
Abstract
Conditionally replicative adenoviruses (CRAds) are widely used for cancer biotherapy and show a significant growth-suppressing effect on many types of cancer. However, it was reported that breast cancer was highly resistant to the infection of traditionally used adenovirus of serotype 5 (Ad5)-based CRAds. Although partial substitution of the fiber protein of replication-deficient Ad5 with that of adenovirus of serotype 35 (Ad35) facilitated infection of breast cancer cells by adenoviral vectors, it is still unknown whether this modification can improve CRAds in their tumor-eliminating capacity. We generated a 5/35 fiber-modified CRAd with a p53 cDNA construct and investigated whether this alteration in fiber region can make CRAds suppress the growth of breast cancer more effectively. Our data reinforced the proposal that 5/35-modified fiber conferred higher adenovirus infectivity for breast cancer cells than natural Ad5 fiber. Interestingly, 5/35 fiber-modified CRAd replicated more efficiently in breast cancer cells than Ad5-based CRAd. We also found 5/35 fiber-modified CRAd mediated higher expression of p53 in breast cancer cells. In vitro, 5/35 fiber-modified CRAd eliminated breast cancer cells more efficiently. Growth of xenograft tumors in nude mice was also significantly retarded by 5/35 fiber-modified CRAd. The 5/35 fiber-modified CRAd suppressed the growth of breast cancer cells more effectively than Ad5-based CRAd, both in vitro and in vivo. Thus CRAd with 5/35 hybrid fiber may be a promising vector for breast cancer treatment.
Introduction
Conditionally replicative adenoviruses (CRAds), also called oncolytic adenovirus, can efficiently replicate and produce progeny in cancer cells of various types, but not in normal cells. In other words, CRAds exhibit cytotoxicity to cancer cells and safety to adjacent normal cells. Thus, they are promising agents for cancer treatment. CRAds have been widely studied in preclinical experiments as well as in clinical trials (Nemunaitis et al., 2007; Opyrchal et al., 2009). The traditional CRAds were always developed from adenovirus of serotype 5 (Ad5), which require the coxsackie virus and adenovirus receptor (CAR) localized on the cell surface to infect cancer cells (Bergelson et al., 1997). However, researchers discovered that breast cancer cells have low expression of CAR on their cell surface and consequently prevent Ad5-based CRAd from infection (Shayakhmetov et al., 2002; Auer et al., 2009). Thus, the application of conventional Ad5-based CRAds was severely limited in breast cancer therapy.
To solve the problem, adenoviruses of other serotypes that are CAR-independent to infect target cells were studied for breast cancer therapy (Ranki et al., 2007; Stoff-Khalili et al., 2007). Adenovirus of serotype 35 (Ad35), a member of group B adenovirus, was reported to infect cells in a CD46-dependent mechanism (Gaggar et al., 2003). Encouragingly, researchers showed that Ad35 fiber was able to facilitate adenovirus to infect breast cancer cells (Shayakhmetov et al., 2002; Yu et al., 2005). The knob and shaft regions of Ad35 fiber were substitutes for the same part of the Ad5-based vector to generate a 5/35 fiber-modified adenoviral vector. Its infectivity was shown to be significantly elevated for breast cancer cells. However, these adenoviral vectors lack the E1A region, which is essential for viral replication. E1A deletion disables 5/35 fiber-modified adenovirus from replication in breast cancer cells and compromises its tumor-suppressing capacity both in vitro and in vivo.
Here, we combined the advantages of oncolytic virus and fiber modification to generate a 5/35 fiber-modified CRAd, aiming to develop a new therapeutic strategy for breast cancer biotherapy. A p53-expressing cassette was also inserted into this CRAd to potentiate tumor-suppressing capacity. CD46 expression of breast cancer cells was quantified in our experiments. The infectivity of 5/35 fiber-modified adenoviral vector was also determined for breast cancer cells. Because it was still unknown whether the 5/35 fiber affects the process of adenoviral replication, the amounts of viral progeny were compared between 5/35 fiber-modified and Ad5-derived CRAds. Subsequently, expression levels of p53 mediated by these two CRAds were quantified in breast cancer cells. Finally, we investigated the tumor-suppressing capacity of 5/35 fiber-modified CRAd, both in vitro and in animal models.
Materials and Methods
Cell cultures
The breast cancer cell lines MCF-7, MDA-MB-231, BT-483, and BT-474 were purchased from the American Type Culture Collection (Manassas, VA). Bcap-37 was obtained from the Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences (Shanghai, China). The human embryonic kidney (HEK)-293 cell line was obtained from Microbix Biosystems (Toronto, Canada). MCF-7 was cultured in Eagle's Minimum Essential Medium (Life Technologies, Rockville, MD) with 0.01 mg/ml bovine insulin and fetal bovine serum to a final concentration of 10% (Life Technologies). Bcap-37 was cultured in RPMI-1640 medium (Life Technologies) with fetal bovine serum to a final concentration of 10%. MDA-MB-231 was cultured in Leibovitz's L-15 medium (Life Technologies) with fetal bovine serum to a final concentration of 10%. BT-483 was cultured in RPMI-1640 medium with 0.01 mg/ml bovine insulin and fetal bovine serum to a final concentration of 20%. BT-474 was cultured in Hybri-Care medium (Life Technologies) with fetal bovine serum to a final concentration of 10%. HEK-293 was cultured in Dulbecco's Minimum Essential Medium (Life Technologies) with fetal bovine serum to a final concentration of 10%. All media were supplemented with 4 mM glutamine, 100 units/ml penicillin, and 100 μg/ml streptomycin. All cells in this experiment were cultured under a 5% CO2 and humidified atmosphere at 37°C.
Virus production
The Ad5-based CRAd, SG600-p53, and E1A-null adenovirus Ad5-enhanced green fluorescent protein (eGFP) were prepared in our laboratory as described previously (Wang et al., 2008). To generate 5/35 fiber-modified CRAd, SG635-p53, the Ad5/35 chimeric fiber gene, was amplified from the plasmid pAd5F35/cre by polymerase chain reaction (PCR). The fragment obtained was digested by the restriction endonucleases PacI and HindIII and then inserted into pClon9-CZ, which contains both homologous arms, to generate pClon9-F35. Thus, we obtained pClon9-F35, which encodes the Ad5/35 chimeric fiber gene. Subsequently, pClon9-F35 was transfected together with pPE3, which contains the majority of the adenoviral genome sequence and is used for Ad5-based vector production in our laboratory (Zhang et al., 2004), into a Escherichia coli strain, BJ-5183, for homologous recombination, generating pPE35, an adenovirus-producing plasmid that bears the Ad5/35 chimeric fiber gene. pSG600-p53, which contains the human p53 gene expression cassette, had been already generated in our laboratory (Wang et al., 2008). pSG600-p53 was co-transfected together with pPE35 into HEK-293 cells using the Effectene transfection reagent (Qiagen, Valencia, CA). After homologous recombination in HEK-293 cells, we obtained the 5/35 fiber-modified CRAd, SG635-p53. Figure 1A shows where SG600-p53 and SG635-p53 are the same in structure, whereas Fig. 1B shows where they differ in structure.

Illustration of structures of adenoviruses used in our experiments. (
Ad5/35-eGFP, an E1A-null adenovirus that contains enhanced an eGFP-expressing cassette under the control of the cytomegalovirus early promoter, was generated through homologous recombination of pDC315-eGFP and pPE35 in HEK-293 cells.
The adenoviruses were harvested and purified with the CsCl gradient centrifugation method. The titers of all involved adenoviruses were quantified using the tissue culture 50% infectious dose (TCID50) method on HEK-293 cells and recorded as plaque-forming units (pfu)/ml.
Quantitative real-time PCR
To quantify the relative mRNA expression level of the tested cells, 5 × 104 cells per well were cultured in 24-well plates. At the time points indicated in the figure legends, cells were harvested for extraction of total RNA. The isolated RNA was reversely transcribed into cDNAs using Rever Tra Ace qPCR RT kit (Toyobo, Tokyo, Japan) according to the manufacturer's instructions. The cDNAs obtained were used for quantitative real-time (qRT)-PCR detection with the StopOne Real-Time PCR system (Applied Biosystems, Foster City, CA). The primers used for qRT-PCR are listed in Table 1. Data were analyzed with StepOne Software version 2.0 (Applied Biosystems).
GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Western blotting
The total proteins isolated were separated in 10% polyacrylamide gels and transferred onto nitrocellulose membranes (pore size, 0.45 μm), followed by blocking with 5% fat-free dry milk in Tween 20-containing Tris-HCl-buffered saline for 2 hr. The film was incubated with the primary antibodies, mouse monoclonal IgG against adenovirus E1A protein or mouse monoclonal IgG against human p53 (Santa Cruz Biotechnology, Santa Cruz, CA) (diluted 1:200), for 2 hr followed by three rinses with Tween 20-containing Tris-HCl-buffered saline and subsequently incubated with horseradish peroxidase–conjugated goat polyclonal antibody against mouse IgG (Immunology Consultants Laboratory, Inc., Newberg, OR) (diluted 1:4,000). After exposure, the membrane was processed with Restore Western Blot stripping buffer (Thermo Scientific, Rockford, IL) to eliminate the attached primary and secondary antibodies. The same membrane was then used for detecting the expression of β-actin using mouse monoclonal IgG against human β-actin (Santa Cruz Biotechnology) (diluted 1:200).
Immunofluorescence
Cells were cultured in 24-well plates at 5 × 104 cells per well. Twenty-four hours later, media were removed from the cultured cells followed by three washes with phosphate-buffered saline (PBS). Cells were fixed with 4% polyoxymethylene solution for 20 min and washed with PBS three times. For CD46 detection, cells were incubated with mouse monoclonal IgG2B against human CD46 (R&D Systems, Minneapolis, MN) (diluted 1:400) for 1 hr, followed by three rinses with PBS, and then incubated with phycoerythrin-conjugated donkey polyclonal antibody against mouse IgG (Santa Cruz Biotechnology) (diluted 1:100) for 1 hr. After three washes with PBS, cells were observed under an Eclipse TE300 fluorescent microscope (Nikon, Tokyo, Japan).
Fluorescent-activated cell sorting analysis
Breast cancer cells were grown in six-well plates at a concentration of 3.5 × 105 cells per well. Twenty-four hours later, the media were replaced with serum-free media, and then Ad5-eGFP or Ad5/35-eGFP was added to the media at MOIs of 1 and 10. Two hours later, the adenovirus-containing media were discarded, and American Type Culture Culture–recommended media were used again. At 48 hr after infection, breast cancer cells infected by Ad5-eGFP or Ad5/35-eGFP were harvested and then suspended with PBS containing 0.5% bovine serum albumin at the concentration of 1 × 106 cells/ml. The samples were assayed for green fluorescent protein expression profile using FACS (BD Biosciences, San Jose, CA).
Virus replication assay
Logarithmically growing breast cancer cells (105 cells per well) were cultured in six-well plates. Twenty-four hours later, cells were infected with SG600-p53 or SG635-p53 at an MOI of 5. Fresh media were added 2 hr later. Then, cells were washed twice with PBS and incubated at 37°C for 48 hr. Cells were lysed through three cycles of freeze and thaw. The titers of viral progenies were quantified on HEK-293 cells with the TCID50 method. The replication multiples of adenovirus in the tested cancer cells were equal to viral progeny titers at 48 hr normalized with that at the beginning of infection.
Cell viability assay
Breast cancer cells were cultured in 96-well plates at 1 × 104 cells per well. Twenty-four hours later, CRAd at the indicated MOIs was administered to cells. Seven days later, 50 μl of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) (1 μg/ml) was added to cell media. Four hours later, MTT was discarded, and 150 μl of dimethyl sulfoxide was loaded in every well. The spectrophotometric absorbance of the samples was measured with a model 550 microplate reader (Bio-Rad Laboratories, Hercules, CA) at 570 nm with a reference wavelength of 655 nm. The percentage of cell survival was calculated using the following formula: cell survival = absorbance value of infected cells/absorbance value of uninfected control cells. Eight replicate wells were measured at each MOI, and every experiment was done at least three times.
Animal experiments
All procedures for animal experiments were approved by the Committee on the Use and Care on Animals (The Second Military Medical University, Shanghai) and performed in accordance with the institution's guidelines. Bcap-37 tumor xenografts were established by subcutaneously inoculating 5 × 106 cells into the right flanks of 4–6-week-old BALB/c nude mice (Institute of Animal Center, Chinese Academy of Sciences, Shanghai). When tumors reached between 6 and 8 mm in diameter, 21 mice were randomly assigned to groups: PBS-treated (n = 6), SG600-p53-treated (n = 7), and SG635-p53-treated (n = 8), respectively. The established tumors were injected with 100 μl of PBS with or without 2 × 108 pfu of SG600-p53 or SG635-p53. The injections were repeated five times every other day, with a total dosage of 1 × 109 pfu of adenoviruses. Tumor growth was monitored by periodic measurements with calipers, and tumor volume was calculated using the following formula: tumor volume (mm3) = maximal length (mm) × (perpendicular width) (mm)2/2. Animals were sacrificed when the diameter of tumors reached 2 cm. In our experiments, no mice were observed to die from tumor loading.
Immunohistochemistry
Deparaffinized tumor sections were treated with goat polyclonal antibody against Ad5 hexon protein (Santa Cruz Biotechnology) (diluted 1:100) or mouse monoclonal antibody against human p53 (Santa Cruz Biotechnology) (diluted 1:100). After incubation with rabbit secondary antibody (Immunology Consultants Laboratory) (diluted 1:100), hexon or p53 expression was detected with 3,3'-diaminobenzidine (Sigma, St. Louis, MO). The sections were then counterstained with hematoxylin.
Statistical analysis
Absorbance value in the MTT assay and mean tumor volume are presented as mean ± SD values and compared at a given time point by unpaired, two-tailed t test. Results were compared for statistical significance by applying the generalized Wilcoxon test. Difference betwee data were considered to be statistically significant when p < 0.05 (*) and to be very significant when p < 0.01 (**).
Results
CAR and CD46 expression in breast cancer cells
CAR is the primary cellular receptor for Ad5 to recognize target cells (Bergelson et al., 1997), whereas CD46 can mediate Ad35 to infect target cells (Gaggar et al., 2003). Therefore, we quantified expression level of these two cell surface proteins in breast cancer cells. CAR expression is positive in the human hepatocellular carcinoma cell line Hep3B, and Ad5 can efficiently infect Hep3B (Li et al., 2008). Thus, we used Hep3B as a positive control for CAR quantification. qRT-PCR data showed that CAR mRNA expression is dramatically lower in MCF-7, Bcap-37, MDA-MB-231, BT-483, and BT-474 human breast cancer cells than that in Hep3B (Fig. 2A). Meanwhile, all breast cancer cells tested in our experiment had higher CD46 expression than Hep3B (Fig. 2B). The immunofluorescent data consistently showed that MCF-7, Bcap-37, and MDA-MB-231 expressed CD46 protein on their cell membrane (Fig. 2C).

Expression profiles of CAR and CD46 in breast cancer cells. Expression levels of CAR and CD46 mRNA in hepatocellular carcinoma cells, Hep3B, and the breast cancer cells MCF-7, Bcap-37, MDA-MB-231, BT-483, and BT-474 were measured by qRT-PCR and the immunofluorescent method. The relative expression level of (
Low CAR expression and high CD46 expression of breast cancer cells make Ad35 a suitable gene therapy vector instead of Ad5 for breast cancer treatment. Next, we selected 5/35 fiber-modified adenoviral vector to transfer the therapeutic gene into breast cancer cells.
Infectivity of 5/35 fiber-modified adenovirus to breast cancer cells
The low Ad5 infectivity for breast cancer cells was reported previously (Shayakhmetov et al., 2002). High-level expression of CD46 in breast cancer cells makes Ad35 as a suitable gene vector for breast cancer treatment. Next, we selected the 5/35 fiber-modified adenoviral vector to transfer the therapeutic gene into breast cancer cells. To investigate the infectivity of 5/35 fiber-modified adenoviral vector to breast cancer cells, Ad5-eGFP and Ad5/35-eGFP at an MOI of 1 or 10 were added to the media, followed by fluorescence detection and FACS analysis. Higher percentages of the eGFP-positive cell population were observed in all 5/35 fiber-modified vector-infected breast cancer cells than that in Ad5-infected groups, whether an MOI of 1 (Fig. 3A) or an MOI of 10 (Fig. 3B) of adenoviral vectors was used. The results showed that 5/35 fiber-modified adenoviral vector had a higher infectivity for breast cancer cells than Ad5-based vector.
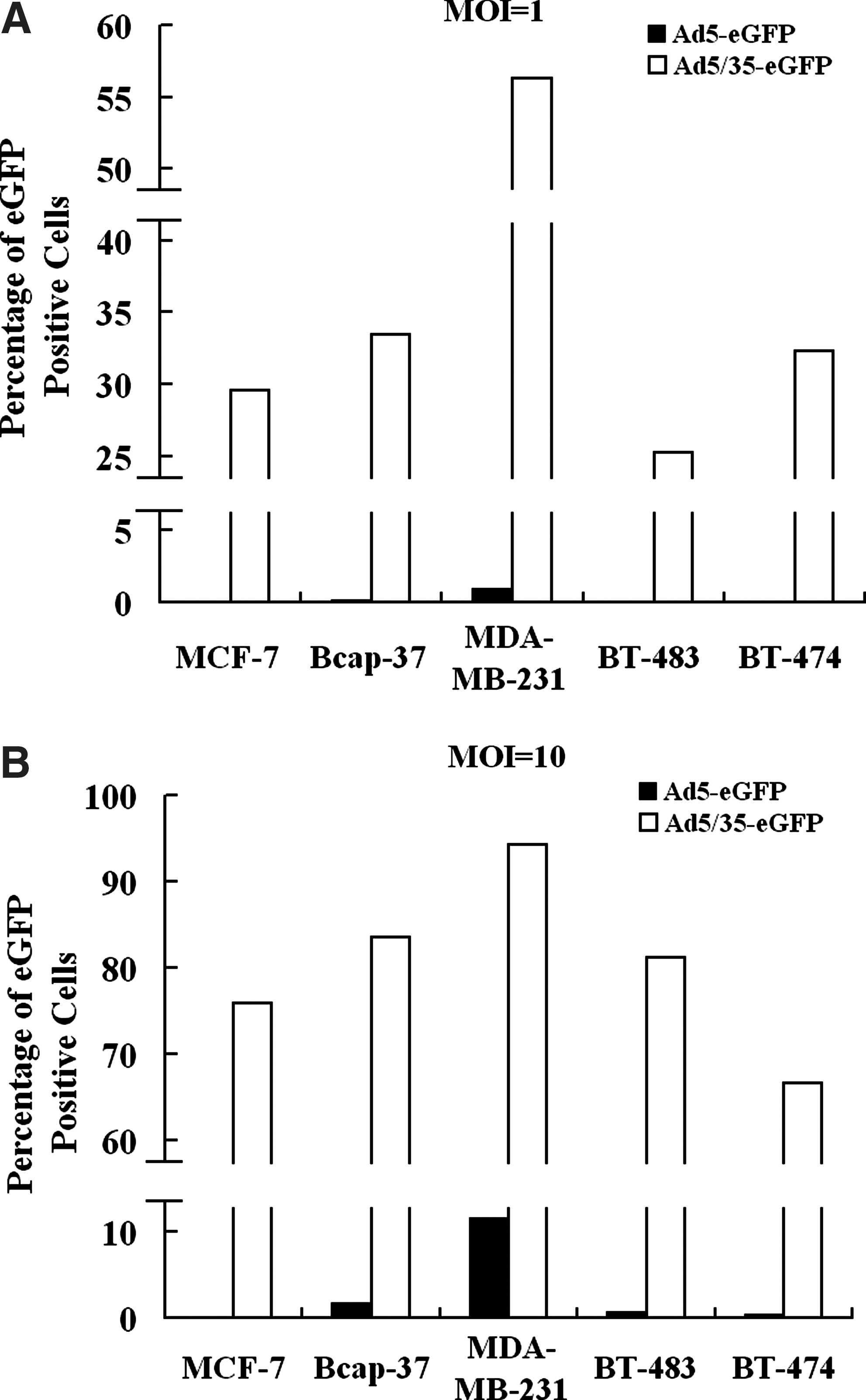
Infectivity of Ad5-eGFP and Ad5/35-eGFP for breast cancer cells. Breast cancer cells—MCF-7, Bcap-37, MDA-MB-231, BT-483, and BT-474 (3 × 104 cells per group)—were infected by Ad5-eGFP and Ad5/35-eGFP at MOIs of (
Replication of 5/35 fiber-modified CRAd in breast cancer cells
Efficient viral replication and high progeny production can greatly contribute to the antitumor capacity of CRAds. However, the replication of this fiber-modified CRAd in breast cancer cells remained poorly understood. Here, we studied the replicative ability of 5/35 fiber-modified CRAd in breast cancer cells. The activity of E1A protein regulates the adenoviral replication in the infected cells. First, we examined the effect of E1A expression on both mRNA and protein levels in SG635-p53- or SG600-p53-infected breast cancer cells. qRT-PCR and western blotting data showed higher expression of E1A in all SG635-p53-infected breast cancer cells (Fig. 4A and B).
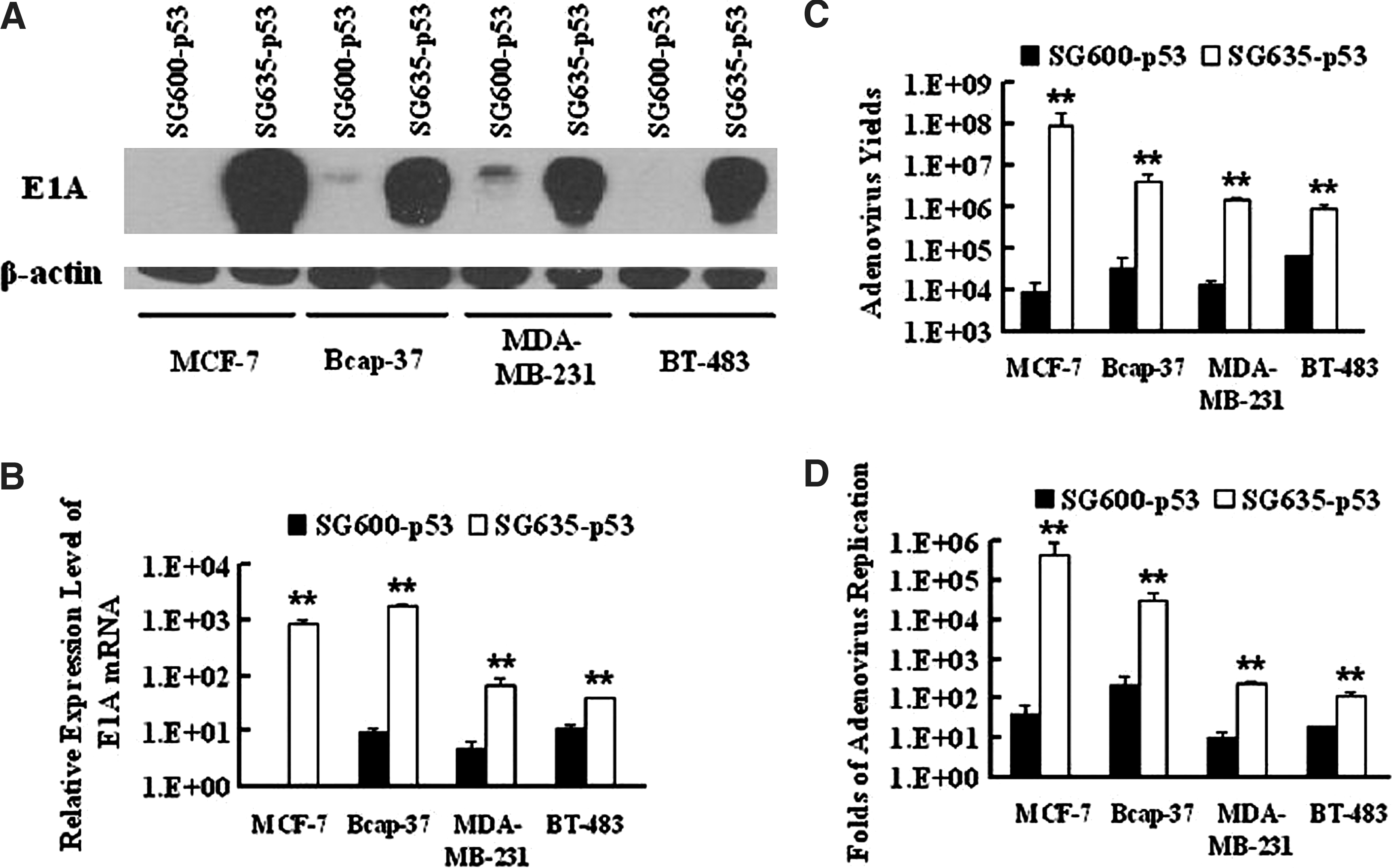
Replication of SG600-p53 and SG635-p53 in breast cancer cells. SG600-p53 and SG635-p53 at an MOI of 5 were used to infect breast cancer cells to evaluate their ability to produce viral progeny within cancer cells. (
Production of infective viral progeny was also quantified in SG600-p53- or SG635-p53-infected breast cancer cells by the TCID50 method. The results demonstrated that SG635-p53-infected breast cancer cells produced higher titers of viral progenies than the SG600-p53-infected groups (Fig. 4C). Interestingly, 5/35 fiber-modified CRAd was shown to have higher replication efficiency than Ad5-based CRAd (Fig. 4D). Above all, CRAd with 5/35 chimeric fiber replicated more efficiently and produced greater amounts of progeny viruses than Ad5-based CRAd in breast cancer cells.
CRAd-mediated p53 expression in breast cancer cells
In addition to directly lysing cancer cells, CRAd can also be used as a vehicle for gene transfer (Zhang et al., 2004; Liu, 2006). Unlike replication-incompetent adenoviral vector, CRAd can amplify the copy number of the transferred cytotoxic gene within infected cancer cells and increase their tumor-suppressing activity (Zhang et al., 2004). Next, we quantified the effect of p53 expression on levels of mRNA and protein. qRT-PCR data demonstrated that the relative expression level of p53 mRNA was dramatically elevated in SG635-p53-infected breast cancer cells (Fig. 5A). Consistently, western blotting results also showed that 5/35 fiber-modified CRAd transferred more p53 into breast cancer cells than Ad5-derived vector (Fig. 5B). In conclusion, we found that 5/35 fiber-modified CRAd could transfer a greater amount of p53 into breast cancer cells than Ad5-based CRAd.
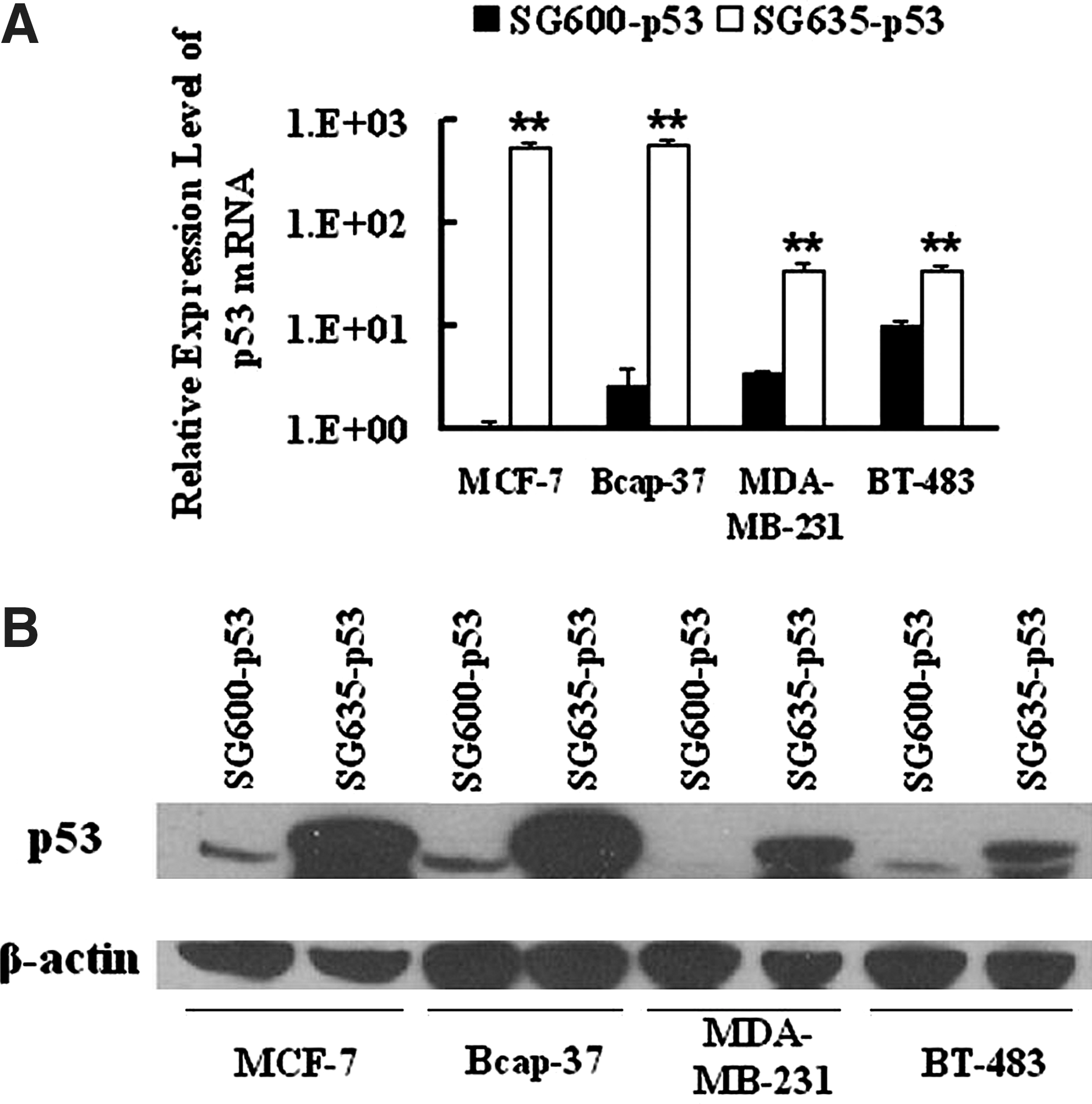
p53 expression mediated by SG600-p53 and SG635-p53 in breast cancer cells. Expression levels of p53 gene transferred into breast cancer cells were examined after infection with SG600-p53 or SG635-p53. (
Tumor-killing capacity of 5/35 fiber-modified CRAd in vitro
The tumor-suppressing ability of the 5/35 fiber-modified and Ad5-based CRAds was investigated in vitro by MTT assay. The data showed that SG635-p53 had a higher antitumor capacity for breast cancer cells than SG600-p53 (Fig. 6). For example, MDA-MB-231 was shown to have a very high sensitivity to SG635-p53 and even was completely eliminated even at a low MOI of CRAd (Fig. 6). The 50% inhibitory concentration between SG600-p53 and SG635-p53 was also calculated for each cell line and is shown in Table 2. Our data demonstrated that 5/35 fiber-modified CRAd can eliminate breast cancer cells more effectively than Ad5-based CRAd in vitro.
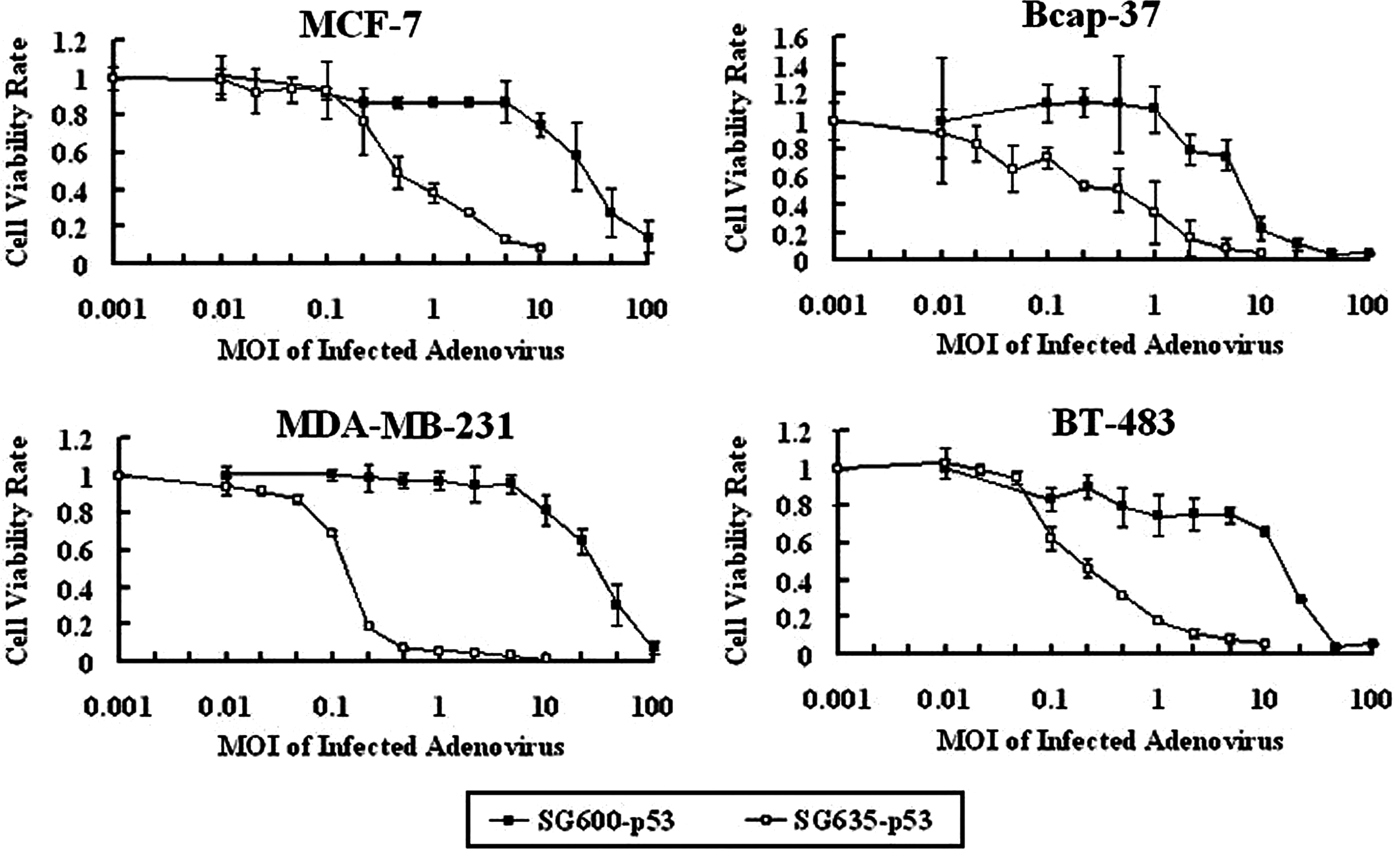
Antitumor capacity of SG600-p53 and SG635-p53 for breast cancer cells. The tumor-suppressing abilities of SG600-p53 and SG635-p53 for breast cancer cells were investigated by the MTT assay. Breast cancer cells (1 × 104 per well) were cultured in 96-well plates. Following overnight culture, SG600-p53 or SG635-p53 at the indicated MOIs was added to media. Seven days later, the viability of infected cells was detected using the MTT assay.
IC50, 50% inhibitory concentration.
Growth-inhibiting ability of 5/35 fiber-modified CRAd for breast cancer in an animal model
We next investigated the tumor-suppressing capacity of SG635-p53 for breast cancer cells in nude mice. Bcap-37 was used to establish tumors in BALB/c nude mice (Zou et al., 2004). When tumor diameters reached between 6 and 8 mm, SG635-p53 and SG600-p53 were intratumorally injected for five times every other day giving a total dose of 1 × 109 pfu. The data of tumor volumes suggested that SG635-p53 was able to efficiently inhibit the growth of Bcap-37 cancer xenograft (Fig. 7A). It was noteworthy that tumors completely disappeared in two of eight mice in the SG635-p53-injected group.

Ability of SG600-p53 and SG635-p53 to inhibit the growth of breast cancer xenografts in nude mice. Bcap-37 breast cancer cells were inoculated subcutaneously into nude mice at 5 × 106 cells per mouse to establish tumor xenografts. When tumor diameters reached 6–8 mm, mice were injected with PBS, SG600-p53, or SG600-p53 every other day for a total of five injections. The tumor volumes were measured at the indicated time points. When tumors grew up to 2 cm in diameter, the mice were sacrificed, and the tumors were dissected. (
Hematoxylin and eosin staining showed that SG635-p53 can lead to a larger area of necrosis in tumors than SG600-p53, indicating that 5/35 fiber-modified CRAd had more destructive effect on tumor tissue than Ad5-based CRAd (Fig. 7B). Immunohistochemical staining of adenoviral hexon protein demonstrated that more viral progenies were produced in the tumor section from SG635-p53 injected mice (Fig. 7B) and that SG635-p53-treated tumor cells had higher p53 expression. Above all, we observed an increased tumor-suppressing efficacy of 5/35 fiber-modified CRAd for breast cancer than traditional Ad5-based CRAd in vivo.
Discussion
Because of the lack of CAR expression in some types of cancer cells and the existence of neutralizing antibody to Ad5 in patients, the improvement of adenoviral vectors in tropism is greatly needed. Many modifications in the fiber region have been made, such as 5/3 chimeric fiber (Rein et al., 2005) and 5/11p chimeric fiber (Lu et al., 2006). Although the substitution of Ad5 fiber with replication-incompetent adenoviral vector with 5/35 chimeric fiber is quite effective in cancer treatment (Brouwer et al., 2007; Ganesh et al., 2007; Hoffmann et al., 2007a,b), the antitumor efficacy of CRAd with 5/35 chimeric fiber for breast cancer had not been studied yet. Here, we reported that 5/35 fiber-modified CRAd has an elevated tumor-suppressing capacity for breast cancer cells compared with traditional Ad5-based vector.
Previous studies showed that breast cancer cells had a low expression level of CAR protein (Shayakhmetov et al., 2002; Stoff-Khalili et al., 2005). In our experiments, all breast cancer cells tested, including MCF-7, Bcap-37, MDA-MB-231, BT-483, and BT-474, were demonstrated to have low CAR expression, on the level of both mRNA and protein. Lack of this primary cellular receptor for adenovirus may be responsible for inefficient infection of Ad5-based CRAd into breast cancer cells.
Ad35 infects target cells in a CD46-dependent manner (Gaggar et al., 2003). Down-regulation of cellular CD46 was reported to reduce infectivity of 5/35 fiber-modified adenoviral vector (Yu et al., 2007), indicating that the variation in CD46 expression of breast cancer cells may affect the antitumor capacity of 5/35 fiber-modified CRAd. Our data clearly showed high CD46 expression in all breast cancer cells tested, which was consistent with a previous report (Stoff-Khalili et al., 2005). These results may account for the efficient infection of 5/35 fiber-modified CRAd for breast cancer cells in our experiments.
In addition to detecting the expression of cellular primary receptor for Ad5/35, we directly investigated the infectivity of 5/35 fiber-modified adenoviral vector for breast cancer cells. A significantly higher percentage of eGFP-positive cells was observed in Ad5/35-eGFP-infected breast cancer cells, demonstrating that 5/35 fiber-modified adenoviral vector has a higher infectivity for breast cancer cells. Efficient infection of 5/35 fiber-modified adenoviral vector was also reported for breast cancer cells by other investigators (Yu et al., 2007). Increased infectivity of 5/35 fiber-modified adenovirus largely contributes to the enhanced antitumor capacity of Ad5/35-based CRAd.
Interestingly, our data strongly showed that replication of 5/35 fiber-modified CRAd is more efficient than that of Ad5-based CRAd in breast cancer cells. Accordingly, 5/3 fiber-modified CRAd was also reported to have higher replicative capacity than Ad5-based vector (Kawakami et al., 2003; Stoff-Khalili et al., 2007). This phenomenon enhanced the antitumor capacity of 5/35 fiber-modified CRAd for breast cancer cells, although the underlying mechanism is still unknown.
Our data clearly demonstrated that p53 gene transfer was dramatically elevated when 5/35 fiber-modified vector was used as a gene therapy vector. Besides the p53 gene, 5/35 fiber-modified CRAd can also be used to transfer other therapeutic genes, including those for anti-angiogenic protein (Zhang et al., 2004; Su et al., 2008), cytotoxic protein (Su et al., 2006), tumor-suppressing microRNA (Ma et al., 2010), and pro-drug activating enzyme or full-length antibody (Jiang et al., 2006; Chen et al., 2008), into breast cancer cells to enhance antitumor capacity. This strategy is also known as gene-virotherapy (Zhang et al., 2004), in which replication of adenoviruses within tumor cells can enhance the tumor-suppressing capacity of therapeutic genes.
Breast cancer cells displayed a very high sensitivity to CRAds carrying 5/35 hybrid fiber. A 5/35 fiber-modified CRAd of low titer can almost eliminate breast cancer cells, such as MDA-MB-231. Animal experiments also confirmed that 5/35 fiber-modified CRAd had an increased antitumor capacity for breast cancer cells in nude mice. All data showed that 5/35 fiber-modified CRAds have a higher antitumor capacity for tumor cells than traditional Ad5-based CRAds, which is consistent with studies on some other types of cancer and leukemia (Jin et al., 2009; Wang et al., 2009).
In spite of improved antitumor efficacy of 5/35 fiber-modified CRAd in our experiments, it was also reported that CD46-dependent adenoviruses inhibit peripheral blood mononuclear cell expression of some pro-inflammatory cytokines and compromise the function of the human immune system (Iacobelli-Martinez et al., 2005). The use of 5/35 fiber-modified adenoviral vector may compromise the tumor-suppressing function of immune cells.
Above all, we successfully engineered CRAd carrying 5/35 chimeric fiber. This fiber-modified CRAd armed with the p53 gene was shown to inhibit the growth of breast cancer cells more effectively both in vitro and in vivo. We provided a novel promising vector to transfer a therapeutic gene into breast cancer. We hope that 5/35 fiber-modified CRAd can contribute to more effective breast cancer treatment.
Footnotes
Acknowledgments
This work is supported by the National Science Fund for Distinguished Young Scholars of the National Natural Science Foundation of China (NSFC) (number 30925037), the Key Program of the NSFC (number 30730104), the Science Fund for Creative Research Groups of the NSFC (number 30921006), the Key Project of the Zhejiang Natural Science Foundation (number Z205618), and the National Significant Science and Technology Special Projects of New Drugs Creation (number 2009ZX09102-235).
Author Disclosure Statement
No competing financial interests exist.