
Abstract
The search for the ideal stem cell gene therapy vector continues as recognized problems persist. Although recombinant adeno-associated virus serotype 2 (rAAV2) mediates gene transfer into hematopoietic stem cells, identified restrictions to transgene expression reduce overall efficiency. Studies have shown that transduction efficiencies are significantly improved by preventing early proteasomal degradation after mutation of surface-exposed tyrosine residues on the capsid to phenylalanine. Here, we report that transduction of human cord blood CD34+ stem cells by tyrosine-modified rAAV2 is significantly enhanced both in vitro and in vivo. Serial long-term in vivo bioluminescent imaging of immune-deficient recipients after xenotransplantation of CD34+ cells transduced with tyrosine-modified rAAV2-luciferase revealed that modification of rAAV2 capsids led to a significant increase in the transduction of human CD34+ cells, without adversely affecting engraftment capacity, or the ability to undergo multilineage differentiation and self-renewal. Together with observations of sustained high-level transgene expression in vivo and efficient persistence of rAAV genomes in human hematopoietic cells, these results suggest that, because of their ability to bypass restrictions to transduction, tyrosine-modified rAAV vectors, particularly Y500F, Y700F, Y444F, and Y704F, represent highly promising candidates for therapeutic evaluation for diseases of human hematopoietic stem cells.
Introduction
Despite much effort focused on genetic modification of the hematopoietic system, the first major target of gene therapy, to date, there are few instances of therapeutic success (Biffi and Cesani, 2008). This is due to the recognized inability of most viral vectors to efficiently transduce quiescent, nondividing hematopoietic stem cells (HSCs) (Miller et al., 1990) as well as safety concerns arising from insertional oncogenesis (Hacein-Bey-Abina et al., 2003; McCormack and Rabbitts, 2004). However, we and others have successfully demonstrated stable gene transfer to both murine and human HSCs by rAAV (Zhou et al., 1993; Einerhand et al., 1995; Fisher-Adams et al., 1996; Ponnazhagan et al., 1997; Chatterjee et al., 1999; Srivastava, 2004; Santat et al., 2005; Paz et al., 2007). In addition, we have demonstrated chromosomal integration of rAAV in human CD34+ HSCs (Fisher-Adams et al., 1996; Chatterjee et al., 1999; Santat et al., 2005; Han et al., 2008) and efficient transduction of primitive, pluripotent, self-renewing human HSCs capable of supporting primary and secondary multilineage engraftment in immune-deficient nonobese diabetic/severe combined immunodeficiency mice (Santat et al., 2005). Subsequently, we showed that transduction of primitive HSCs capable of supporting serial engraftment was attributable to the propensity of rAAV to efficiently transduce primitive, quiescent CD34+CD38– cells residing in G0 (Paz et al., 2007). Despite several reports of successful rAAV-mediated gene transfer into human HSCs in vitro and in murine and nonhuman primate HSCs in vivo, controversy regarding the utility of rAAV for HSC transduction still persists. These discrepancies arose primarily from short-term in vitro studies that assessed transduction by expression profiling and are attributable to the identified restrictions to transgene expression from rAAV2, including viral uncoating (Zhong et al., 2004b), intracellular trafficking (Zhong et al., 2008a), and nuclear transport and second-strand synthesis (Zhong et al., 2007). Tyrosine-modified AAV capsids, which directly circumvent proteasomal degradation and intracellular trafficking, thus constitute valuable tools to assess rAAV-mediated gene transfer into human HSCs.
In this study, we tested the hypothesis that tyrosine-modified rAAV2 will efficiently transduce human cord blood (CB) CD34+ HSCs, capable of long-term in vivo engraftment and stable transgene expression in a xenotransplantation model. Our results revealed that transduction of human HSCs with tyrosine-modified rAAV2 is significantly enhanced as compared with wild-type rAAV2, resulting in sustained transgene expression and higher genome copy frequencies.
Materials and Methods
CD34+ cell isolation
Umbilical cord blood was obtained according to an institutional review board-approved protocol. Low-density mononuclear cells were isolated by Ficoll-Paque (GE Healthcare, Piscataway, NJ) centrifugation. The CB CD34+ mononuclear cells were isolated by immunomagnetic selection, using CD34+ isolation kits (Miltenyi Biotech, Auburn, CA) as per the manufacturer's directions. CD34+ cells were passed through two successive positive selection columns to increase the CD34+ cell purity. Purity was previously assessed to be 96–98% by flow cytometry after labeling with fluorescein isothiocyanate (FITC)-conjugated hematopoietic progenitor cell antigen (HPCA)-2.
Virus packaging
Wild-type and tyrosine-modified rAAV vectors were packaged in HEK-293 cells as previously described (Chatterjee et al., 1992; Chatterjee and Wong, 1993; Santat et al., 2005). The encapsidated rAAV genomes encoded self-complementary enhanced green fluorescent protein (EGFP) and single-stranded luciferase. The integrity of the ITRs was confirmed before packaging. Titers of packaged rAAV were determined by real-time TaqMan amplification assay with vector-specific primers and probes (Santat et al., 2005).
Transduction of human cord blood-derived CD34+ cells
Purified CB CD34+ cells from up to five samples were pooled and transduced with rAAV at a multiplicity of infection (MOI) of 20,000 in Iscove's modified Dulbecco's medium (IMDM; Irvine Scientific, Santa Ana, CA) containing 20% fetal calf serum (FCS; Invitrogen, Carlsbad, CA), streptomycin (100 μg/ml) and penicillin (100 U/ml) (Invitrogen), 2 mM
Xenotransplantation of CD34+ cells
All animal care and experiments were performed under protocols approved by the Institutional Animal Care and Use Committee (City of Hope, Duarte, CA). Transplantations were performed as described (Santat et al., 2005). Briefly, 6- to 8-week-old male NOD.CB17-Prkdcscid /NCrCrl (NOD/SCID) mice (Charles River, Wilmington, MA) were maintained in a specific pathogen-free facility at the Animal Resources Center (City of Hope). Mice were administered sulfamethoxazole and trimethoprim oral pediatric antibiotic (Hi-Tech Pharmacal, Amityville, NY) (10 ml/500 ml H2O) for at least 48 hr before transplantation. Mice were irradiated with a sublethal dose of 350 cGy from a 137Cs source and allowed to recover for a minimum of 4 hr before transplantation. For the majority of transplants, 7 × 105–1.1 × 106 transduced CD34+ cells were infused via the tail vein in a total volume of 150–300 μl. Three to five mice were transplanted per vector group. Bone marrow from femurs and tibias, and the spleen were harvested for analysis from each mouse at 16 to 22 weeks posttransplantation.
Serial bioluminescence analysis of luciferase expression
Luciferase expression in xenografted mice was monitored by serial biweekly bioluminescent imaging with a Xenogen in vivo imaging system (Caliper Life Sciences, Hopkinton, MA). Mice were anesthetized with oxygen containing 4% isoflurane (Phoenix Pharmaceuticals, St. Joseph, MO) for induction, and 2.5% for maintenance. Luciferin (Caliper Life Sciences) was injected intraperitoneally at a dose of 0.15 mg/g mouse weight. Photons were accumulated over a 5 min exposure from the ventral aspect, 10 min postinjection. Living Image 3.0 software (Caliper Life Sciences) was used to calculate light emission.
Flow cytometric analysis
In vitro expression in 20,000 cells was analyzed 24 hr after rAAV-EGFP transduction. Cells were washed in phosphate-buffered saline (PBS) (Mediatech, Manassas, VA) containing 5% FCS and 0.1% sodium azide before analysis on a CyAn ADP flow cytometer (Dako, Glostrup, Denmark). Specific EGFP was quantified after the subtraction of autofluorescence. In vivo engraftment of human cells in both the bone marrow and spleen of xenografted mice was analyzed as described previously (Santat et al., 2005). Lineage distribution was assessed in bone marrow and spleen cell suspensions after staining with human specific antibodies: FITC-conjugated anti-CD45; FITC- or allophycocyanin (APC)-conjugated anti-CD34; APC-conjugated anti-CD33, anti-CD14, and anti-glycophorin A; phycoerythrin (PE)-conjugated anti-CD19; and FITC-, PE- and APC-conjugated IgG controls (BD Biosciences, Mountain View, CA). Bone marrow lineages were sorted by fluorescence-activated cell sorting (FACS) using FITC–CD34, APC–CD33, PE–CD19, and APC–glycophorin A, as well as the appropriate controls. FITC and PE fluorescence was excited with a 488 nm laser, and APC fluorescence was excited with a 670 nm laser. Flow cytometry data were then analyzed for specific populations with FlowJo software (Treestar, Ashland, OR).
rAAV frequency detention
rAAV2 frequencies were detected by quantitative real-time PCR with vector-specific primers and probe and a 7900HT sequence detection system (Applied Biosystems, Foster City, CA). High molecular weight DNA was extracted from human lineages isolated from the marrow of mice transplanted with single-stranded luciferase (ssLuc)-transduced HSCs according to standard methods. Vector-specific sequences were amplified by real-time TaqMan PCR, using primers Luc1 (5′-AACTGCACAAGGCCATGAAGA-3′) and Luc2 (5′-CTCAAAGTATTCAGCATAGGTGATGTC-3′), and were detected with the TaqMan probe 5′-FAM-TTGCCTTCACTGATGCTCACATTGAGGT-TAMRA-3′. Samples were also evaluated for the single-copy human gene ApoB, which served to quantitate human cell equivalents and as a template integrity control (Santat et al., 2005).
Results
Transduction of human CD34+ cells in vitro by tyrosine-modified rAAV2
We initially tested the capacity of tyrosine-modified rAAV2 capsids to transduce human CB-derived CD34+ cells in vitro as compared with wild-type rAAV2. Freshly isolated, highly purified CD34+ cells pooled from three different CB samples were transduced with wild-type or tyrosine-modified rAAV2 vectors encoding a self-complementary EGFP (scEGFP). The scEGFP vector was used to circumvent limitations associated with second-strand synthesis. EGFP expression assessed by flow cytometric analysis at 24 hr posttransduction (Fig. 1) revealed that tyrosine-modified capsids Y700F, Y704F, Y444F, and Y500F showed higher levels of EGFP expression than wild-type AAV2. Y700F and Y704F showed the highest levels of transduction at 43 and 31%, respectively. Y444F and Y500F showed the next highest transduction efficiencies at 25 and 24% EGFP expression, respectively. Y730F and Y252F supported the lowest EGFP expression at levels comparable to wild-type AAV2 or lower. These results suggest that modification of specific surface-exposed tyrosine on AAV2 capsids markedly enhanced transduction of human CD34+ HSCs in vitro.
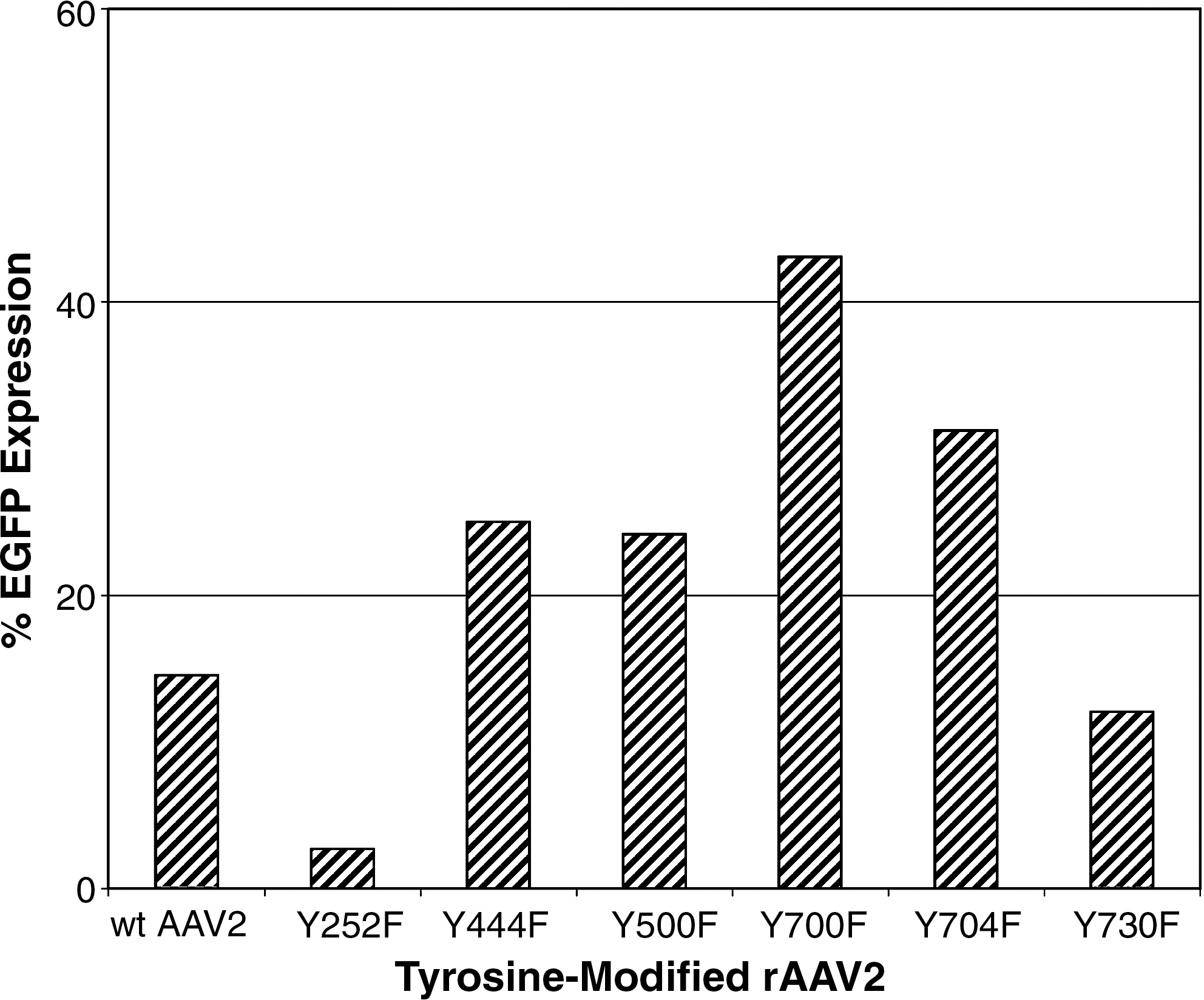
In vitro transgene expression in human cord blood CD34+ cells after transduction with AAV2-self-complementary EGFP packaged in wild-type and tyrosine-modified AAV2 capsids. EGFP expression was determined by flow cytometric analysis. Data shown represent CD34+ cells pooled from three separate CB samples per group.
In vivo engraftment of transduced human CD34+ cells
To determine whether cord blood CD34+ HSCs transduced with tyrosine-modified rAAV2 encoding either scEGFP or ssLuc could support long-term multilineage engraftment in immune-deficient NOD/SCID mice, we evaluated human hematopoietic engraftment 16–22 weeks posttransplantation in the bone marrow of xenograft recipients (Fig. 2A). Human cell engraftment in the bone marrow ranged from 0.5 to 86% (median, 43.37%; n = 40), as determined by the frequency of human CD45+ cells. Engraftment was found to be stable throughout the period of analysis, up to 22 weeks posttransplantation, suggesting a lack of toxicity associated with transduction of CD34+ cells with tyrosine-modified rAAV2, comparable to that observed with wild-type rAAV2 (Santat et al., 2005). In addition, we conclude that CD34+ cells transduced with tyrosine-modified rAAV2 were capable of supporting long-term human hematopoietic engraftment.
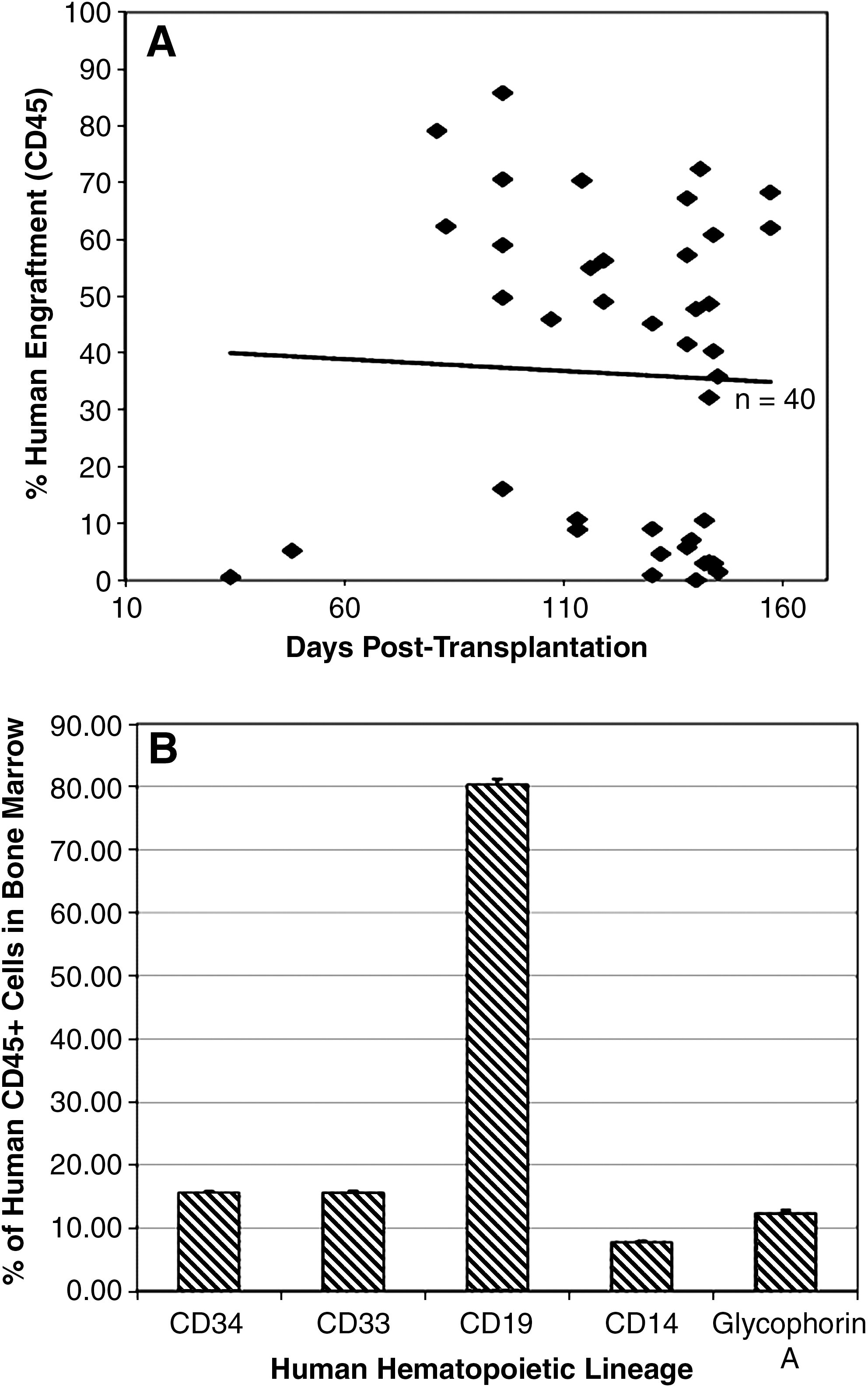
Engraftment of human CB CD34+ cells transduced with tyrosine-modified AAV2. Engraftment was determined in NOD/SCID mice transplanted with tyrosine-modified rAAV2 encoding either self-complementary enhanced green fluorescent protein or single-stranded luciferase. Mice were transplanted with transduced CD34+ cells pooled from one to five blood samples. (
The presence of differentiated human B lymphoid cells (CD19+), erythroid cells (glycophorin A+), and myeloid cells (CD14+ and CD33+) in the bone marrow up to 22 weeks posttransplantation indicated that the highly purified input human CD34+ cells were capable of differentiation after transplantation (Fig. 2B). The continued presence of CD34+ stem/progenitor cells (15.54 ± 6.30%) throughout the study indicated the ability of transplanted CD34+ cells to persist and/or self-renew in vivo. CD19+ B cells comprised the most frequent human cell subpopulation in the bone marrow (80.29 ± 19.70%). CD33+ and CD14+ myeloid cells and glycophorin A+ erythroid cells accounted for 15.55 ± 8.11, 7.69 ± 3.49, and 12.30 ± 8.46% of bone marrow cells, respectively (Fig. 2B). Importantly, no pathology or toxicity was associated with the transplantation or engraftment of CD34+ cells transduced with tyrosine-modified rAAV2.
Analysis of the spleen in transplanted mice indicated that human CD45+ cells were also present (range, 0.2–47.5%; n = 40), representing either direct homing or trafficking from the marrow. CD19+ B cells constituted most (89.7 ± 16.2%) of the splenic human subpopulation. These results indicate the ability of transduced, transplanted human CD34+ cells to safely engraft, undergo multilineage differentiation, and possibly traffic in vivo.
Serial bioluminescence analysis of in vivo transgene expression after transplantation of CD34+ HSCs transduced with tyrosine-modified rAAV2
To determine which capsid best supported long-term in vivo transgene expression, we transduced CD34+ cells with tyrosine-modified rAAV2 encoding the firefly luciferase gene under the control of the cytomegalovirus (CMV) enhancer/chicken β-actin
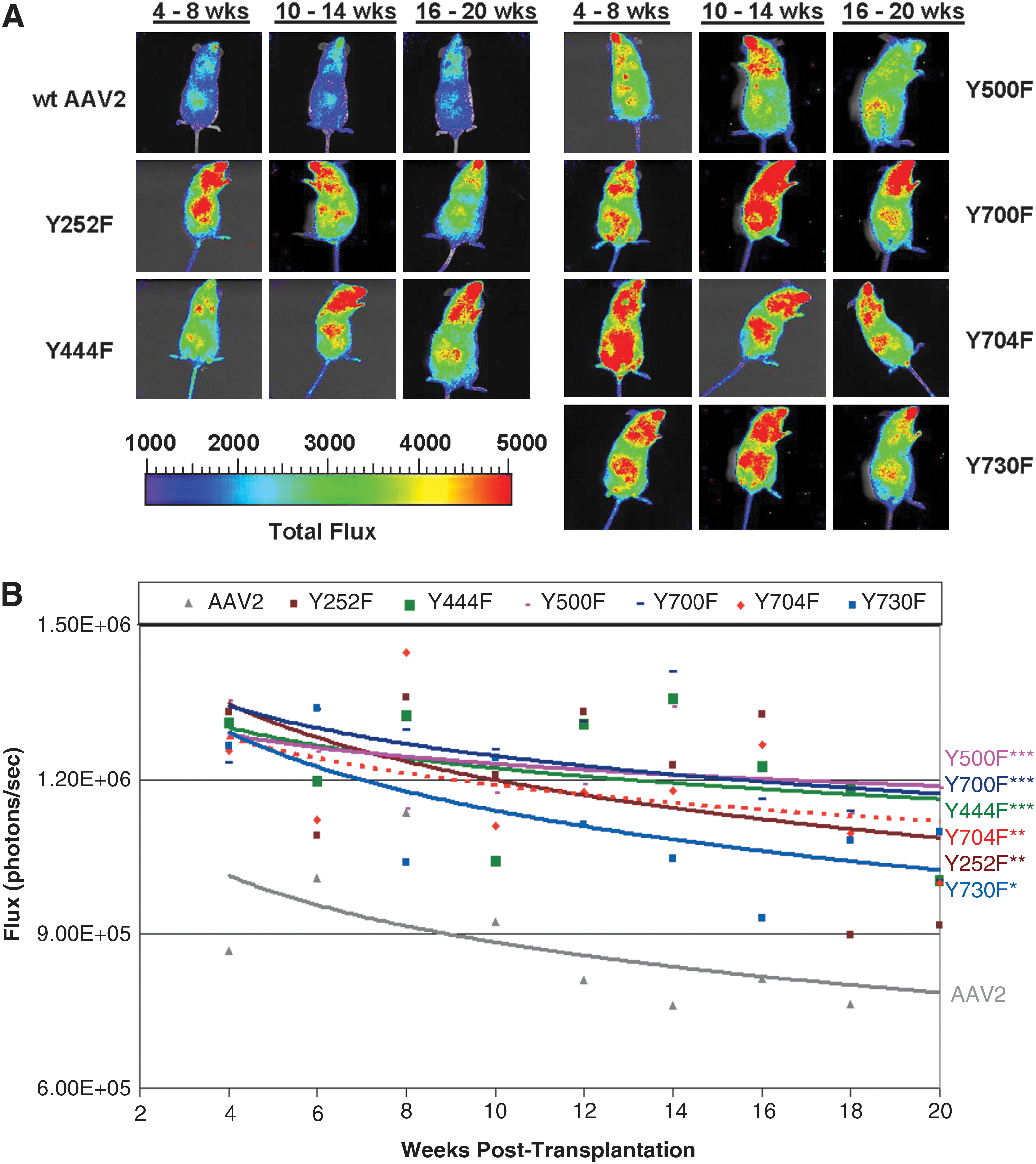
(
Serial quantitation of whole body bioluminescence in xenograft recipients of AAV-luciferase-transduced human HSCs revealed long-term in vivo transgene expression from tyrosine-modified rAAV2 capsids. Figure 3B shows regression analysis of the mean luciferase expression for each capsid type from 4 to 20 weeks posttransplantation. All six tyrosine mutants analyzed showed significantly enhanced levels of luciferase expression as compared with wild-type rAAV2 controls (p < 0.001, for all groups except Y730F). Y500F supported the brightest expression, followed by Y700F, Y444F, and Y704F. Although mice transplanted with Y730F-transduced HSCs showed lower levels of luciferase expression, they were still brighter than wild-type rAAV2 (p < 0.005). Interestingly, transgene expression was found to be remarkably stable over time in the Y500F- and Y444F-transduced groups. The slight level of decline in luciferase expression observed over time for the other groups was most likely due to a possible correlative decline in human cell engraftment. These results demonstrate that human CD34+ cells transduced with tyrosine-modified rAAV2 engraft efficiently, and support long-term stable transgene expression to a significantly higher level than wild-type rAAV2. Importantly, no toxicity or pathology was detected in any of the groups evaluated.
Frequency of rAAV genome copies in human hematopoietic cells in xenograft recipients
We next confirmed gene transfer into human HSCs with tyrosine-modified rAAV by an unrelated method. The frequency of the rAAV genomes was quantitated in flow-sorted human hematopoietic lineages from the marrow of xenograft recipients at 18 to 22 weeks posttransplantation, using a real-time TaqMan amplification assay with vector-specific primers and probes. The frequency of rAAV-transduced cells in each lineage was quantitated by comparing the copy number of rAAV genomes with the total number of human cell equivalents as determined by the single-copy human gene ApoB (Table 1). Every sample analyzed showed the presence of rAAV genomes. Y500F displayed the highest rAAV genome copy frequency in all lineages except for CD19+ B cells, for which it was the second highest. Y444F displayed the highest CD19+ cell marking. The highest level of stable rAAV marking of long-term engrafted human hematopoietic lineages was observed with Y500F, followed by Y444F and Y700F. The frequency of wild-type AAV2-transduced cells, as reported previously (Santat et al., 2005), was included for comparison.
Abbreviations: GlycoA, glycophorin A; ND, not done; rAAV, recombinant adeno-associated virus; wtAAV2, wild type AAV2.
Copies per cell.
Of all the human lineages harvested from transplant recipients, the long-term rAAV genome copy frequency was found to be the highest in erythroid and CD34+ cells at 18–22 weeks posttransplantation. Marking of CD34+ cells was most efficient with Y500F, Y444F, and Y704F, which marked 100, 86, and 84% of all human long-term engrafted CD34+ cells, respectively. One hundred percent of glycophorin A+ erythroid cells were found to be marked by Y500F and Y700F. These results suggest that tyrosine-modified rAAV2, especially Y500F, are excellent candidates for mediating stable gene transfer into primitive human CD34+ stem cells and that erythroid cells derived from these HSCs maintain the rAAV genome in a stable fashion. Moreover, these results further confirm that CB CD34+ cells transduced with tyrosine-modified rAAV2 support long-term engraftment and are capable of long-term lymphomyeloid reconstitution in vivo, and that transduced CD34+ cells are capable of persistence and/or self-renewal long-term posttransplantation.
Discussion
Due to their many recognized advantages, rAAV have become the vector of choice for an increasing number of human gene therapy applications as evidence of clinical efficacy rapidly mounts (Manno et al., 2003; Flotte et al., 2004; Kaplitt et al., 2007; Bainbridge et al., 2008; Brantly et al., 2009; Cideciyan et al., 2009). However, high-level transgene expression from rAAV2 has been shown to be restricted by a series of well-defined limitations in many tissues (Zhong et al., 2007, 2008a). These limitations are particularly evident in primitive, quiescent HSCs and have thus far represented significant impediments to the clinical application of this promising gene therapeutic approach for hematological diseases. The Srivastava group recently demonstrated that rational mutagenesis of surface-exposed tyrosine residues to phenylalanine on the AAV2 capsid reduced phosphorylation and, consequently, ubiquitination and proteasomal degradation of capsids, thus increasing transduction efficiencies dramatically (Jayandharan et al., 2008; Zhong et al., 2008b; Petrs-Silva et al., 2009). Here we show that tyrosine-modified rAAV2 vectors transduce human CB-derived CD34+ HSCs significantly better than wild-type rAAV2. HSCs transduced with these modified vectors engraft efficiently, are capable of persisting and/or self-renewing, as well as undergoing multilineage differentiation. In addition, xenotransplantation of transduced HSCs results in sustained, augmented transgene expression and long-term, stable vector genome persistence in vivo.
The rapid onset of high-level transgene expression after in vitro transduction of CD34+ cells with tyrosine-modified rAAV2, as compared with wild-type rAAV2, indicated that modification of surface-exposed tyrosine residues dramatically increased transduction efficiencies of human HSCs, suggesting that tyrosine phosphorylation, ubiquitination, and proteasomal degradation play a significant role in limiting transgene expression from rAAV2 in CD34+ cells. The striking enhancement of EGFP expression supported by Y704F, Y444F, Y700F, and Y500F suggested that these capsids in particular successfully avoided degradation and increased intracellular trafficking and nuclear entry. Interestingly, of these, Y444F and Y500F also displayed improved transduction of the mouse liver (Markusic et al., 2010). Differences in the transduction efficiency of mouse liver and human HSCs by Y730F may be attributable to cell-specific differences in handling and processing of AAV capsids.
Importantly, CD34+ human HSCs transduced with tyrosine-modified rAAV2 engrafted efficiently in immune-deficient NOD/SCID mice at levels comparable to that observed with wild-type AAV2 (Santat et al., 2005), suggesting that tyrosine modification had no adverse effects on the ability of these cells to home to stem cell niches, engraft, and give rise to human hematopoiesis. The detection of all expected human lineages indicated that tyrosine-modified rAAV2 did not alter human hematopoietic differentiation.
Serial bioluminescent imaging analysis of the recipients of human CB CD34+ cells transduced with tyrosine-modified rAAV encoding the firefly luciferase gene allowed a linear evaluation of transgene expression after stem cell xenotransplantation in the same recipients over time, eliminating mouse-to-mouse variability. Each of the six tyrosine mutants analyzed showed significant enhancement of luciferase expression in vivo as compared with wild-type rAAV2. In concordance with the results of the in vitro transduction assay, this was especially marked for Y500F, Y444F, Y700F, and Y704F, suggesting that these tyrosine residues are likely the most critical for ubiquitination and the subsequent proteasomal degradation processes in CD34+ cells. As compared with the wild-type rAAV2 control, whole body luciferase expression both initiated at a higher intensity and persisted at significantly elevated levels for up to 22 weeks posttransplantation, the end of the experiment. This sustained high-level expression suggests that tyrosine-modified vectors would be well suited for long-term expression of therapeutic genes in vivo.
Importantly, the rAAV genomes were readily detected in all human hematopoietic lineages harvested from xenotransplant recipients at 18–22 weeks posttransplantation, including in differentiated B lymphocytes, myeloid cells, erythroid cells, and CD34+ stem and progenitor cells. The presence of rAAV genomes in these populations indicated that the mature progeny of transduced CD34+ cells stably retained the vector genomes throughout the differentiation process, which involves multiple rounds of cell division. In addition, the relatively high frequencies of rAAV genomes in CD34+ cells purified from the marrow of xenograft recipients long-term suggested that rAAV-transduced CD34+ cells had the capacity to either persist long-term in vivo and/or self-renew. Importantly, taken together these results demonstrate that CD34+ HSCs transduced with tyrosine-modified rAAV are capable of multilineage differentiation and self-renewal without concomitant loss of rAAV genomes.
The overall frequency of rAAV genomes persisting in human hematopoietic subpopulations purified from the marrow of xenograft recipients was highest for the erythroid lineage, followed by CD34+ stem cells and CD33+ myeloid cells. Interestingly, a similar pattern of rAAV2 genome distribution were observed in wild-type rAAV-transduced human xenografts, with stem/progenitor and myeloid cells showing higher levels of gene marking than CD19+ B lymphoid cells (Santat et al., 2005). Notably, the higher frequency of rAAV genomes in erythroid cells suggests that tyrosine-modified rAAV vectors may be ideally suited for gene therapy of hemoglobinopathies. Efficient gene marking by Y500F rAAV2 specifically, coupled with the correlative high-level long-term expression, suggests that it may be especially well suited for therapeutic use in primitive human HSCs.
The frequency of rAAV genomes in human lineages derived from cells transduced with tyrosine-modified rAAV2 ranged from approximately 10- to 100-fold higher than that observed with wild-type rAAV2 capsids (Santat et al., 2005). This is directly attributable to the increased intracellular persistence of a larger number of rAAV genomes after inhibition of phosphorylation, ubiquitination, and proteasomal degradation of tyrosine-modified capsids. This in turn likely leads to greater intranuclear transport and long-term persistence in HSCs. Dividing and differentiating HSCs are expected to lose episomal rAAV. Thus, for rAAV genomes to persist at relatively high frequencies in a stable fashion long-term in the dividing and differentiating progeny of transduced HSCs, they would most likely exist in a chromosomally integrated form. It is also probable that the higher frequency of rAAV genomes packaged in tyrosine-modified capsids, represents more efficient chromosomal integration in stem cells compared with those packaged in wild-type AAV capsids, possibly due to alterations in processes downstream to signal transduction events initiated by tyrosine phosphorylation, and that may represent events associated with chromosomal integration.
This study clearly demonstrates that tyrosine modification of rAAV2 capsids leads to a significant increase in the transduction of human CB CD34+ HSCs, without adversely affecting engraftment capacity, or their ability to undergo multilineage differentiation and self-renewal. Together with our observations of long-term, sustained high-level transgene expression in vivo and efficient persistence of rAAV genomes in HSCs and their progeny, these results suggest that because of their ability to bypass restrictions to transduction, tyrosine-modified rAAV vectors represent highly promising candidates for therapeutic evaluation, particularly for diseases of stem cells and the erythroid and myeloid lineages. The increased transduction efficiency of these rAAV2 capsids with tyrosine modifications suggests that multiple mutations in tandem may further improve the efficiency of gene transfer.
Footnotes
Acknowledgments
This research was supported in part by NHLBI grant R01HL087285 (to S.C.) and grant 8187368876 (Roche Foundation for Anemia Research), and by a grant from the Fanconi Anemia Research Fund (to L.Z.). City of Hope (COH) core facilities used for this study were supported by Cancer Center grant P30CA33572 from the NCI. The authors thank John A. Zaia and Stephen J. Forman (COH) for support; Cameron Mroske (COH) for thoughtful discussions; Valerie Gonzales (COH) for secretarial help; Elizabeth Garcia and Lucy Brown for technical advice. The following COH Cancer Center cores were used in this study: the Animal Resources Center, Flow Cytometry Core, In Vivo Imaging Core, Sequencing Cores, and the Cloning Laboratory.
Author Disclosure Statement
No competing financial interests exist.