
Abstract
The aim of this study was to investigate the effect of microRNA-20a on pancreatic carcinoma cell proliferation and invasion and to find a new effective treatment strategy for pancreatic carcinoma. MicroRNA-20a expression was determined in 10 matched normal pancreatic tissues and pancreatic carcinoma by in situ hybridization. Quantitative real-time RT-PCR was used to evaluate the expression of microRNA-20a in two pancreatic carcinoma cell lines (BxPC-3 and Panc-1) and immortal human pancreatic duct epithelial cell line H6C7. Proliferation and invasion capacity were analyzed for the cells with lentivirus-mediated overexpression of microRNA-20a both in vitro and in vivo. In addition, the regulation of signal transducer and activator of transcription proteins 3 (Stat3) by microRNA-20a was determined to elucidate the underlying mechanisms. The pancreatic cancer cell lines (Panc-1 and BxPC-3) stably overexpressing microRNA-20a showed reduced proliferation and invasion capacity in vitro and in vivo, compared with parental cells or cells transfected with a control vector. Furthermore, we found that microRNA-20a negatively regulated Stat3 protein expression in a dose-dependent manner without changing the Stat3 mRNA level and decreased the activity of a luciferase reporter construct containing the Stat3 3′-untranslated region. These results show that microRNA-20a regulates Stat3 at the post-transcriptional level, resulting in inhibition of cell proliferation and invasion of pancreatic carcinoma. It may open a new perspective for the development of effective gene therapy for pancreatic carcinoma.
Introduction
MicroRNAs (miRNAs; ∼22 nucleotides [nt]) are small noncoding RNAs regulating gene expression by hybridizing to the sequences usually located in the 3′-untranslated region (UTR) of coding transcripts, and play critical roles in cancer cell proliferation (Bandres et al., 2009; Hiyoshi et al., 2009), apoptosis (Matsubara et al., 2007), and metastasis (Ma et al., 2007). Some microRNAs have been classified as tumor suppressors or oncogenes. Functional studies also have linked aberrant microRNA expression to carcinogenesis. Recently, several groups have reported the microRNA profile of pancreatic cancer (Schmittgen et al., 2004; Volinia et al., 2006; Bloomston et al., 2007; Zhang et al., 2009). However, in these studies, microRNAs were isolated from the whole tumors but not from microdissected pancreatic ductal adenocarcinoma (PDAC) cells, and thus the results of miR-17–92 cluster expression in pancreatic cancer were distinct. The entire miR-17–92 cluster is located within the third intron of an open reading frame termed C13orf25 and correlates with loss of heterozygosity in several different cancers, including breast cancer and lung cancer (Eiriksdottir et al., 1998; Lin et al., 1999). MicroRNA-20a is a member of the miR-17–92 family, which shares extensive sequence similarity and significant overlapped targets (Yu et al., 2008). The 3′-UTR of signal transducer and activator of transcription proteins 3 (Stat3) has been known to contain the microRNA-20a binding site, and Stat3/microRNA-20a interaction could regulate ES cell differentiation (Foshay and Gallicano, 2009). Stat3 is activated in primary pancreatic cancer and is involved in various physiologic functions (Greten et al., 2002; DeArmond et al., 2003; Aggarwal et al., 2006; Sahu and Srivastava, 2009), including apoptosis, cell cycle regulation, angiogenesis, and metastasis. Therefore, we hypothesized that microRNA-20a should be a genuine regulator of oncogene Stat3 in pancreatic carcinoma.
In the present study, we found that expression of microRNA-20a was reduced in pancreatic cancer tissues and cell lines. MicroRNA-20a regulated Stat3 protein expression negatively at the post-transcriptional level and inhibits cell proliferation and invasion of pancreatic carcinoma in vivo and in vitro. Our data provided strong evidence for the potential use of microRNA-20a as a novel approach to treat human pancreatic cancer.
Materials and Methods
Tissue samples and cell lines
Ten matched normal pancreatic tissues and pancreatic carcinoma used in this study were obtained from patients in Wuxi People's Hospital of Nanjing Medical University from 2008 to 2009. The samples were obtained with patients' informed consent and were histologically confirmed. All tissue samples were derived from untreated patients undergoing surgery, fixed by formalin immediately, and embedded in paraffin. The human immortalized pancreatic ductal epithelial cell line H6C7 was kindly provided by Prof. Ming-sound Tsao (Ontario Cancer Institute, Toronto University, Toronto, ON, Canada) and cultured in keratinocyte serum-free medium (Invitrogen, Carlsbad, CA). The pancreatic ductal adenocarcinoma cell lines Panc-1 and BxPC-3 were abtained from the American Type Culture Collection (Manassas, VA) and maintained in DMEM supplemented with 10% fetal bovine serum (FBS; GIBCO), 100 units/ml penicillin, and 100 μg/ml streptomycin. All the cells were cultured in a humidified 37°C incubator with 5% CO2. Cells were tested regularly for mycoplasma (the new MycoProbe Mycoplasma Detection Kit from R&D Systems, Minneapolis, MN).
Total RNA extraction and real-time RT-PCR
Total RNA was extracted from the cultured cells using TRIzol (Invitrogen), RNA was submitted to DNase digestion, and the concentration of total RNA was determined by measuring the absorbance at 260 nm. Synthesis of cDNA and real-time RT-PCR were performed using the TaqMan MicroRNA Reverse Transcription Kit and the microRNA-20a TaqMan MicroRNA Assay Kit, respectively (Applied Biosystems, Carlsbad, CA). Real-time PCR was performed on an Applied Biosystems 7900HT real-time PCR instrument equipped with a 96-well reaction block. The expression level of microRNA-20a was normalized to RNU6B and determined using the 2-ΔΔC(T) method (Livak and Schmittgen, 2001).
Detection of microRNA-20a by in situ hybridization
Probes for microRNA-20a and scramble labeled at the 5′-end with fluorescein isothiocyanate (Exiqon, Vedbaek, Denmark). Thirty percent of the oligonucleotides within the sequence is modified to an LNA. Probe was designed to hybridize to the entire mature miRNA sequence. In situ hybridization was performed according to the GEISHA protocol (from the University of Arizona) available on the Exiqon Web site and modified as per previously published protocol (Silahtaroglu et al., 2007; Nuovo, 2008). All reagents and apparatus were treated with diethylpyrocarbonate water. Hybridizations were conducted overnight at 55°C. The result was analyzed by ImageJ 1.37v (National Institutes of Health, Bethesda, MD).
Immunocytochemistry
Tumor cells were harvested by trypsinization and seeded onto poly-L-lysine–treated glass slides for 24 hr. The cells were then fixed for 5 min in methanol at −20°C, and the immunocytochemistry was performed as reported previously (Qiu et al., 2007).
Lentivirus-mediated overexpression of microRNA-20a
MicroRNA-20a precursor and flanking sequence (approximately 100 bp) was amplified from human genomic DNA and cloned into the BamHI and MfeI site of the lentiviral vector pEZX-MR03 (GeneCopoeia, Rockville, MD). Primer sequences for microRNA-20a are: Forward: 5′-GCCCTGCAATATTTGCATGTCGCT-3′; and Reverse: 5′-GTACATTATAAGCTGCAA-3′. Primer sequences for control are: Forward: 5′-CCGACAACCACTACCTGA-3′; and Reverse: 5′-CGTGAAGAATGTGCGAGAC-3′. Lentiviruses were generated by cotransfecting 2.5 μg of lentiviral vector and 5 μl of each packaging vector. Following transfection of 293FT packaging cells, lentiviral supernatants were collected, filtered, and added to Panc-1 and BxPC-3 for 8 hr in the presence of 8 μg/ml polybrene. Two days after infection, 1 μg/ml puromycin was added to the media to select the cell populations infected with the lentivirus for 1 week. The two cell lines expressing microRNA-20a stably were named LV-miR-20a Panc-1 and LV-miR-20a BxPC-3, respectively; the control vector cell lines were named LV-miR-control Panc-1 and LV-miR-control BxPC-3. Real-time PCR assay was used to detect the expression of microRNA-20a in the cell lines as described above and in parent cells.
MicroRNA-20a RNA oligonucleotides and transfection
The microRNA-20a mimics (UAAAGUGCUUAUAGUGCAGGUAG) and inhibitors (CUACCUGCACUAUAAGCACUUUA) were purchased from Shanghai GenePharma Co. (Shanghai, P.R. China). siRNA duplexes with nonspecific sequences were used as a negative control (AGACUAGCGGUAUCUUUAUCCC). For transient transfection, Panc-1 and BxPC-3 cells were seeded in a 24-well plate at a concentration of 1 × 105 cells/well and allowed to adhere for 24 hr. Then Lipofectamine 2000 (Invitrogen) was used for transfecting cells according to the manufacturer's instructions. Cells were cultured for 6 hr in a humidified 37°C incubator with 5% CO2 atmosphere, and then the medium was replaced with fresh medium supplemented with 10% FBS. After transfection, cells were cultured for 48 hr and were analyzed by Western blot or used for proliferation and invasion.
Cell proliferation and assessment of clonogenicity
For the cell proliferation assay, the tumor cells of each group were harvested and seeded at a density of 50 cells/mm2 in triplicate. The total cell number was counted every 2 days with a hematocytometer and an Olympus inverted microscope. The cell viability was assessed by trypan blue staining.
For the clone formation assay, the tumor cells were seeded at low density (5 × 103 cells per 10-cm2 plate) and incubated for 10 days. Colony formation and growth were visualized with crystal violet staining. After the wells were photographed, the dye was solubilized with methanol and the optical density at 590 nm was measured by an ELISA reader. The surviving fraction was calculated relative to the control cells.
Cell cycle analysis
Each cell line above was cultured in triplicate onto six-well plates; when cells were about 75% confluent, the cells were collected and washed with ice-cold PBS and fixed in 70% ethanol at 4°C overnight. Then fixed cells were washed and resuspended in 100 μl of PBS, stained with 50 μg/ml propidium iodide for 30 min, and finally analyzed by Flow Cytometry (Beckman Coulter, Brea, CA).
Migration and invasion assay in vitro
Invasion assay was carried out using a specialized invasion chamber (BD Biosciences, Franklin Lakes, NJ). DMEM supplemented with 10% FBS was poured into the lower chamber. Tumor cells starved overnight in serum-free medium were resuspended in DMEM with 0.1% FBS (1.0 × 106 cells/ml) and seeded 0.3 ml into the upper chambers. After 8 hr of incubation at 37°C, cells on the upper surface of the membrane were scrubbed off. Cells on the lower surface of the membrane, which had migrated through the polycarbonate membrane, were fixed with ethanol and stained by crystal violet for 10 min. For the invasion assay, the cell culture insert surface was coated with 5 μg of Matrigel and incubated at 37°C for 16 hr. The other steps of invasion are the same as migration. The migration rate and invasion rate were quantified by counting the migration cells in six random fields. The experiment was repeated three times.
Western blotting
Cell lysates were prepared by extracting proteins with RIPA buffer containing a protease inhibitor cocktail (Upstate, Lake Placid, NY) according to the manufacturer's instructions. Anti-Stat3 monoclonal antibody, matrix metalloproteinase-2 (MMP2) monoclonal antibody (Cell Signaling Technology, Beverly, MA), and anti-GAPDH antibody (Santa Cruz Biotechnology, Santa Cruz, CA) were used at 1:3,000, 1:1,000, and 1:5,000 dilution, respectively. Western blotting of GAPDH on the same membrane was performed as a loading control. Proteins were ultimately visualized using the SuperSignal enhanced chemiluminescence kit (Pierce, Rockford, IL).
ELISA for vascular endothelial growth factor (VEGF) protein
VEGF was determined using an ELISA kit specific to human VEGF (BioSource Europe). Our pancreatic cancer cells lines were seeded at 70% to 80% density for 72 hr. Analyses of culture supernatants were performed according to the manufacturer's protocol.
Luciferase reporter assays
The 3′-UTR of Stat3 mRNA containing the microRNA-20a binding site was amplified by PCR (primers: 5′-GCTCTAGATGC TATCTTTGAGCAATCTGG-3′ [forward] and 5′-GCTCTAGATCCCAAAGTGCTGG GAT-3′ [reverse]), cloned into the XbaI site of pGL3 (Promega, Madison, WI), and named pStat3-W. The mutations were built and named pStat3-M1, pStat3-M2, and pStat3-M3. Panc-1 and BxPC-3 cells (2.5 × 105) were plated in triplicate wells of a 24-well plate and 16 hr later transfected with 800 ng of the indicated pGL3-Stat3-3′-UTR reporter constructs and 16 ng of pGL4.73 (Promega) by using Lipofectamine 2000 (Invitrogen). Where indicated, microRNA-20a mimics or scramble (GenePharma) were cotransfected at 50 nM final concentration; 48 hr after transfection, cells were lysed and assayed for firefly and renilla luciferase activity using the Dual-Luciferase Reporter Assay System (Promega). Firefly luciferase activity was normalized to renilla luciferase activity for each transfected well. Data depicted are representative of three independent experiments performed on different days.
Semiquantitative RT-PCR
Semiquantitative RT-PCR was done to assess Stat3 mRNA expression by using the following sense and antisense primers: Stat3-Forward (310 bp): 5′-CTTGGGTGGAGAAGGACA-3′; and Stat3-Reverse: 5′-CGGCAGGTCAATGGTATT-3′. GAPDH mRNA expression was chosen as a housekeeping gene, and a 628-bp sequence was amplified by using the following primers: 5′-GGTGAAGGTCGGAGTCAA-3′ and 5′-GCCAGTAGAGGCA GGGAT-3′. PCR was carried out in the exponential range of amplification with the following conditions: Stat3 and GAPDH: 28 cycles of 40 sec at 94°C, 30 sec at 55°C, and 40 sec at 72°C.
In vivo proliferation and metastasis assay
Specific pathogen-free female athymic BABL/c nude mice (6 weeks old) were obtained from the Animal Center of Chinese Academy of Science (Shanghai, P.R. China). The use of animals in this study complies with the Guide for the Care and Use of Laboratory Animals (NIH publication no. 86–23, revised 1985) and the current Chinese regulations and standards on the use of laboratory animals. To investigate the proliferation capacity of cell lines stably expressing microRNA-20a in vivo, we adopted the procedure for animal experiments described previously (Wei et al., 2003; Qiu et al., 2007). Tumor cell suspensions (1 × 106 cells/mouse) were injected subcutaneously into each nude mouse (six mice for each group). The tumor volume was evaluated using the following formula: tumor volume = 4π/3 × (width/2)2 × (length/2). Tumor size was represented by the three-dimensional volume of an ellipse. To evaluate experimental lung metastasis, tumor cell suspensions (5 × 105 cells/mouse) were injected into the lateral tail veins of each anesthetized nude mouse (n = 6). The metastases were counted in a double-blind manner with the aid of a dissecting microscope as described previously (Huang et al., 1998), and the metastasis nodi were confirmed by histopathological examination. To detect the expression of Stat3, MMP2, and VEGF in vivo, we carried out the immunohistochemistry analysis as described previously (Qiu et al., 2007).
Statistical analysis
The significance of the in vitro results was determined using Student's t test (two-tailed), whereas the significance of the differences between the median values of the in vivo data was determined using the two-tailed Mann–Whitney or Kruskal–Wallis test. A value of P < 0.05 was considered statistically significant.
Results
Expression level of microRNA-20a and Stat3 in pancreatic carcinoma
In situ hybridization showed that the expression level of microRNA-20a was lower in pancreatic carcinoma tissues than that in the normal pancreatic tissues. To detect the specificity of the probes, an LNA probe with a scrambled sequence was used as a control, and no signals were obtained in either the tumor or normal tissue (Fig. 1A). Real-time RT-PCR showed that expression of microRNA-20a was down-regulated in Panc-1 and BxPC-3 compared with that in human immortalized pancreatic ductal epithelial cell line H6C7 (P < 0.01; Fig. 1B). It should be noted that there was strong positive expression of Stat3 in pancreatic adenocarcinoma tissues, but weak or negative expression of this gene in normal tissues as indicated by immunohistochemistry results (Fig. 1C). Furthermore, immunocytochemistry showed that Stat3 was localized in the cytoplasm of Panc-1 and BxPC-3, but was negative in H6C7 (Fig. 1D). These data indicated there was an inverse correlation between microRNA-20a and Stat3 in pancreatic adenocarcinoma cells.
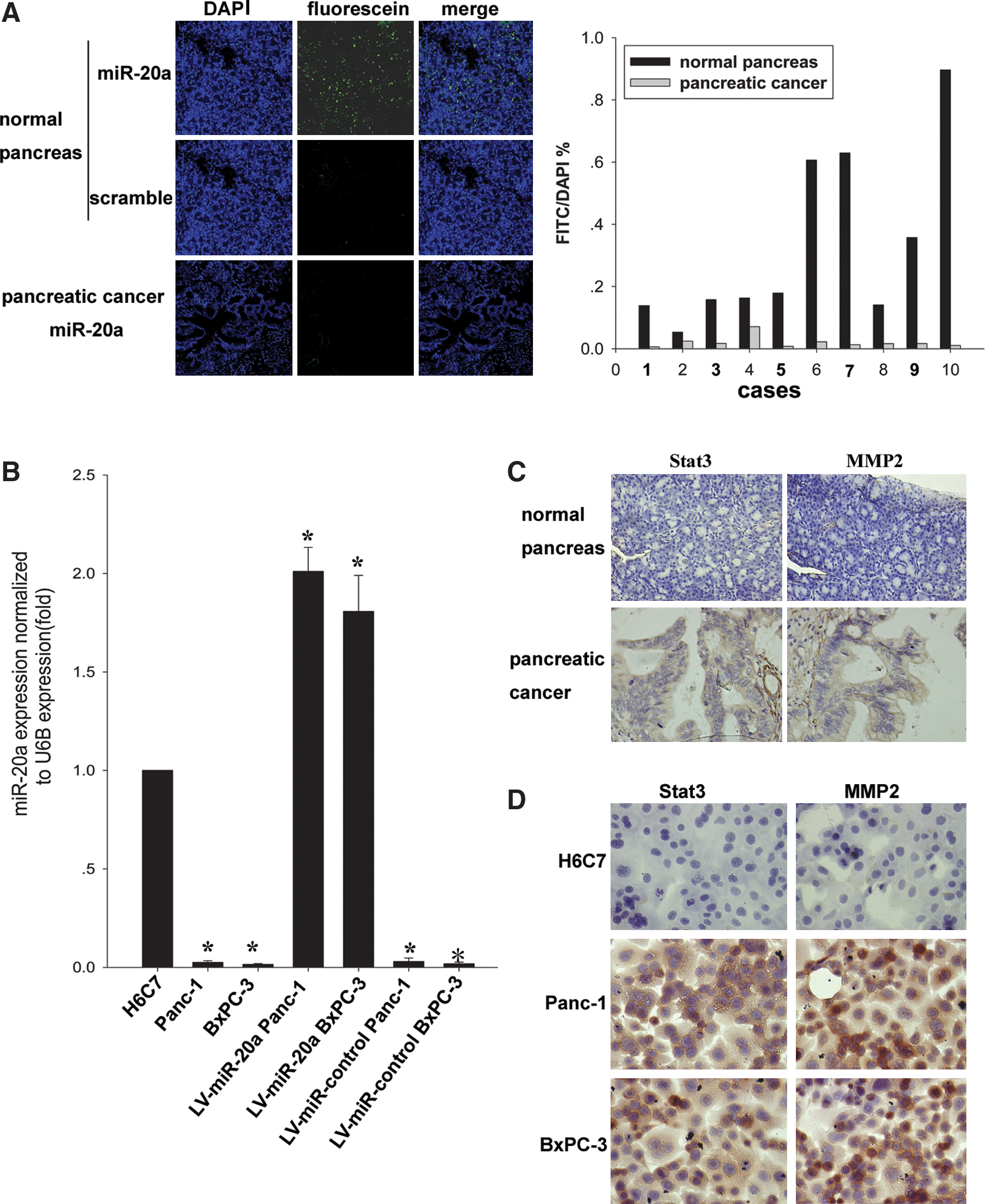
Detection of microRNA-20a, Stat3, and MMP2 in pancreatic carcinoma tissues and cell lines.
Antiproliferative effects of microRNA-20a on pancreatic cancer cells in vitro
Cell proliferation assay was performed for LV-miR-20a Panc-1 and LV-miR-20a BxPC-3 cells in which microRNA-20a was up-regulated (P < 0.01; Fig. 1B) and for the control groups. The cell growth curves showed that the growth rate of tumor cells was significantly decreased by overexpression of microRNA-20a in a time-dependent manner (P < 0.01; Fig. 2A). The maximum inhibitory rates for Panc-1 and BxPC-3 were 59.21 ± 2.31% and 59.00 ± 2.01% on day 8, respectively. There was no significant difference between the control vector cells and parental cells (P > 0.05; Fig. 2A). We also evaluated the effect of ectopic expression of microRNA-20a on cell proliferation through clonogenic assay. Compared with the control groups after 10 days, proliferation of LV-miR-20a Panc-1 and LV-miR-20a BxPC-3 was reduced to 27.47 ± 1.30% and 27.94 ± 2.23%, respectively (P < 0.01; Fig. 2B). MicroRNA-20a inhibitors were also used for proliferation assay of Panc-1 and BxPC-3. No significant difference was found between experimental groups and the control groups (P > 0.05; Supplementary Fig. S1A; Supplementary data are available online at

The effect of microRNA-20a overexpression on cell proliferation of pancreatic cancer cells.
Overexpression of microRNA-20a alters the cell cycle profile of pancreatic carcinoma cells
Flow cytometry indicated that the cell cycle distribution in the cells with overexpressed microRNA-20a was significantly different from that in the control groups, with a higher proportion of cells within the G0/G1 phase. The G0/G1 phase was increased in LV-miR-20a Panc-1 (60.8 ± 0.5% vs. 48.7 ± 0.3%) and LV-miR-20a BxPC-3 (61.3 ± 0.2% vs. 51.1 ±0.6%) compared with control cells, respectively (P < 0.05; Fig. 2C). Western blot analysis showed that cyclin D1 was inhibited in LV-miR-20a Panc-1 and LV-miR-20a BxPC-3 (Fig. 2D), but there was no significant difference between control vector and parental cells. No change was detected for cyclin E expression after overexpression of microRNA-20a (Fig. 2D). Our results were consistent with those reported previously (Yu et al., 2008).
Overexpression of microRNA-20a inhibited the migration and invasion of pancreatic carcinoma cells in vitro
An in vitro invasion system was used to evaluate the effect of microRNA-20a overexpression on the invasion of pancreatic carcinoma cells. Compared with control vector and parental cells, LV-miR-20a Panc-1 and LV-miR-20a BxPC-3 showed a significant reduction in migratory and invasive capacity. The migration rate of LV-miR-20a Panc-1 and LV-miR-20a BxPC-3 was reduced to 30.51 ± 1.15% and 24.79 ± 1.01%, and the invasion rate of the two cell lines was reduced to 34.87 ± 4.25% and 23.21 ± 3.92%, respectively (P < 0.05; Fig. 3A). The microRNA-20a-overexpressed cells showed a significant reduction in the cellular migration and invasion. Furthermore, to evaluate the effect on migration and invasion of pancreatic cancer cells when microRNA-20a was down-regulated, microRNA-20a inhibitors were used for the migration and invasion assay of Panc-1 and BxPC-3. Our data indicated that there was no significant difference between the experimental group and the control group (P > 0.05; Supplementary Fig. S1B; Supplementary data are available online at
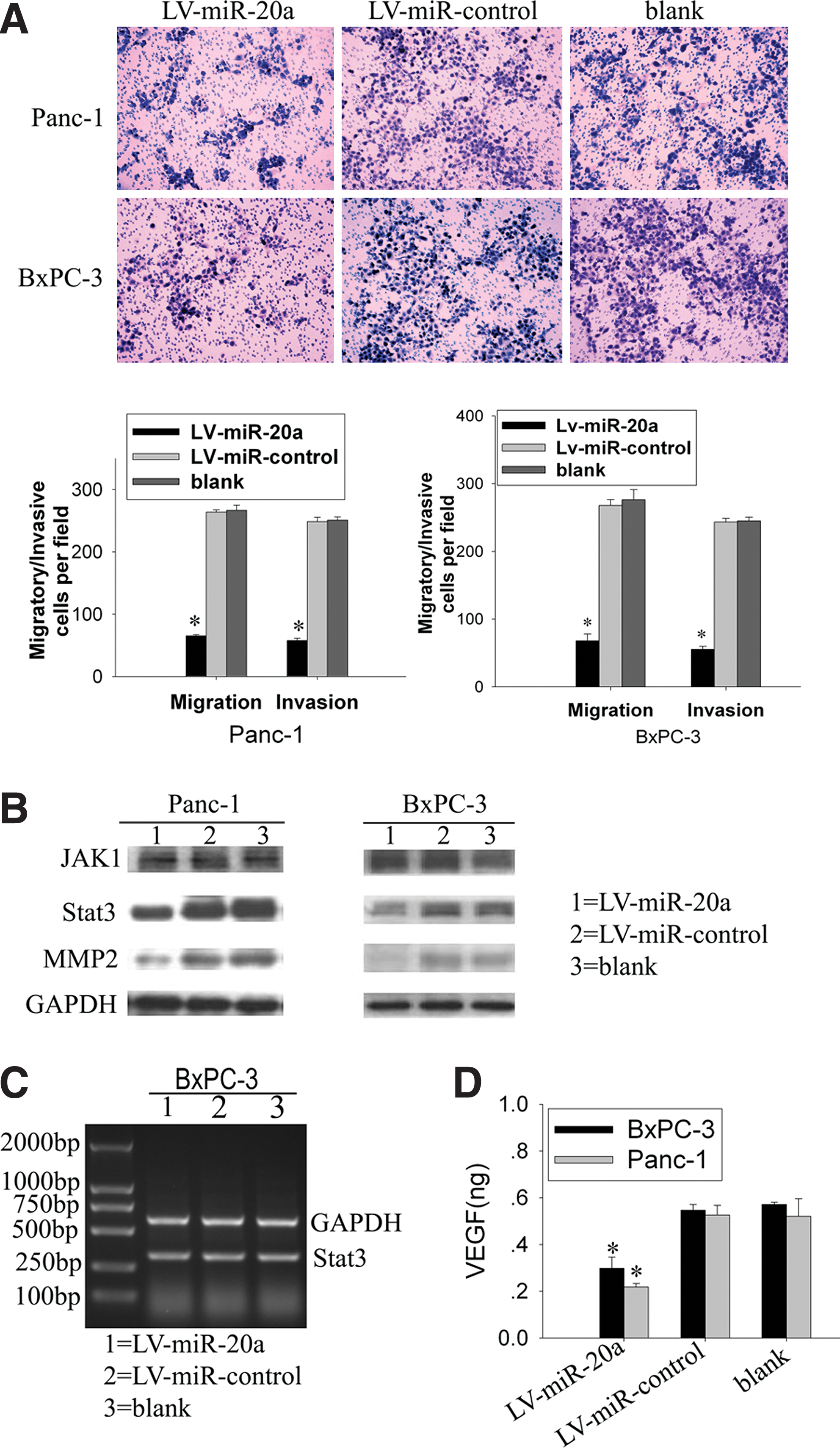
Effect of microRNA-20a on migration and invasion of pancreatic cancer cells.
MicroRNA-20a regulates Stat3 expression at the post-transcriptional level
Western blot analysis was performed in our pancreatic cancer cell lines to investigate the genes involved in pancreatic cancer metastasis. Our data showed that the Stat3 protein level was significantly reduced in LV-miR-20a Panc-1 and LV-miR-20a BxPC-3 (Fig. 3B), without any change in Stat3 mRNA (Fig. 3C). There was no significant difference in the Stat3 protein level between control vector and parental cells. Meanwhile, western blot assay did not detect any change of JAK1 protein level, which was the upstream regulatory gene of Stat3 (Fig. 3B). Our results also showed that the Stat3 protein level was decreased in a dose-dependent manner when BxPC-3 cells were transfected with 50–200 nM concentrations of microRNA-20a mimics (Fig. 4D). These data suggested that microRNA-20a negatively regulated Stat3 protein expression at the post-transcriptional level.
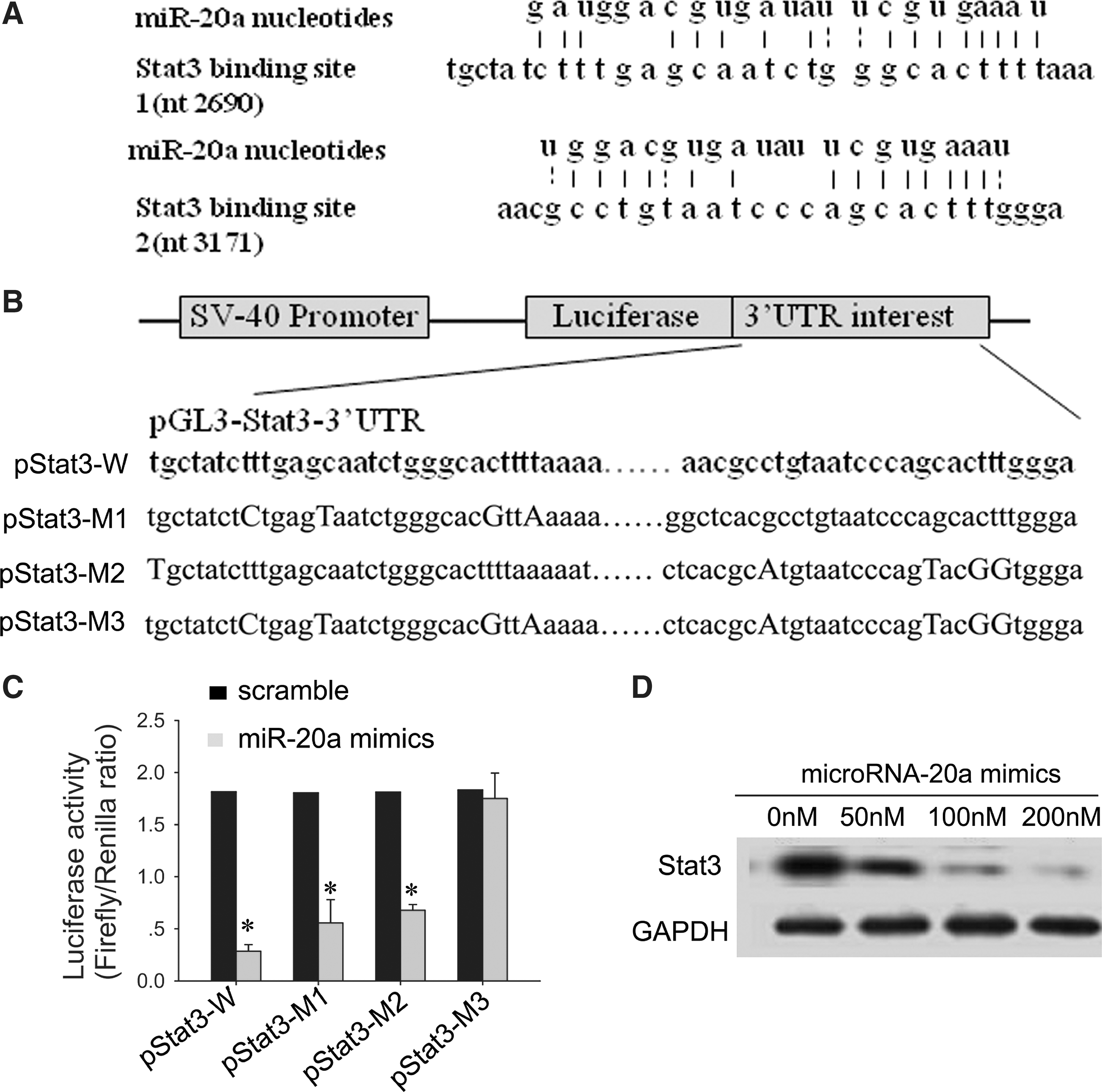
MicroRNA-20a regulates the Stat3 expression by binding the 3′-UTR of Stat3 mRNA.
Inhibition of MMP2 and VEGF expression after down-regulation of Stat3
In our results, MMP2 was also positive in pancreatic carcinoma cells (Fig. 1C and D). MMP2 and VEGF, which are the downstream target genes of Stat3 associated with tumor metastasis, were detected in our tumor cell lines. We found that MMP2 and VEGF were both down-regulated upon overexpression of microRNA-20a (Fig. 3B and D).
MicroRNA-20a regulates Stat3 by targeting the Stat3 3′-UTR
The miRBase Web site (
Overexpression of microRNA-20a inhibits the growth and metastasis of pancreatic carcinoma cells in vivo
LV-miR-20a Panc-1, LV-miR-20a BxPC-3, and their control cells mentioned previously were used to evaluate cell growth in vivo. The mice were euthanized 28 days after tumor cell injection when they had become moribund. As shown in Fig. 5A, the volume and weight of tumors resulting from injection of cells with overexpression of microRNA-20a were significantly less compared with those from the control vector groups or parental cell groups (P < 0.05). There was no significant difference in the tumor volume or in the weight between the control vector and the parental cell group in both models (P > 0.05; Fig. 5A).
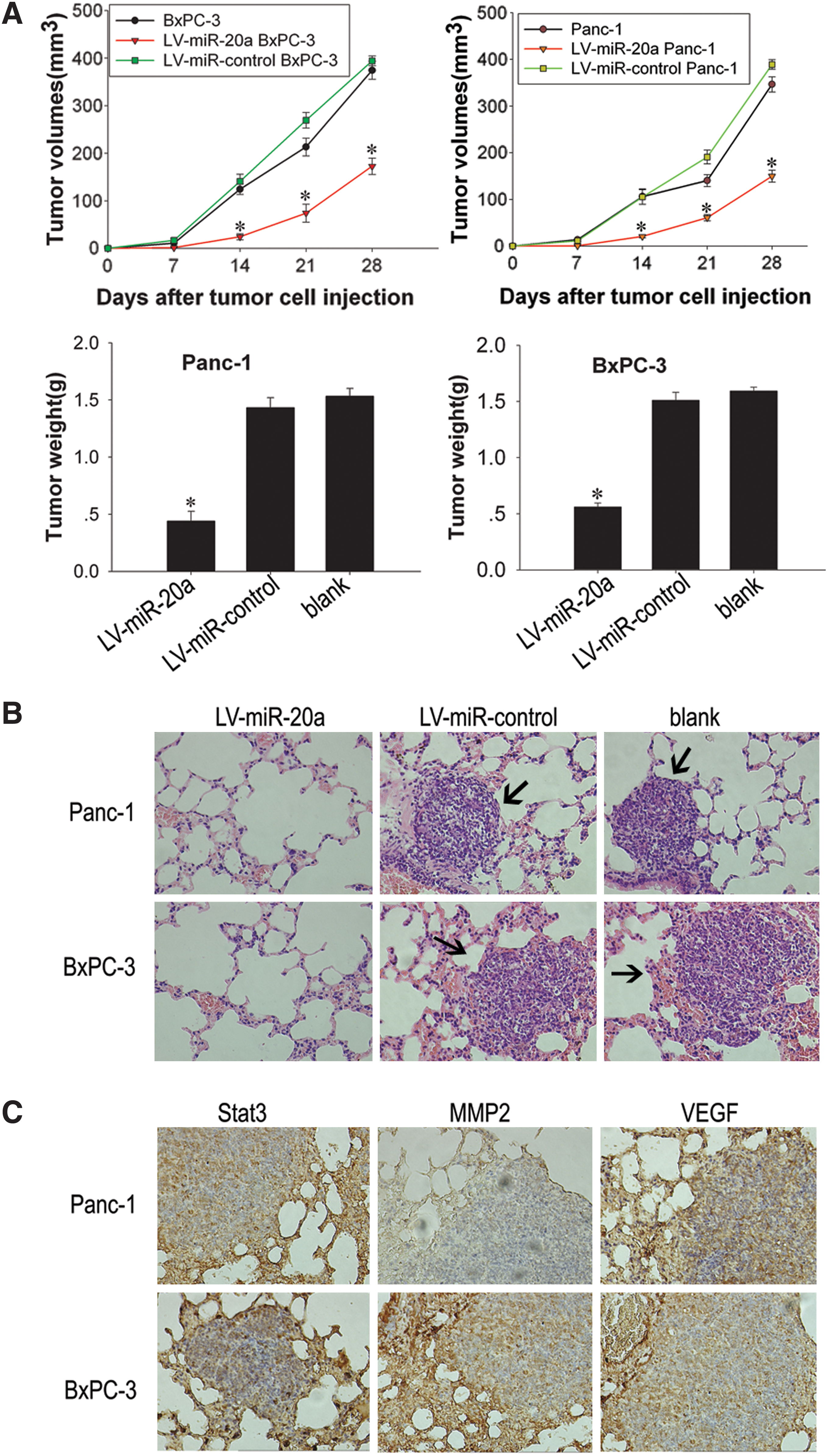
Antitumor effect of microRNA-20a overexpression on pancreatic carcinoma xenografts.
Because ectopic implantation of pancreatic cancer cells rarely leads to distant metastasis in most xenograft models, we used vena caudalis injection of tumor cells to evaluate the effects of microRNA-20a overexpression on tumor metastasis. Parental cells and control vector groups produced a large number of lung metastatic nodules in most injected mice. In contrast, injection of cells with overexpression of microRNA-20a did not produce metastatic nodules in any mice (P < 0.05; Table 1). The lung metastasis nodus was analyzed by histopathological examination with hematoxylin and eosin (H&E) staining (Fig. 5B). Immunohistochemistry analyses revealed that Stat3, MMP2, and VEGF were positive in the lung metastasis nodus of nude mice (Fig. 5C).
P < 0.01 compared with parental cells and LV-miR-control.
Discussion
In this study, we confirmed that the expression level of microRNA-20a was reduced in pancreatic cancer tissues and cell lines. We proposed that reduced expression of microRNA-20a may disrupt cell cycle control, in turn promote cell proliferation, and consequently facilitate the development of cancers like pancreatic cancer. In our study, overexpression of microRNA-20a in pancreatic cancer cell lines significantly inhibited cell growth and invasion both in vitro and in vivo. This is the first study showing that microRNA-20a regulates cellular proliferation and invasion in pancreatic carcinoma cells. Moreover, we identified that microRNA-20a negatively regulates oncogene Stat3 via binding to the 3′-UTR of Stat3 mRNA in pancreatic carcinoma cells. Together, we suggest that overexpression of microRNA-20a is a potential therapy targeting proliferation and invasion pathways in pancreatic carcinoma.
MicroRNAs play an important role in the development and maintenance of normal cellular functions. Aberrant expression of microRNAs can result in the alteration of a variety of cellular pathways through the regulation of multiple target genes (Bartel, 2004). The microRNA profiles of pancreatic cancer from different groups were not accordant (Schmittgen et al., 2004; Volinia et al., 2006; Bloomston et al., 2007; Zhang et al., 2009). TaqMan miRNA assay is a highly sensitive, specific, and precise method specific for mature microRNAs and can discriminate related microRNAs differing by as little as one nucleotide without genomic DNA contamination (Chen et al., 2005). Although TaqMan real-time PCR is an easy method for microRNA analysis, it is not suitable for detection of microRNA expression in cancer tissues due to the existence of stroma and surrounding normal tissue. In situ hybridization using the LNA probe is a useful method to visualize the microRNA expression in formalin-fixed, paraffin-embedded tissues (Kloosterman et al., 2006). Therefore, we used this approach to visualize the microRNA-20a expression in pancreatic cancer tissues. As shown in Fig. 1A, lower expression of microRNA-20a was observed in the pancreatic cancer cells compared with the matched normal pancreatic tissues. Furthermore, we used TaqMan real-time RT-PCR to detect microRNA-20a expression in pancreatic cancer cell lines. We found that the expression levels of microRNA-20a in Panc-1 and BxPC-3 were also lower than that of human immortalized pancreatic ductal cell H6C7 (Fig. 1B). The stem-loop RT primers and TaqMan probe are better than conventional ones.
According to many other published articles, we found that the expression levels and functions of microRNAs are not the same in different cancer cells. Because the target genes of microRNA are different in different cancer cells, it can work as an oncogene or a tumor suppressor. For example, in the human B cell line P493-6, the miR-17/20 cluster works as a tumor suppressor that can decrease the expression level of E2F1 and then inhibit myc-induced cell proliferation (O'Donnell et al., 2005). It also acts as a tumor suppressor in some breast cell lines by inhibiting translation of AIB1 mRNA (Hossain et al., 2006). In our study, microRNA-20a acts as a tumor suppressor in pancreatic cancer cells. However, in lung cancer, ovarian cancer, and lymphomas, the miR-17/20 cluster works as an oncogene, enhancing cell proliferation (Matsubara et al., 2007; Inomata et al., 2009; Fan et al., 2010). This discrepancy is likely due to the difference in cell context and suggests that altered expression of this microRNA may have diverse effects in tumor cells. In our study, proliferation and metastasis of pancreatic cancer cells could be inhibited by microRNA-20a overexpression, but there was no change when transfected with microRNA-20a inhibitors; this is consistent with a previous report showing that microRNA-195 expression was reduced in hepatocellular carcinoma (HCC) and acted as a tumor suppressor (Xu et al., 2009). As the same microRNA performs different functions through distinct pathways dependent on the tissue or cell type, it is important to understand the mechanisms by which microRNA-20a regulates the cell proliferation and invasion in pancreatic cancer.
Stat3, a downstream transcription factor in the JAK-STAT signal cascade, is an important player in both developmental and stem cell biology. It is a transcription factor activated by a wide range of cytokines and growth factors (Bowman et al., 2000). Stat3 is constitutively activated in human pancreatic cancer cells and directly contributes to the activation of VEGF and MMP2. Stat3 directly regulates VEGF and MMP2 expression and hence angiogenesis, growth, and metastasis of human pancreatic cancer (Wei et al., 2003; Huang et al., 2009). Conversely, inactivation of Stat3 leads to an inhibition of cell proliferation and metastasis in pancreatic cancer (MacLeod, 1999; Qiu et al., 2007). In our study, we found that down-regulation of Stat3 by overexpression of microRNA-20a significantly suppressed the expression of MMP2 and VEGF (Fig. 3B and D), which was accompanied by marked inhibition of tumor cell proliferation and metastasis both in vitro and in vivo. Our results demonstrated that inactivation of Stat3 is critical for the inhibition of invasive and metastatic behavior of pancreatic cancer. Delivery of microRNAs that are highly expressed and therefore tolerated in normal tissues but lost in cancer cells may provide a general strategy for microRNA replacement therapies (Kota et al., 2009). Overexpression of microRNA-20a may offer a novel strategy for controlling the metastatic behavior of pancreatic cancer.
Lentivirus-mediated expression of microRNAs and microRNA-specific antagomirs can induce stable gain- and loss-of-function phenotypes for individual microRNAs (Fidler, 2003; Scherr et al., 2007). We used this stable expression technology to up-regulate the microRNA-20a in human pancreatic cancer cell lines (BxPC-3 and Panc-1) and found its inhibitory effect on the proliferation and invasion behavior. We also found that microRNA-20a inhibited the expression of Stat3 in a dose-dependent manner (Fig. 4D), indicating that microRNA-20a is specifically involved in the regulation of Stat3. Furthermore, we used three different prediction algorithms to predict the binding site of microRNA-20a in the 3′-UTR of Stat3, then cloned the Stat3 3′-UTR containing the binding sites of microRNA-20a into pGL3-control vector, and cotransfected with microRNA-20a mimics. We found that luciferase activity was significantly decreased by transfection with microRNA-20a mimics (Fig. 4C). In addition, we found that Stat3 mRNA level did not change after overexpression of microRNA-20a (Fig. 3C). Therefore, decreased expression of Stat3 in cells overexpressing microRNA-20a could be partially explained by microRNA-mediated regulation, which supports the hypothesis that Stat3 is a functional target of microRNA-20a in pancreatic carcinoma. There are only three reports of microRNA-mediated regulation of Stat3 in the current literature (Krichevsky et al., 2006; Meng et al., 2007; Foshay and Gallicano, 2009). To the best of our knowledge, this is the first report to elucidate a Stat3/microRNA-20a interaction, which results in a functional regulation of pancreatic cancer cell proliferation and metastasis. The microRNA guides the RNA-induced silencing complex, or RISC, to the 3′-UTR of target mRNAs. The fate of the targeted mRNA is determined by the extent of complementary pairing between targeted mRNA and the microRNA. In one mechanism, perfect complementarity can be found between microRNA and its target mRNA, which can induce mRNA cleavage; in the other mechanism, incomplete complementarity can be found between microRNA and its target mRNA, which can cause translational repression without changing the level of mRNA. This modulation is at the post-transcriptional level. In our study, the interaction mechanism between microRNA-20a and Stat3 mRNA is the latter mechanism. In the present study, we identified microRNA-20a as a genuine regulator of oncogene Stat3 translation.
Cyclin D1 plays a key role in the regulation of the G1/S phase transition and tumorigenesis (Fu et al., 2004). In our study, we also analyzed the cell cycle and found that pancreatic cancer arrested the cell cycle at the G0/G1 phase after microRNA-20a was overexpressed, consistent with its known roles (Fig. 2C). Western blots showed that the abundance of cyclin D1 was repressed by microRNA-20a (Fig. 2D), which is consistent with the previous study showing that microRNA-20a suppressed breast cancer cell proliferation and tumor colony formation by negatively regulating cyclin D1 (Yu et al., 2008). Cyclin E, which plays a similar role as cyclin D1 in enhancing the G1/S phase transition, was not affected by overexpression of microRNA-20a in this study (Fig. 2D).
Although our novel data clearly demonstrated that modulation of microRNA-20a expression can alter the pancreatic cancer cell proliferation and invasion, several questions remain to be answered. For example, are Stat3 and cyclin D1 the only targets mRNA responsible for mediating the effects of microRNA-20a? As microRNAs exhibit promiscuous binding, it is likely that other genes associated with proliferation and invasion also are functionally down-regulated by microRNA-20a. Certainly, further investigation must be performed in the future.
In summary, we investigated the potential role of microRNA-20a in pancreatic carcinoma cell proliferation and invasion. Our data suggest an important role of microRNA-20a in the molecular etiology of cancer and implicate its potential application in pancreatic carcinoma therapy.
Footnotes
Acknowledgments
We thank Prof. Tiebang Kang (Sun Yat-sen University, Guangzhou, P.R. China) for critical reading of the manuscript and comments; and Miss Jiemin Chen and Ling Zhou (Sun Yat-sen University, Guangzhou, P.R. China) for their technical assistance. This work was supported by the National Key Basic Research Program of China (project 973, grant 2010CB912201), National High Technology Research and Development Program of China (grants 2007AA021202, 2007AA021203, and 2010CB529904), Science Research Project of Eleventh Five-Year Plan (grant 2008ZX09101-051), and National Science Fund of China (grant 30973448).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.