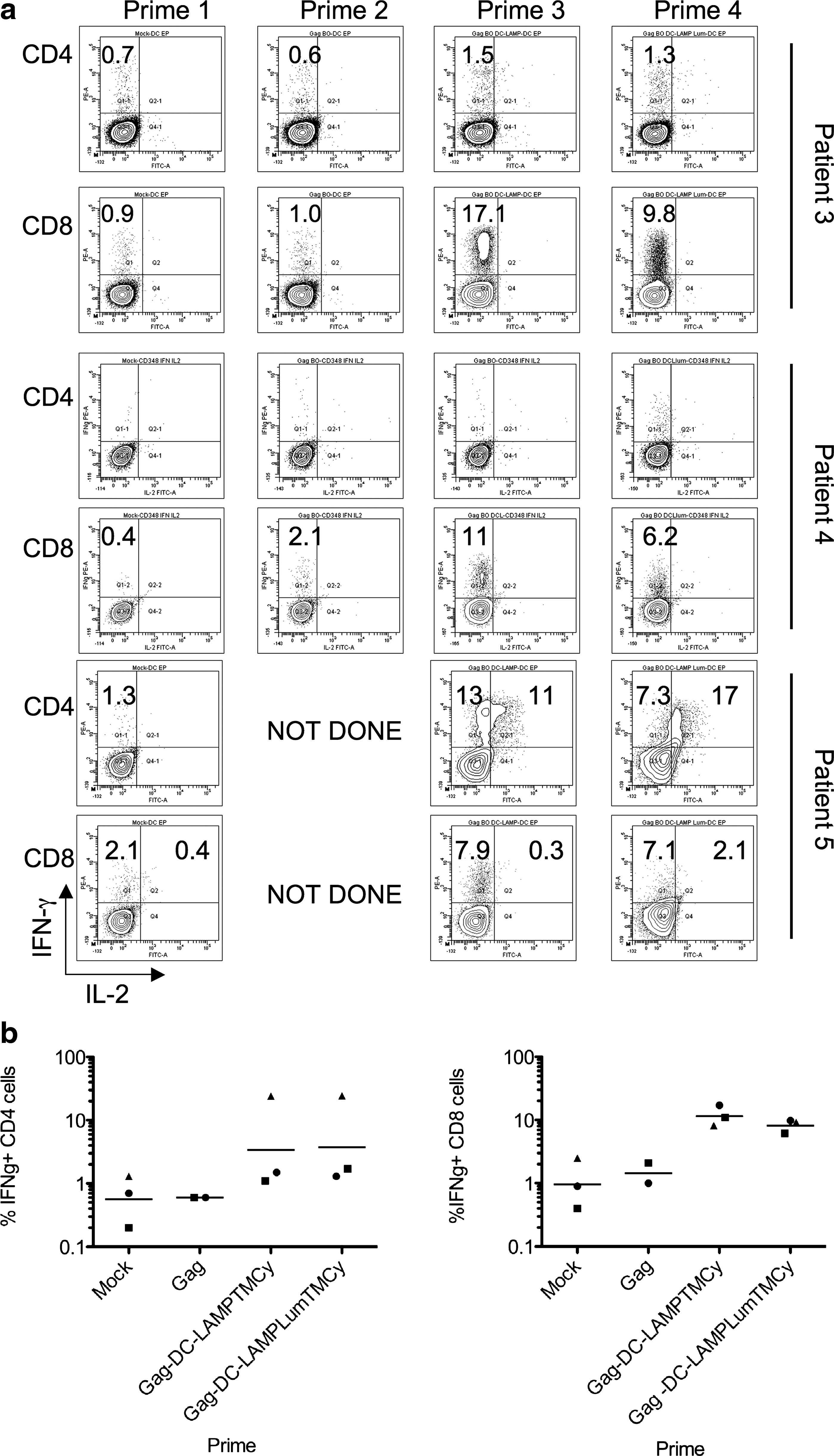
CD4+ and CD8+ T cell Gag responses determined by intracellular FACS staining. T cells were stimulated for 11 or 12 days with mock-electroporated DCs (Prime 1) or with DCs electroporated with unmodified Gag (Prime 2), pSig-Gag-DC-LAMPTMCy (Prime 3), or pSig-Gag-DC-LAMPLumTMCy (Prime 4). Afterwards, the cells were restimulated overnight with electroporated EBV-B cells and stained intracellularly with IFN-γ and IL-2. The plots in (
In summary, addition of the lumenal part of DC-LAMP to the mRNA constructs did not improve the overall induction of HIV-specific CD4+ and CD8+ T cell responses against Gag and TaReNef, nor could it broaden the T cell responses against these antigens. It has been shown that Gag encoded with LAMP, without its lumenal domain, did not traffic to lysosomes or MHC II compartments of DNA-transfected cells and elicited a limited immune response in DNA-immunized mice, whereas addition of the LAMP lumenal domain to the construct resulted in a high level of expression of the Gag–LAMP chimeric protein in transfected cells and in strong cellular and humoral immune responses of immunized mice (Marques et al., 2003). However, differences between our observations and the results previously described by Marques and colleagues may be explained by the differences in experimental setup. First, we used T cells derived from HIV-infected patients in order to evaluate T cell responses elicited by electroporated DCs in vitro. Marques and colleagues tested their various DNA constructs by immunizing mice, which are not natural hosts for HIV. In this mouse system, there is no immune dysfunction effect typical of the T cells derived from an HIV-infected patient. Another major difference is the fact that in the Marques study a prophylactic vaccination setup was used, whereas we wanted to examine the effect of various targeting constructs in a therapeutic setting. Second, we used mRNA electroporation of DCs for our stimulations, whereas Marques and colleagues used a DNA vaccination approach; mice were injected intramuscularly with naked plasmid DNA. This most likely will result in the expression of Gag in (antigen-presenting) cells other than DCs, resulting in different T cell responses. Furthermore, we used full-length Gag whereas Marques and colleagues used a construct consisting of p55Gag only. Finally, we tested chimeric constructs that consist of Gag or TaReNef linked to DC-LAMP, whereas Marques and colleagues described the effects induced by a construct in which Gag is linked to LAMP. In another study (Arruda et al., 2006), T cell responses induced in Gag-DC-LAMP- and Gag-LAMP-immunized mice were compared and it was shown that Gag-DC-LAMP and Gag-LAMP follow different cellular trafficking pathways, that IL-4 production between the two mouse groups was different, and that mice immunized with Gag-DC-LAMP and Gag-LAMP showed T cell responses to other peptides. However, no significant differences between the groups were found in the amount of IFN-γ secreted by CD4+ or CD8+ T cells.
We showed that DC-LAMP-mediated antigen targeting is absolutely required for optimal T cell stimulation of both CD4+ and CD8+ T cell subsets, as DCs electroporated with unmodified Gag mRNA were not able to elicit T cell responses that exceeded background values, in contrast to Gag-DC-LAMP-electroporated DCs. This is in line with earlier results showing that DCs electroporated with the tumor antigens Melan-A and MAGE-A3 linked to DC-LAMP were not only indispensable for CD4+ T cell activation but also showed a greatly enhanced stimulatory capacity for CD8+ T cell clones compared with constructs without any targeting signal (Bonehill et al., 2004). This might be due to the translation of abnormal proteins and the increased formation of so-called DriPs, or defective ribosomal products. The enhanced peptide presentation after electroporation with the DC-LAMP-containing mRNA might be due to presentation by recycling HLA class I molecules. Internalized HLA class I molecules can associate with peptides generated in the endosomal pathway. In other studies, Gag mRNA constructs without an HLA class I targeting sequence were able to induce robust T cell responses (e.g., Van Gulck et al., 2008). However, most studies have used truncated versions of Gag for T cell stimulation, whereas we used full-length Gag, which might be more prone to degradation. This hypothesis is supported by the fact that enhanced CD8+ T cell stimulation was found when the truncated Gag was used in a side-by-side comparison with the full-length Gag versus the partially truncated Gag used in the study published by Van Gulck and colleagues (2008) (our unpublished results). As the last 63 amino acids (amino acids 467–529) from the Gag protein are missing from the partially truncated Gag, thus potentially omitting epitopes from this region, we decided to continue our experiments with the full-length Gag construct.
To conclude, we show that DC-LAMP-mediated antigen targeting is absolutely required for optimal induction of Gag- and TaReNef-specific T cells. For the stimulation of HIV-infected patient-derived T cells with autologous DCs electroporated with mRNA, the lumenal part of DC-LAMP does not improve the overall antigen-specific T cell responses.
Footnotes
Acknowledgments
The authors thank Elsy Vaeremans and Sandra Van Lint for the preparation of mRNA; Jurgen Corthals, Inge Betz, Gwenny de Metter, Hilde Lambrecht, and Mattias Van den Abeele for help with the isolation of T cells and DCs; and the staff of the Department of Internal Medicine (UZ Brussel, Brussels, Belgium) and the Department of Internal Medicine II (Erasmus Medical Center, Rotterdam, The Netherlands) for the recruitment of patients. This work was supported by the DC-THERA Network, the Cancer Immunology and Immunotherapy Research Project (EU254), and the Institute for Science and Technology (IWT, IWT-TBM 60511 project). J.A. and A.B. are postdoctoral fellows of the FWO (Fund for Scientific Research Flanders).
Author Disclosure Statement
No competing financial interests exist.