
Abstract
Transcriptional activation of gene expression directed by the long terminal repeat (LTR) of HIV-1 requires both the transactivation response element (TAR) and Tat protein. HIV-1 mutants lacking a functional tat gene are not able to proliferate. Here we take a genetic approach to suppress HIV-1 replication based on Tat-dependent production of MazF, an ACA-specific endoribonuclease (mRNA interferase) from Escherichia coli. When induced, MazF is known to cause Bak- and NBK-dependent apoptotic cell death in mammalian cells. We first constructed a retroviral vector, in which the mazF (ACA-less) gene was inserted under the control of the HIV-1 LTR, which was then transduced into CD4+ T-lymphoid CEM-SS cells in such a way that, upon HIV-1 infection, the mazF gene is induced to destroy the infecting HIV-1 mRNA, preventing HIV-1 replication. Indeed, when the transduced cells were infected with HIV-1 IIIB, the viral replication was effectively inhibited, as HIV-1 IIIB p24 could not be detected in the culture medium. Consistently, not only cell growth but also the CD4 level was not affected by the infection. These results suggest that the HIV-1-LTR-regulated mazF gene was effectively induced upon HIV-1 IIIB infection, which is sufficient enough to destroy the viral mRNA from the infected HIV-1 IIIB to completely block viral proliferation in the cells, but not to affect normal cell growth. These results indicate that the T cells transduced with the HIV-1-LTR-regulated mazF gene acquire HIV-1 resistance, providing an intriguing potential for the use of the HIV-1-LTR-regulated mazF gene in anti-HIV gene therapy.
Introduction
In the HIV-1 life cycle immediately after HIV-1 infection, Tat (transactivator of transcription), an early regulatory protein encoded by the HIV-1 genome, is produced, which subsequently binds to the TAR (transactivation response) sequence to induce the transcription of the HIV-1 genome leading to the expression of other HIV-1 proteins (Berkhout et al., 1989). Therefore, for prevention of HIV-1 infection, it would be a best strategy to preferentially destroy the HIV-1 transcript upon HIV-1 infection. For this purpose, we constructed a Tat-dependent MazF expression system in a retroviral vector, in which the mazF gene was fused downstream of the TAR sequence. As the E. coli mazF open-reading frame contains nine ACA sequences, all of them were engineered to MazF-uncleavable sequences without changing the amino acid sequence of MazF. This vector was then transduced into T cells so that MazF production is expected to be induced upon HIV-1 infection. Note that Tat protein produced upon HIV-1 infection induces not only the transcription of infected HIV-1, but also the transcription of the HIV-1 long terminal repeat (LTR)-regulated mazF (ACA-less) gene integrated into the genome of the T cells. In the present article, CD4+ T lymphoid line CEM-SS cells were used as T cells, which were transduced with the retroviral vector containing the Tat-inducible mazF (ACA-less) gene under the HIV-1-LTR promoter. When the transduced cells were infected with HIV-1 IIIB, the replication of the infected virus was effectively inhibited without affecting cell growth. Notably, the CD4 level after HIV-1 IIIB infection was not affected either. These results suggest that the HIV-1-LTR-regulated mazF (ACA-less) gene was effectively induced upon HIV-1 IIIB infection, which is sufficient enough to destroy the viral mRNA from the infected HIV-1 IIIB to completely block viral proliferation in the cells. However, the level of MazF induced is not enough to cause any serious cellular damage, thus maintaining normal cell growth and the CD4 level. These results suggest an intriguing potential for the use of the HIV-1-LTR-regulated mazF (ACA-less) gene in anti-HIV gene therapy.
Materials and Methods
Cell lines
293T (ATCC no. CRL-11268) cells were cultured in Dulbecco's modified Eagle medium (DMEM; Sigma-Aldrich, Steinheim, Germany) supplemented with 10% (v/v) fetal bovine serum (FBS; Invitrogen, Carlsbad, CA). CEM-SS cells (Kim et al., 1989) were cultured in RPMI-1640 (Sigma-Aldrich) containing 10% (v/v) FBS (Invitrogen). The doubling time of the cells for each culture condition was calculated by linear regression analysis using Microsoft Excel software (Microsoft, Seattle, WA).
Retroviral vectors
The self-inactivating retroviral vector pMTD3 was constructed by deleting a segment consisting of 267 nucleotides from the 3’LTR U3 region of pMT (Lee et al., 2004). An ACA-less mazF gene was synthesized by engineering all nine ACA sequences in the original E. coli mazF gene to MazF-uncleavable sequences without changing the amino acid sequence of MazF. The HIV-LTR fragment was obtained from pQBI-LTRgagGFP (Quantum Biotechnologies Inc., Montreal, QC, Canada). To minimize the HIV-LTR sequence, U3-TAR fragments were obtained by PCR. The ACA-less mazF gene was inserted downstream of U3-TAR to obtain the final self-inactivating retroviral vector plasmid, pMTD3-U3TAR-MazF. As a control, the green fluorescent protein (GFP) gene was inserted into the vector to obtain pMTD3-U3TAR-GFP.
To mimic HIV replication, two kinds of retroviral vectors that express the HIV-1 Tat protein were constructed as follows: (1) Constitutitve Tat expression system from MLV-LTR. The HIV-1 tat gene was synthesized and inserted at the multiple-cloning site of pMT. To easily monitor the gene expression in transduced cells, an internal ribosome entry site (IRES) and a coding region for a fluorescent protein, ZsGreen, were fused downstream of the tat gene. Thus, the resulting plasmid, pM-LTR-Tat-ZG, expresses Tat as well as ZsGreen from MLV-LTR. (2) Tat expression system from the HIV-1 LTR. The HIV-LTR-tat-polyA cassette was inserted in the opposite direction of pMT, and the ZsGreen marker gene was expressed from a phosphoglycerate kinase (PGK) promoter in the normal orientation of pMT. The resulting vector plasmid was designated as pH-LTR-Tat-ZG.
To enhance the viral titer for efficient mazF gene transduction, the HIV-LTR-MazF-polyA cassette was introduced in the opposite direction of the MoMLV-LTR at the multi-cloning site of pMT plasmid (Lee et al., 2004). A truncated form of the human low-affinity nerve growth factor gene (ΔLNGFR) (Verzeletti et al., 1998) was also introduced into the retrovirus vector as a surface marker. The ΔLNGFR gene is under the control of human PGK promoter. The resultant vector plasmid was designated as pMT-MFR-PL2 (Fig. 1B).

Construction of retroviral vector under the control of HIV-LTR promoter. To remove promoter activity of the MoMLV LTR, the self-inactivating retroviral vector pMTD3 was constructed based on pMT (Lee et al., 2004) by deleting a 276-bp fragment from its 3’LTR U3 region. A synthetic ACA-less mazF gene was then inserted downstream of HIV-1 U3-TAR resulting in the self-inactivating retroviral vector, pMTD3-U3TAR-MazF. As a control, the GFP gene was inserted in place of the mazF gene, which resulted in pMTD3-U3TAR-GFP. The self-inactivating retroviral vectors were generated using the transient transfection method with the packaging plasmids MoMLV-gag-pol, GALV-env, and the self-inactivating retroviral vector in 293T cells. The viral preparation was obtained 2 days after transfection by filtering the culture supernatant. To improve the viral titer for efficient gene transduction over an initial vector, HIV-LTR-MazF-polyA cassette was inserted in the opposite direction of the MoMLV-LTR at the multi-cloning site of pMT. A truncated form of the human low-affinity nerve growth factor gene (ΔLNGFR) (Verzeletti et al., 1998) was used as a surface marker. The resultant vector plasmid was designated pMT-MFR-PL2. GALV-env retroviral vector was generated as described in Materials and Methods.
Preparation of retroviral vectors
The self-inactivating retroviral vector was generated by the transient transfection method as follows: The GALV-env expression vector plasmid, pVM-GeR, was constructed by replacing the amphotropic-env gene of pVM-AE (Yu et al., 2003) with the gibbon ape leukemia virus envelope gene. The GALV-env retroviral vector was produced by co-transfecting 293T cells with the retroviral gag-pol expression vector plasmid, pVM-GP (Yu et al., 2003), pVM-GeR, and the self-inactivating retroviral vector plasmid. Two days after transfection, viral supernatant was harvested by filtration of the culture fluid from 293T cells with use of a 0.45-μm filter.
GALV-enveloped retroviral Tat expression vectors and MazF expression vector were also generated as follows: Ecotropic retroviral vectors were generated by the transient transfection method using the packaging plasmids pGP (MLV-gag-pol; Takara Bio, Otsu, Shiga, Japan) and pE-eco (ecotropic env; Takara Bio) with the retroviral vector plasmid pM-LTR-Tat-ZG, pH-LTR-Tat-ZG, or pMT-MFR-PL2. This was performed with use of human embryonic kidney 293T-derived G3T-hi cells (Takara Bio) by using the calcium phosphate co-transfection method. The GALV-env retroviral vector was obtained from PG13 packaging cells (ATCC no. CRL-10686) by infection with the ecotropic retrovirus vector as prepared above. After selection of the infected PG13 cells, the virus was collected from the growth medium by filtration of the supernatant with use of a low-protein binding filter (0.45 μm).
Retroviral transduction into CEM-SS cells
CEM-SS cells were infected with self-inactivating retroviral vectors in the presence of 8 μg/ml Polybrene (hexadimethrine bromide; Sigma-Aldrich). Polyclonal gene-transduced cell populations were obtained by selecting the cells with G418 (Invitrogen) at a concentration of 1 mg/ml.
CEM-SS cells or CEM-SS cells transduced with MTD3-U3TAR-MazF were infected with Tat expression retroviral vectors M-LTR-Tat-ZG or H-LTR-Tat-ZG in the presence of RetroNectin (Takara Bio) according to the manufacturer's protocol.
Retroviral transduction into primary rhesus macaque CD4+ T cells
Rhesus macaque CD4+ T cells were isolated from peripheral blood mononuclear cells (PBMC) using anti-CD4 monoclonal antibody-conjugated beads (Dynal CD4 Positive Isolation Kit; Invitrogen). Prior to gene transduction, the isolated CD4+ T cells were activated for 3 days with a combination of anti-monkey-CD3 clone FN-18 (BioSource, Camarillo, CA) and anti-human-CD28 monoclonal antibody clone L293 (BD Biosciences, Franklin Lakes, NJ)-conjugated beads at a cell-to-bead ratio of 1:1 in GT-T503 (Takara Bio) supplemented with 10% FBS and 200 IU of interleukin-2 (Chiron, Emeryville, CA). On day 3, activated CD4+ T cells were infected with the MazF retroviral vector (MT-MFR-PL2) in the presence of RetroNectin (Takara Bio) as per the manufacturer's instructions. The transduction was repeated again on day 4. The cells were further incubated for another 3 days. The genetically modified cells marked with the ΔLNGFR+ were concentrated with anti-CD271 monoclonal antibody-conjugated beads (CD271 MicroBeads; Miltenyi Biotec, Bergisch Gladbach, Germany). Aliquots of the mazF gene-modified cells (designated as MazF-Tmac cells) were collected and cryopreserved until use. As a control, the nontransduced CD4+ T cells were also prepared using the same method as used above.
HIV infection
CEM-SS cells and CEM-SS cells transduced with MTD3-U3TAR-MazF or MTD3-U3TAR-GFP were infected with HIV-1 IIIB at the different multiplicities of infection (MOIs) of 0.07, 0.0007, and 0.00007. After infection, cells were washed with PBS and subsequently cultured in 10 ml of RPMI 1640 containing 10% FBS. HIV-1 p24 levels in the culture supernatant were calculated using the p24 ELISA kit (PerkinElmer, Waltham, MA). Viable cell numbers were measured using the trypan blue exclusion assay. The doubling time of cells was calculated by logistic regression analysis of each growth curve for the HIV-1 infection sets.
SHIV infection
The cryopreserved cells of the control CD4+ T and MazF-Tmac cells were recovered in GT-T503 medium supplemented with 10% FBS and 200 IU of interleukin-2 and reactivated with anti-monkey-CD3 and anti-human-CD28 monoclonal antibody-conjugated beads at a cell-to-bead ratio of 5:1. After a 6-day incubation, the cells were infected with simian/human immunodeficiency virus (SHIV) 89.6P (Reimann et al., 1996) at the MOI of 0.01 and cultured for 6 more days. SHIV RNA levels in the culture supernatant and intracellular RNAs were determined by using quantitative real-time PCR (Thermal Cycler Dice Real Time System; Takara Bio Inc.) with a set of specific primers designed in the SHIV gag region (Miyake et al., 2006).
Flow cytometry
Flow cytometry was used for the analysis of surface CD4 expression and transduction efficiency. Endogenous expression levels of CD4 in CEM-SS cells and CEM-SS cells transduced with MTD3-U3TAR-MazF were analyzed using phycoerythrin (PE)-labeled anti-human CD4 antibody (Beckman Coulter, Fullerton, CA). Intracellular p24 levels were analyzed using fluorescein isothiocyanate-labeled anti-p24 antibody (Beckman Coulter) after the cells were fixed and permeabilized for flow cytometric analysis.
Gene transfer efficiencies of the retroviral Tat expression vector into CEM-SS cells and CEM-SS cells transduced with MTD3-U3TAR-MazF were analyzed by detecting the ZsGreen marker fluorescence. Immediately before flow cytometry, propidium iodide (PI) was added at the concentration of 100 ng/ml to stain dead cells. Samples were run through a FACSCantoII flow cytometer (BD Biosciences), and data were analyzed using the FACSDiva software (BD Biosciences).
Genomic DNA analysis
Genomic DNA was extracted by phenol/chloroform extraction from CEM-SS cells and CEM-SS cells transduced with MTD3-U3TAR-MazF cells infected with HIV-1 IIIB at the MOI of 0.007. Two different regions of the HIV-1 gag gene (246–467 and 905–1046) were amplified by PCR at 14 days after HIV-1 IIIB infection. As a positive control, genomic DNA was amplified from H9 cells chronically infected with HIV-1 IIIB. Human mitochondrial DNA (mtDNA) was amplified as a control for the PCR.
Co-culture with chronically infected cells
The CEM-SS cell line chronically infected with HIV-1 IIIB (CH-1) was mixed with CEM-SS cells or CEM-SS cells transduced with MTD3-U3TAR-MazF. CH-1 cells were mixed at different ratios of 10, 1, or 0.1%. After 6 and 14 days of infection, intracellular p24 levels were analyzed by flow cytometric analyses.
Results
We first constructed the retroviral vector system in which the gene for MazF was inserted downstream of the HIV-1 TAR sequence (Fig. 1). As the E. coli mazF gene contains nine ACA sequences in its open-reading frame, all of these ACA sequences were first engineered to other MazF-uncleavable sequences without altering the amino acid sequence of MazF to make the mazF mRNA resistant to MazF. The resulting self-inactivating retroviral vector (MTD3-U3TAR-MazF) was used to transduce CD4+ T lymphoid CEM-SS cells to create a system in which MazF induction in CEM-SS cells upon infection with HIV-1 effectively suppressed HIV-1 replication without causing apoptosis of infected T cells. The MTD3 retroviral vector contained an intact 5’ LTR and a mutated 3’ LTR that lacks most of the transcriptional elements present in U3. Cells transduced with the resulting retroviral vector contained the defective LTR at both ends (Yu et al., 1986). The self-inactivating retroviral vector was transiently produced and subsequently transduced into the human T lymphoid line CEM-SS cells, which are highly susceptible to HIV infection. Transduced cells were subjected to G418 selection to obtain drug-resistant populations. A GFP-expressing retroviral vector under the control of HIV-LTR (MTD3-U3TAR-GFP) was also used as a control.
The growth rate of CEM-SS cells transduced with MTD3-U3TAR-MazF was comparable to that of the parental CEM-SS line (Fig. 2A), suggesting that MazF expression was tightly controlled and did not inhibit cell growth. Furthermore, the CD4 levels of MTD3-U3TAR-MazF-transduced CEM-SS cells were identical to those of the parental CEM-SS cells (Fig. 3A).
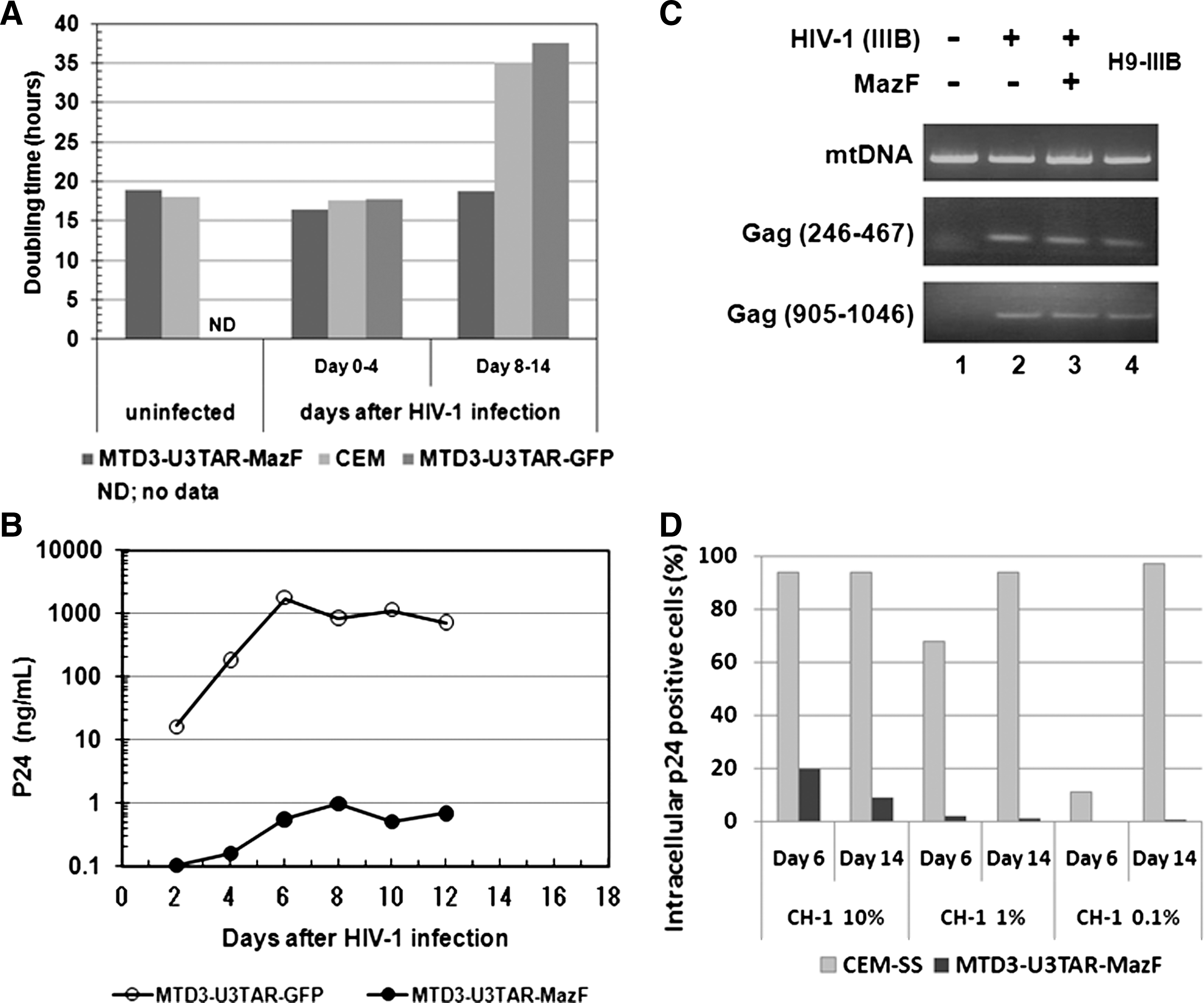
Analysis of MazF-transduced CEM-SS cells after HIV-1 infection. (
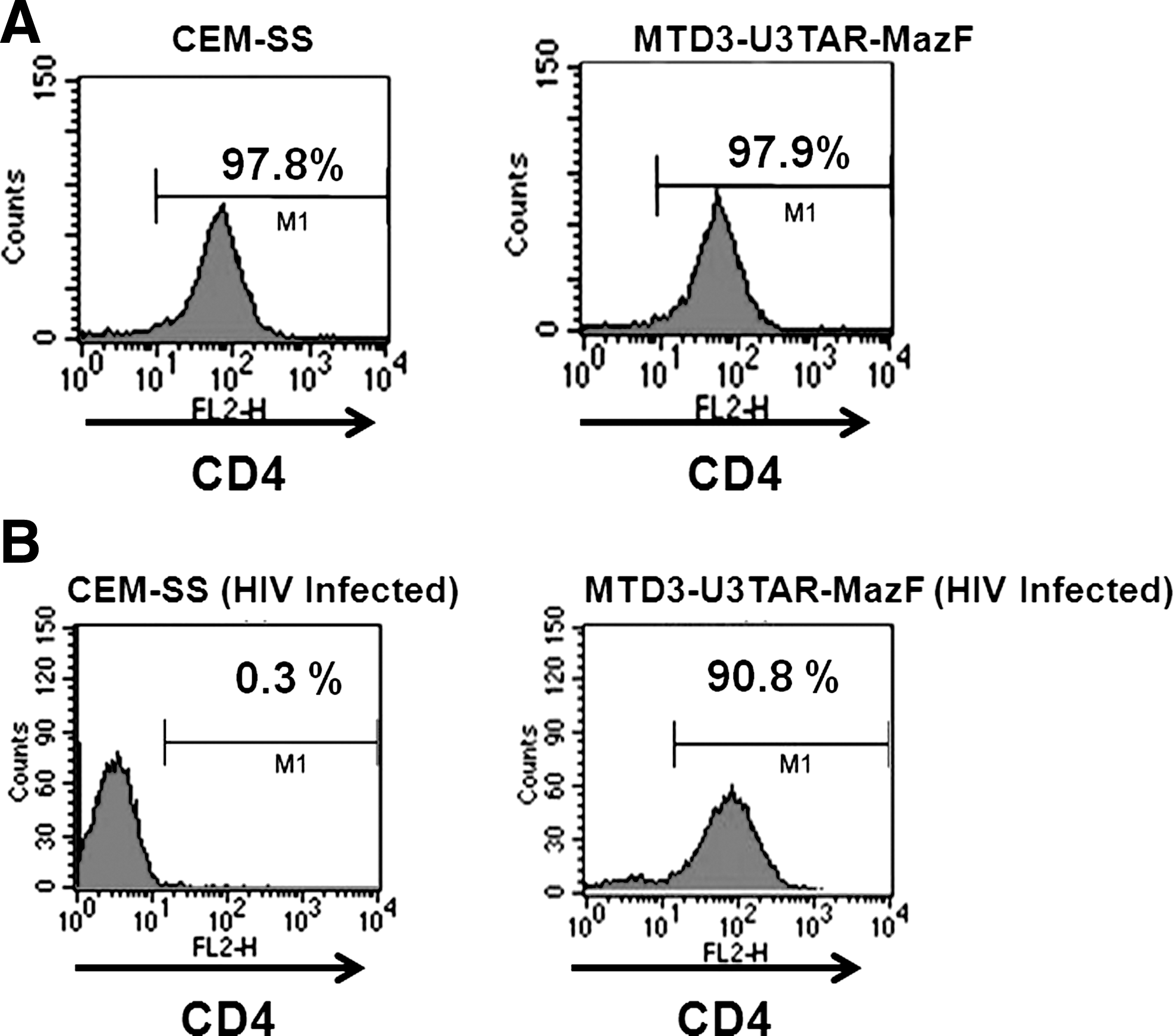
CD4 levels in MazF-transduced cells. (
To investigate the effects of HIV-1 infection, MazF-transduced or GFP-transduced CEM-SS cells were infected with HIV-1 IIIB at different MOIs, specifically 0.07, 0.0007, and 0.00007 (Fig. 4). Levels of the HIV-1 p24 antigen in the culture media were examined 16 days post infection. As shown in Fig. 4, in MazF-transduced CEM-SS cells, HIV-1 replication was effectively suppressed. To more precisely investigate the antiviral effects of MazF, viral production and cell growth were measured every other day after HIV-1 IIIB infection at the MOI of 0.07. As shown in Fig. 2A, in the beginning of the culture from day 0 to day 4, cell growth was similar among CEM-SS cells, MazF-transduced CEM-SS cells, and GFP-transduced CEM-SS cells, as well as uninfected CEM-SS cells. CEM-SS cells harboring the mazF (ACA-less) gene grew at a normal rate throughout the time course of HIV-1 IIIB infection, whereas both GFP-transduced CEM-SS cells and the parental cell line showed aberrant growth rates due to HIV-1 infection in late cultures after day 8 (Fig. 2A). Indeed, a high level of p24 was detected in the GFP-transduced cell populations during the course of infection (Fig. 2B). In the case of MazF-transduced cells, however, levels of p24 were three orders of magnitude lower than those of GFP-transduced cells throughout the experiment (Fig. 2B). Notably, CD4 levels of MazF-transduced cells infected with HIV-1 IIIB were largely unaffected (Fig. 3B). Together with the fact that the HIV-1 IIIB infected cells harboring the mazF gene grew normally (Fig. 2A), these results suggest that HIV-1 IIIB gene expression in the HIV-1-LTR-regulated mazF (ACA-less)-transduced cells is effectively inhibited by blocking HIV-1 replication with little damage to cellular function.
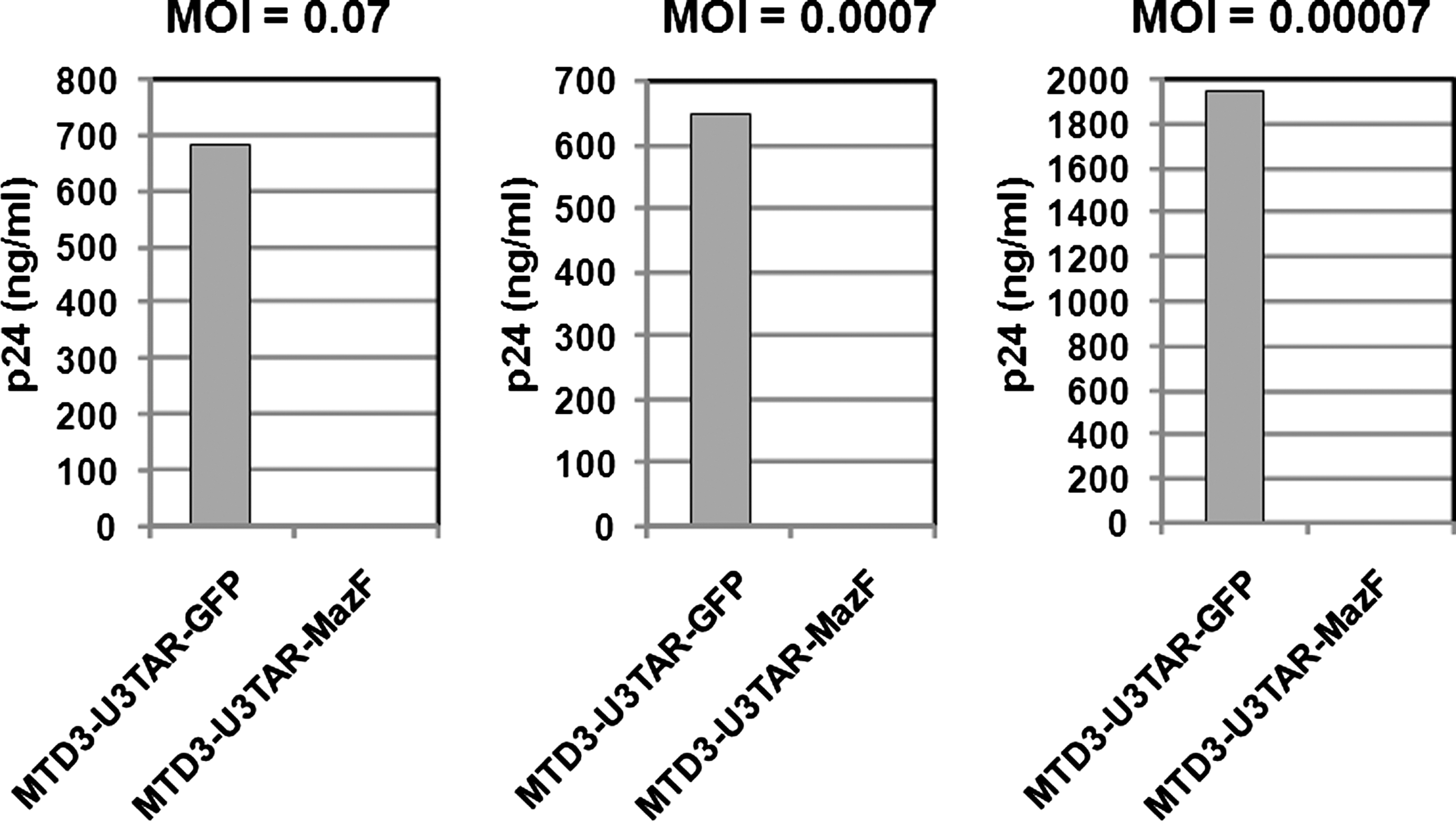
HIV-1 IIIB infection using MazF-transduced CEM-SS cells at different MOIs. Polyclonal cell populations of CEM-SS resulting from gene transduction with retroviral vectors MTD3-U3TAR-MazF or MTD3-U3TAR-GFP were infected with HIV-1 IIIB at different MOIs (0.07, 0.0007, and 0.00007). Sixteen days after infection, HIV-1 p24 levels in the culture supernatant were estimated using the p24 ELISA kit (PerkinElmer). Given the cytopathic effect of HIV-1, the MTD3-U3TAR-GFP cell population showed delayed proliferation after HIV-1 infection in contrast to the MTD3-U3TAR-MazF population. The delay was more pronounced for the high-MOI group (0.07) than for the low-MOI group (0.00007) at later time points. On day 16 post infection, the accumulated cell number of the high-MOI group was threefold lower than that of the low-MOI group, so the difference in HIV-1 p24 levels between the two MOI groups (0.07 and 0.00007) reflects total cell numbers.
Next, we examined if HIV-1 IIIB was integrated into the genome of MazF-transduced CEM-SS cells upon HIV-1 infection. Two different regions of the HIV-1 gag gene were amplified by PCR using genomic DNA 14 days after HIV-1 IIIB infection. As shown in Fig. 2C, both regions of the gag gene were detected in the genome of MazF-transduced CEM-SS cells, which were resistant to HIV-1 replication (lane 3). Similarly, HIV-1 DNA was detected in the genomes of CEM-SS cells (lane 2) and H9-IIIB cells (lane 4) (positive control H9 cells chronically infected with HIV-1 IIIB), whereas no bands were detected in noninfected cells (lane 1). We also established a CEM-SS cell line chronically infected with HIV-1 IIIB (CH-1). When this cell line was mixed with CEM-SS cells or MazF-transduced CEM-SS cells at a ratio of 10, 1, or 0.1%, CEM-SS cells were gradually infected with HIV-1 produced from CH-1 cells (Fig. 2D) and their cell growth was suppressed. Alternatively, MazF-transduced CEM-SS cells showed no growth inhibition (data not shown), indicating that HIV-1 replication was suppressed in MazF-transduced CEM-SS cells. As a result, the culture was eventually taken over by normally growing MazF-transduced CEM-SS cells over the slow-growing CH-1 cells. These data demonstrate that MazF-transduced cells are resistant to HIV-1 IIIB infection by blocking HIV-1 IIIB replication.
To investigate the mazF gene expression and subsequent effects more precisely, CEM-SS cells and CEM-SS cells transduced with MTD3-U3TAR-MazF were infected with the Tat-expressing retroviral vectors, M-LTR-Tat-ZG or H-LTR-Tat-ZG (Fig. 5A). Induction of the mazF gene in CEM-SS cells transduced with MTD3-U3TAR-MazF was monitored by real-time PCR, and the relative ratios were compared with mock infection (Fig. 5B). Infected cells were also subjected to flow cytometry, and both Tat-positive (ZsGreen-positive) cells and dead cells (PI-positive) were monitored (Fig. 5B). As shown in Fig. 5B, strong induction of mazF expression was observed upon constitutive M-LTR-Tat-ZG vector transduction, and there was a significant decline in Tat-positive (ZsGreen-positive) cell population. On the other hand, mazF induction in HIV-LTR-driven Tat expression was lower, and the influence on cell death was also less than by MLV-LTR-driven Tat expression as observed in the PI-positive population. Although these experiments do not directly reflect HIV-1 replication, these data support the hypothesis that only low levels of MazF are expressed upon HIV-1 infection and MazF-positive cells can survive with HIV-1 provirus.

Analysis of MazF induction upon Tat expression. (
As the SIN-based retroviral vector contains the mazF gene in the normal orientation, the mazF gene is expressed from viral mRNA, resulting in the degradation of the viral RNA and thus significantly reducing the viral titer from this vector. On the other hand, when the MazF expression cassette is inserted in the opposite direction from the retroviral genome, the viral titer increased and the gene transfer efficiency was improved more than 10 times (data not shown). To investigate the antiviral effect of the TAR-mazF system in the primary CD4+ T lymphocytes, the reversely orienting MT-MFR-PL2 vector was introduced into rhesus macaque primary CD4+ T cells from two individual monkeys (#14 and #15). The resulting mazF-containing cells were then infected with SIV/HIV-1 chimeric virus SHIV 89.6P. As the SHIV 89.6P harbors HIV-1-derived env, rev, vpu, and tat genes, the TAR-mazF system is expected to function when MazF-Tmac cells are infected with SHIV 89.6P. Indeed, efficient suppression of SHIV 89.6P replication was observed for both primary cell lines, #14 and #15 (Fig. 6A).

Effect of MazF-induction into rhesus macaque primary CD4+ T cells on SHIV 89.6P replication. (
To evaluate further how well the retroviral mazF system is able to suppress viral RNA production, total cellular RNAs were extracted from MazF-Tmac cells to estimate quantitatively the amounts of SHIV RNA, as well as the mRNAs for ribosomal protein L13a (RPL13a, XM_001093017) and β-actin (NM_001033084), by real-time PCR. The relative ratios were normalized by using 18S rRNA (FJ436026), which is protected from MazF cleavage in ribosomes (Shimazu et al., 2007). We obtained similar results in MazF-Tmac cells from both #14 and #15 primary cell lines. Representative results from MazF-Tmac cells from #14 are shown in Fig. 6B, where one can see that SHIV RNA was preferentially cleaved, whereas the cellular mRNAs were not affected. These results clearly demonstrate that MazF induction from the Tat system upon SHIV 89.6P infection leads to severe defect in maintaining SHIV 89.6P RNA but does not affect cellular mRNAs in SHIV-infected CD4+ T cells.
Discussion
This study demonstrates the distinct feasibility of RNase-based strategies for gene therapy. RNase-based strategies may be preferred over RNA-based strategies for HIV therapy, because RNases cleave HIV-RNA to cause permanent damage to HIV RNA function. Additionally, as RNases function as an enzymatic catalyst, they are required only at low concentrations in the cells to effectively block HIV proliferation. In the present study, the gene for MazF, an ACA-specific mRNA interferase, was engineered under the HIV-1 LTR promoter and inserted in the genome of the CD4+ T lymphoid cells so that MazF is expected to be produced only when the cells are infected with HIV-1 to produce the Tat protein. We demonstrated that mazF-Tmac cells indeed acquired resistance against SHIV replication, but cell growth was not inhibited after SHIV infection (data not shown), indicating that cellular mRNAs were not significantly affected. Notably, MazF was also able to function against the expression of SHIV proviral genome, because the production of SHIV in the culture supernatant was dramatically reduced.
Acquisition of HIV-1 resistance, and more remarkably the ability of MazF-transduced cells to suppress HIV-1 replication, may be explained as follows: Upon HIV-1 infection, Tat expression is first induced from the HIV-1 proviral genome. Tat then triggers the transcription of the mazF gene under the LTR promoter, as well as the full-length HIV proviral genome. The resulting induction of MazF expression leads to the cleavage of newly emerged HIV-1 mRNAs so that Tat protein synthesis is no longer sustainable. However, it is important to note that HIV-1 infection does not hamper cell growth and that the HIV-1 provirus genome is retained in the MazF-transduced cells. Therefore, the cellular level of Tat appears to be maintained at a very low level so that the level of MazF induction is also kept very low enough to cleave HIV-1 mRNAs, but not cellular mRNAs. Depending on the integration site and proviral copy number, there might be some MazF-transduced cells that were not resistant to HIV-1 replication. However, these cells could not survive due to HIV-1-induced cell death.
In mammals, virus infection is known to activate the interferon response to induce RNaseL, which mediates degradation of 28S and 18S ribosomal RNAs. This results in inhibition of protein synthesis as part of the host antiviral response (Silverman, 2003). An amphibian ribonuclease, Onconase, is able to inhibit protein synthesis in mammalian cells and has been used as a protein drug. When it was added to the culture media of H9 cells persistently infected with HIV-1, HIV-1 replication was inhibited without blocking cell growth, as degradations of 18S and 28S rRNAs and cellular mRNAs were prevented (Saxena et al., 1996). MazF induction in mammalian cells has shown to cause apoptotic cell death as a result of degradation of cellular mRNAs (Shimazu et al., 2007). However, in the present study, MazF expression induced by HIV-1 Tat appears to be maintained at very low levels, just enough to cleave HIV-1 RNA but not cellular mRNAs, so that cells were able to grow normally. MazF expression may be autoregulated in the cell in such a way that when Tat-induced MazF eliminates invading HIV-1 RNA, Tat expression from the HIV-1 provirus is simultaneously stopped, resulting in simultaneous arrest of MazF production to recover normal cellular functions.
Targeting HIV RNA as a therapeutic strategy using antisense RNA (Levine et al., 2006), ribonucleases (Agarwal et al., 2006), and RNA interference (RNAi) technology (Morris and Rossi, 2004) has been attempted. However, the use of antisense RNA and RNAi technology has not been effective as an anti-HIV technology, as HIV can easily circumvent these RNA inhibitors by creating mutations at the target sequence regions (Lee and Rossi, 2004). On the other hand, the present strategy using MazF targets abundant ACA sequences in HIV-1 RNA (>240), so that it is not possible for HIV-1 to escape from MazF attack by mutations. Furthermore, because MazF has no homology to any mammalian ribonucleases, MazF mRNA interferase activity cannot be inhibited by ribonuclease inhibitors existing in mammalian cells.
In summary, the use of MazF appears to be a novel and highly effective tool for anti-HIV gene therapy. It is effectively able to suppress HIV-1 replication, preventing the emergence of mutated HIV-1. Importantly, MazF induction by invading HIV-1 shows little toxicity to host cells while it efficiently suppresses HIV-1 replication. Specific inhibition of HIV-1 replication by MazF without affecting cell growth is the key feature of MazF-based HIV-1 gene therapy. This may be the first step for RNase-based HIV-1 gene therapy with efficacy in vitro. The feasibility of the MazF-based ex vivo gene therapy may be verified using autologous CD4+ T lymphocytes from HIV-1 patients. To use our mazF vector system for gene therapy, its safety has to be critically evaluated and it should not have any negative impacts on T-cell function. For example, it needs to be shown that there is no alteration in the secretion of functionally important cytokines even though it was observed that MazF expression in HIV-infected CD4+ T cells does not inhibit cell growth. We are currently addressing this question.
Footnotes
Acknowledgments
The authors thank Dr. Keith A. Reimann of Harvard Medical School and Dr. Tomoyuki Miura of Kyoto University for providing the SHIV 89.6P. The authors also thank Dr. Koich Inoue of Takara Bio Inc. for his critical reading of the manuscript.
Author Disclosure Statement
No competing financial interests exist.