
Abstract
High mortality and therapy resistance of melanoma demand the development of new strategies, and overcoming apoptosis deficiency appears as particularly promising. Tumor necrosis factor–related apoptosis-inducing ligand (TRAIL) has shown high potential for apoptosis induction in melanoma cells and may be applicable for gene therapy because of its selective impact on tumor cells. We have constructed a conditional replication-competent adenoviral vector with TRAIL controlled by a tetracycline-inducible promoter (AdV-TRAIL). A variant E1A protein and the lack of E1B aimed at the restriction of viral replication to tumor cells. In particular, the replication gene E1A is controlled by a tyrosinase promoter with high selectivity for melanoma cells. AdV-TRAIL mediated strong expression of E1A and doxycycline-dependent induction of TRAIL selectively in melanoma cells, which resulted in tumor cell lysis and induction of apoptosis. In contrast, non-melanoma cells and normal human melanocytes appeared to be protected. Comparison of the AdV-TRAIL approach with a comparable CD95L vector revealed similar efficacy in vitro. In mouse xenotransplantation models, AdV-TRAIL demonstrated its activity by significant melanoma growth reduction. Melanoma cell killing by AdV-TRAIL was further improved in vitro by combinations with chemotherapeutics. We demonstrate that melanoma cells may be efficiently targeted by TRAIL-based gene therapy, and resistance may be overcome by combined chemotherapy.
Introduction
In previous work, we have characterized two principal strategies for efficient apoptosis induction in melanoma cells as proven in vitro and in mouse models. These are based on pro-apoptotic Bcl-2 proteins (Eberle and Hossini, 2008) and death ligands, respectively, as CD95 ligand (CD95L)/Fas ligand and tumor necrosis factor–related apoptosis-inducing ligand (TRAIL) (Fecker et al., 2005; Kurbanov et al., 2005; Eberle et al., 2007). Most melanoma cell lines were susceptible for conditional CD95L overexpression, which induced apoptosis, enhanced chemosensitivity, and prevented melanoma growth in mice (Eberle et al., 2003). CD95L also showed high efficacy in a conditional replication-competent adenoviral vector proving the principle suitability of death ligands for a gene therapeutic approach for melanoma (Fecker et al., 2010).
The use of CD95L is, however, highly problematic becasue of liver damage and apoptosis of hepatocytes seen in mice when treated with CD95 agonists (Ogasawara et al., 1993). The death ligand TRAIL appears as a suitable alternative. It is expressed on natural killer cells, T cells, dendritic cells, and macrophages and plays significant roles in the immune response to viral infections and in immune surveillance of tumors and metastases (Taieb et al., 2006; Falschlehner et al., 2009). In contrast to CD95L, TRAIL or TRAIL receptor agonists have shown only few side effects in vivo and in clinical trials, which was related to a selective activity in tumor cells, whereas normal cells were largely protected (Kelley and Ashkenazi, 2004; Newsom-Davis et al., 2009). Thus, suppression of tumor xenografts in mouse models without obvious signs of systemic toxicities and the fact that soluble TRAIL or agonistic antibodies to TRAIL receptors were well tolerated in clinical trials (Newsom-Davis et al., 2009; Trarbach et al., 2010) may suggest the use of this death ligand also for gene therapeutic strategies.
TRAIL binds to the agonistic death receptors TRAIL-R1/DR4 and TRAIL-R2/DR5 as well as to the decoy receptors DcR1, DcR2 and osteoprotegerin (Newsom-Davis et al., 2009). In previous studies we have demonstrated the principal responsiveness of melanoma cells to TRAIL (Kurbanov et al., 2005, 2007). As replication-deficient adenoviral vectors have frequently revealed only limited efficacy, we have constructed a melanoma cell–selective, replication-competent adenoviral vector for inducible expression of TRAIL. Its efficacy is demonstrated in vitro and in mouse models as well as in combination with chemotherapy.
Materials and Methods
Cell culture
Five human melanoma cell lines (SK-Mel-19, SK-Mel-13, Mel-2a, Mel-HO, and MeWo) were used that had revealed significant tyrosinase mRNA expression (Eberle et al., 1995). As controls, the neuroectodermal tumor cell line PFSK-1 (CRL 2060, American Type Culture Collection, Manassas, VA, USA) and the hepatoma cell line Hep-G2 (HB-8065, American Type Culture Colleciton) were examined. Human embryonal kidney HEK-293 cells were used for adenovirus amplification. Growth conditions were Dulbecco's Modified Eagle's Medium (4.5 g/l glucose) (Invitrogen, Karlsruhe, Germany), 10% fetal calf serum, and antibiotics (Biochrom, Berlin, Germany), at 37°C and 5% CO2. Cells were treated with doxycycline (Sigma-Aldrich, Taufkirchen, Germany), TRAIL (soluble human killer TRAIL, ALX-201-073-C020, Alexis, Gruenberg, Germany), doxorubicin (Enzo Life Sciences, Lörrach, Germany), vinblastine, or paclitaxel (both from Sigma-Aldrich).
Adenoviral vectors and subcloning
The adenoviral shuttle plasmid for AdV-TRAIL was constructed by substitution of a CD95L cDNA fragment of Shuttle-02 (Fecker et al., 2010). It was excised by partial EcoRI digestion and replaced by an EcoRI fragment containing TRAIL full-length cDNA that derived from pCMV-SPORT-TRAIL (IRATp970G0955D) (imaGENES, Berlin). The TRAIL full-length cDNA encodes a type II membrane protein of 281 amino acids with a theoretical molecular mass of 32.5 kDa (Pitti et al., 1996).
Shuttle-TRAIL was linearized by XbaI and ligated to an adenovirus type 5 (Ad5) long arm generated by XbaI digestion of adenovector RR5 (Marienfeld et al., 1999), which is deficient for adenoviral E1 and E3 genes, to give rise to AdV-TRAIL (Fig. 1A). It was isolated from a single viral plaque, propagated in HEK-293 cells, and purified in cesium chloride gradients as described previously (Hurtado et al., 2005). Viral titers were determined by standard plaque assays in HEK-293 cells. Identity of AdV-TRAIL was proven by polymerase chain reaction (PCR) analysis for reverse tetracycline transactivator (rtTA) and the tyrosinase promoter, whereas contaminations were excluded by PCR with Ad5 wild-type primers, as described previously (Fecker et al., 2010). In addition, PCR for TRAIL cDNA was performed using 5′-CATGGCTATGATGGAGGTCCAG-3′ and 5′-GTCAGTTAGCCAACTAAAAAGGC-3′ as primers. Used as controls were the constitutive replication-competent adenoviral vector Ad5-TRE-E1A (AdV-TRE), which drives E1A expression by a constitutive cytomegalovirus (CMV)-derived promoter (Fechner et al., 2003), and the replication-deficient Ad5-CMV-Luc (AdV-Luc), which is characterized by CMV promoter-controlled expression of luciferase (Fechner et al., 1999). For control amplification of β-actin from genomic DNA the primers 5′-ATCTGGCACCACACCTTCTA-3′ and 5′-GATGTCCACGTCACACTTCA-3′ have been used.

Selective expression of E1A and TRAIL in melanoma cells. (
Determination of apoptosis and cytotoxicity
For adenoviral transduction, cells were seeded in 24-well plates (105 cells per well). The next day, fresh growth medium was supplied containing the respective adenoviral vectors. Doxycycline (1 μg/ml) was added when promoter induction was intended. After 24 hr, apoptosis was quantified by a cell death detection enzyme-linked immunosorbent assay (Roche Diagnostics, Mannheim, Germany), which detects mono- and oligonucleosomes formed in apoptotic cells. Cytotoxicity was determined by measuring lactate dehydrogenase activity in cell culture fluids (cytotoxicity detection system; Roche Diagnostics). Both assays were performed according to the protocols of the supplier with minor modifications described previously (Eberle et al., 2003). These assays give relative values that can be compared with the values of non-treated control cells. The values of control cells were set to 1, leading to fold increases in apoptosis and cytotoxicity for the treated cells. For inhibition of caspase-mediated apoptosis, the pancaspase inhibitor Q-VD-OPh (MP Biochemicals, Heidelberg, Germany) was supplied at a concentration of 10 μM, simultaneously with AdV-TRAIL transduction.
Western blot analysis
Protein extraction and western blot analysis were performed as described previously (Eberle et al., 2003). Antibodies used were TRAIL (sc-7877, Santa Cruz, Heidelberg; diluted 1:400) CD95L (N-20, sc-834, Santa Cruz; diluted 1:400), E1A (sc-58653, Santa Cruz; diluted 1:400), glyceraldehyde 3-phosphate dehydrogenase (sc-32233; Santa Cruz; diluted 1:1,000), and β-actin (AC-15, Sigma-Aldrich; diluted 1:10,000). As secondary antibodies horseradish peroxidase–labeled goat anti-rabbit and goat anti-mouse immunoglobulins were used (DakoCytomation, Hamburg, Germany; diluted 1:5,000).
Cell surface expression of TRAIL
Cells were harvested from culture plates by treatment with 0.02% EDTA in phosphate-buffered saline (PBS). After washing with PBS, aliquots of 5 × 105 cells in 50 μl of PBS containing 1% bovine serum albumin were incubated for 30 min with a phycoerythrin-conjugated TRAIL antibody (mouse monoclonal, IgG1, RIK2, sc-56246 PE, Santa Cruz; diluted 1:5). As a control, phycoerythrin-labeled isotype IgG1 antibody (mouse monoclonal IgG1, 555749, BD Pharmingen, San Diego, CA, USA; diluted 1:5) was used. After cells were washed twice again with PBS and resuspended in PBS containing 1% paraformaldehyde, surface expression was determined with a FACSCalibur flow cytometer (Becton Dickinson, Heidelberg).
Dot blot assay for quantification of TRAIL
Aliquots of cell lysates corresponding to 1.25, 2.5, and 5 μg of total protein were dotted on a nitrocellulose membrane. For comparison, a standard curve with increasing amounts of a commercially available TRAIL preparation (ALX-201-073, Enzo Life Science) diluted in lysis buffer was applied. Blots were incubated with a TRAIL antibody (sc-7877), followed by incubation with a horseradish peroxidase–labeled secondary antibody. The chemiluminescence signals on Hyperfilm ECL (GE Healthcare, Munich, Germany) were quantified by the TINA software program (Raytest, Straubenhardt, Germany).
Cell killing assays
Cell lines were seeded in 24-well plates (105 cells per well). Growth medium was exchanged the next day, followed by adenovirus transduction. After 2 days, fresh growth medium was added. After 5 days, adherent cells were fixed with glutaraldehyde (5% in PBS, 30 min). Staining was for 30 min with crystal violet (0.01% in PBS). For quantification, bound crystal violet was dissolved in PBS containing 0.1% Tween, and the optical density at 550 nm was determined.
For real-time monitoring of viable, adherent cells the xCELLigence system (Roche, Penzberg, Germany) was used. The technique is based on microelectrodes integrated in the bottom of each well of special 96-well E-plates. The electric impedance corresponds to the cell density. Several aliquots of 5,000 cells were seeded per well, and treatment with adenoviral vectors, doxycycline, and/or chemotherapeutics was started after 24 hr. The electric impedance was determined regularly every 15 min to obtain continuous growth curves.
Viral replication assay
Cells (1 × 105 per well) were seeded in six-well plates and grown overnight. Viral transduction (multiplicity of infection [MOI] = 5) was performed for 3 hr, followed by a washing step with PBS and further incubation with growth medium. Lysates were prepared immediately after washing (3-hr values) or after 96 hr of incubation. At the time of harvesting, complete cultures were subjected to three freeze–thaw cycles for cellular lysis. Cellular debris was removed by centrifugation, and viral titers in the supernatant were determined in HEK-293 cells.
Mouse experiments
Two experimental protocols were applied in female 6–8-week-old BALB/c-nu/nu mice (M&B A/S, Ry, Denmark).
In the first, melanoma cells were suspended in PBS at 106 cells/200 μl, and 200 μl were injected subcutaneously into both flanks. After growth of small tumors (20–60 mm3), adenoviral vectors (108 PFU) were injected into the tumors every second day for a total of four injections. Following the first viral injection, mice received doxycycline with the drinking water (2 mg/ml, enriched with 50 mg/ml sucrose).
In the second protocol, melanoma cells were transduced in six-well plates (2 × 105 cells per well) with AdV-TRAIL or with the control AdV-Luc (MOI = 100). After 3 hr, cells were washed with PBS, harvested by trypsin/EDTA, washed with growth medium, and washed again twice with PBS. Then, transduced cells were pooled with a ninefold excess of non-transduced cells, before the pooled cells were injected subcutaneously into both flanks (106 cells per site, 200 μl). Doxycycline was supplied at day 3 after inoculation, as described above.
Mice were seen twice a week, to determine body weight and tumor size. The tumor volume was calculated by the formula 0.5 × length × width2. Statistical significance was determined by Student's t test, and p values of < 0.05 were considered as statistically significant. Animal experiments were performed according to the guidelines of the Federation of European Laboratory Animal Sciences Associations.
Results
Construction of AdV-TRAIL
For generation of a conditional replication-competent adenoviral vector with inducible expression of TRAIL, a shuttle plasmid (Shuttle-TRAIL) was constructed that contains a mutated variant of E1A (E1AΔpRb) under the control of a melanoma-specific tyrosinase promoter. It encodes a bidirectional expression cassette with the doxycycline/tetracycline-dependent transactivator rtTA-M2 and the full-length sequence of the death ligand TRAIL, each under control of a bidirectional tetracycline-responsive promoter (Tight1-bi) (Fig. 1A). After transient transfection in SK-Mel-13 melanoma cells, Shuttle-TRAIL revealed efficient and doxycycline-dependent induction of apoptosis (data not shown). Shuttle-TRAIL was then used for generation of the adenoviral vector AdV-TRAIL in an Ad5 background with deleted E1B and E3 genes (Fig. 1A).
Selective expression of E1A protein and inducible expression of TRAIL in melanoma cells
The functionality of the regulation elements in AdV-TRAIL was tested in four representative melanoma cell lines (SK-Mel-19, Mel-2a, Mel-HO, and MeWo). These melanoma cell lines had previously been characterized by sufficient tyrosinase expression (Eberle et al., 1995), whereas the non-melanoma cell lines Hep-G2 and PFSK-1, used as controls, were tyrosinase-negative (Fecker et al., 2005). All six cell lines had shown sufficient adenoviral transduction rates (Fecker et al., 2010). Also, according to previous work, Mel-HO and SK-Mel-19 were TRAIL-sensitive, Mel-2a was weakly sensitive, and MeWo was resistant (Kurbanov et al., 2005).
High, constitutive E1A protein expression and significant, doxycycline-inducible expression of TRAIL were seen in the four melanoma cell lines at 24 hr after AdV-TRAIL transduction (MOI = 10). TRAIL was expressed as a 34-kDa band in western blot analysis, which is in close agreement with the 281 amino acids of the full-length size of membrane-bound TRAIL and with the theoretical molecular mass of 32.5 kDa (Fig. 1B). TRAIL was also detectable on the melanoma cell surface, as shown by flow cytometry at 24 hr after transduction (Fig. 1D).
In contrast, the non-melanoma cell lines did not reveal significant expression either of E1A or of TRAIL (Fig. 1B). The lack of TRAIL expression in non-melanoma cells is explained by the requirement of E1A transactivation for initial rtTA-M2 expression and thus for starting the doxycycline-dependent expression cascade. This appears as an important step in determining the selectivity of the construct.
We further aimed to compare the suitability of the two death ligands, TRAIL and CD95L, in this gene therapeutic approach. Thus, cells were transduced in parallel with a recently described AdV-CD95L (FFE-02) (Fecker et al., 2010) at MOI = 10. Both constructs resulted in a largely comparable expression and regulation of E1A as well as of death ligand, thus proving the reliability of the vector (Fig. 1B).
For estimation of the amount of TRAIL produced by melanoma cells, total protein lysates of transduced Mel-2a, Mel-HO, and SK-Mel-19 cells (MOI = 10, 24 hr) were compared in a dot blot assay with a commercially available TRAIL preparation. At 24 hr after transduction with AdV-TRAIL and doxycycline induction, 5 μg of Mel-2a protein extract corresponded to 19 ng of TRAIL standard, 5 μg of Mel-HO corresponded to 1.8 ng of TRAIL standard, and 2.5 μg of SK-Mel-19 corresponded to 9.5 ng of TRAIL standard. The following calculation included the yield of protein extracts (Mel-2a, 441 μg; Mel-HO, 375 μg; SK-Mel-19, 125 μg) and the cell numbers at the time of harvesting (1 × 106 cells). Thus, Mel-2a cells had produced an equivalent of 167 ng/105 cells, whereas the production of Mel-HO was 13 ng/105 cells, and that of SK-Mel-19 was 47 ng/105 cells. Considering that this TRAIL preparation is usually applied at 20 ng/ml for 105 cells for induction of apoptosis, AdV-TRAIL transduction had produced a comparable and considerable amount of TRAIL. In clear contrast, no significant level of TRAIL was obtained without doxycycline or after transduction of a control vector (Fig. 1C).
Selective induction of apoptosis and selective viral replication
Induction of apoptosis as resulting from TRAIL expression was investigated as the main antitumor activity of AdV-TRAIL. The apoptotic responses of melanoma cell lines were clearly dependent on their general TRAIL sensitivity. Thus, high induction of apoptosis was seen in TRAIL-sensitive SK-Mel-19 and Mel-HO after AdV-TRAIL transduction (MOI = 1) and treatment with doxycyline, whereas apoptosis induction was weaker in Mel-2a and absent in the TRAIL-resistant cell line MeWo (Fig. 2A). It is important that no induction of apoptosis was seen in PFSK-1 and Hep-G2.
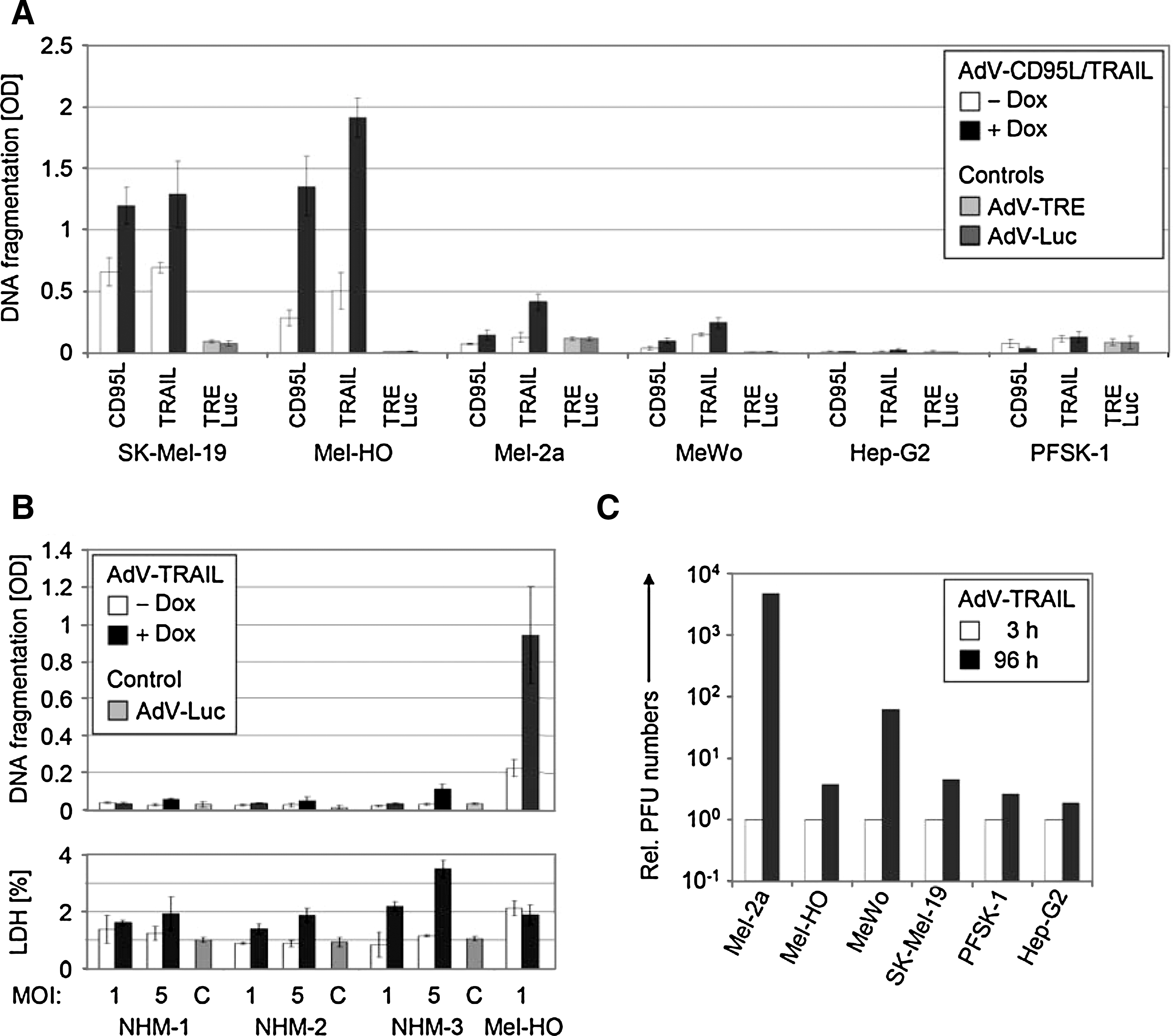
Melanoma cell-selective apoptosis and viral replication. (
Here, we were able to directly compare the efficacy of the two death ligands, TRAIL and CD95L, in an adenoviral approach in melanoma cells. When using the same MOI of 1 for AdV-TRAIL and AdV-CD95L, apoptosis induction was at a comparable level with both constructs (SK-Mel-19 and Mel-HO), or it was even higher with AdV-TRAIL (Mel-2a), thus proving the high responsiveness of melanoma cells to TRAIL. MeWo cells were resistant to both death ligands.
The stringency of doxycycline regulation of apoptosis was, however, less pronounced in SK-Mel-19 and Mel-HO, in which both constructs resulted in significantly elevated apoptosis even without doxycycline, as compared with a luciferase-encoding adenovirus and a constitutive replication-competent adenovirus without death ligand (Fig. 2A). The “leakiness” indicates the high sensitivity of melanoma cells even to low levels of death ligand (below the detection limit in western blot analysis, Fig. 1B).
Induction of apoptosis by AdV-TRAIL was also investigated in three normal human melanocyte cultures, to prove the tumor selectivity of TRAIL in our models of normal and malignant melanocytic cells. In contrast to melanoma cells, two of the normal human melanocyte cultures did not respond to AdV-TRAIL with enhanced apoptosis (up to MOI = 5), and a third culture was resistant to MOI = 1 but showed a weak response to MOI = 5. Also, cytotoxicity in response to AdV-TRAIL transduction remained at a low level (Fig. 2B).
As the second arm of AdV-TRAIL's antitumor effects, its replication competence was investigated by monitoring viral propagation. The numbers of infective particles were determined in a plaque titration assay for the six cell lines at 3 hr and at 96 hr after transduction with AdV-TRAIL at MOI = 5. Whereas there was only a weak increase for PFSK-1 and Hep-G2, the virus titers increased by 60-fold for MeWo and more than 1,000-fold for Mel-2a. For Mel-HO and SK-Mel-19, the production of viral particles was less (around a fourfold increase), which may be explained by the residual leakiness of AdV-TRAIL-mediated apoptosis that may have prevented efficient viral replication at the viral titer applied (Fig. 2C).
Selective killing of melanoma cells by AdV-TRAIL
The overall effects of AdV-TRAIL on melanoma cells (without doxycycline) were monitored in cell killing assays, which revealed dramatically reduced cell numbers of melanoma cells at 5 days after transduction, with almost no cells left when MOI = 20 were applied (Fig. 3A and B). As before, the efficacy of AdV-TRAIL was largely comparable to that of AdV-CD95L, monitored in parallel, and both death ligand–armed vectors were significantly more efficient in Mel-2a, Mel-HO, and SK-Mel-19 than a constitutive replication-competent adenovirus without death ligand (AdV-TRE). Only in the death ligand–resistant melanoma cell line MeWo did AdV-TRE exert a similarly high activity as AdV-TRAIL, which reflects the isolated oncolytic activity. In clear contrast, the growth of the non-melanoma cells was not affected by AdV-TRAIL up to MOI = 5, but they were strongly responsive to the nonselective replication-competent AdV-TRE, proving the principal susceptibility for adenovirus-mediated tumor cell lysis.
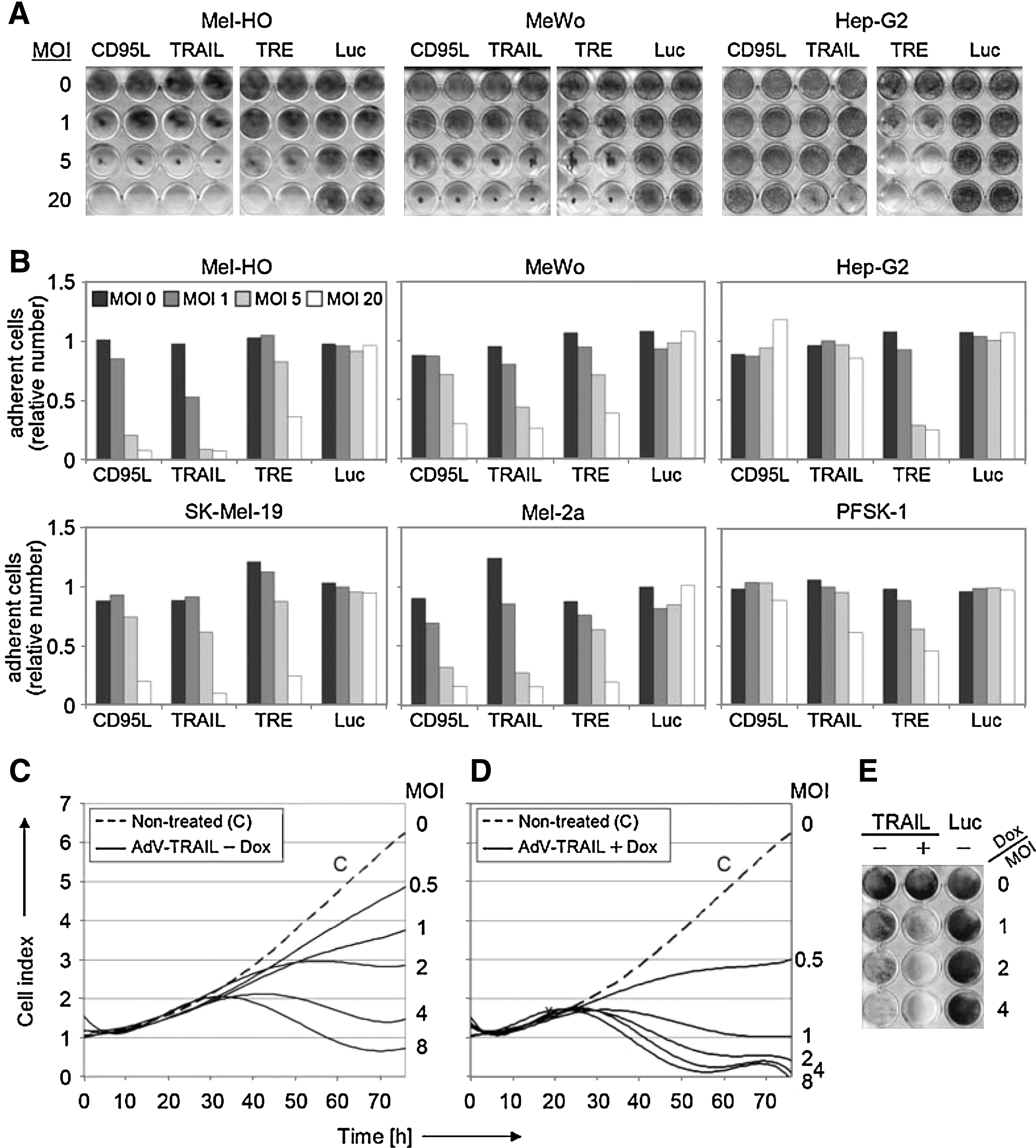
Reduced melanoma cell numbers by AdV-TRAIL. (
The mutual enhancement of adenovirus-mediated cytolysis and induction of apoptosis was investigated by real-time cell analysis, which enabled the continuous monitoring of cell growth and attachment in microtiter wells for periods of up to 1 week. Mel-HO cells revealed enhanced growth retardation with increasing concentrations of AdV-TRAIL, already without doxycycline (Fig. 3C). A further strong decrease of attached cell numbers was seen when cells were treated with doxycycline for induction of TRAIL (Fig. 3D). A similar picture was seen when Mel-HO cells were stained with crystal violet at Day 5 after AdV-TRAIL transduction (Fig. 3E). These experiments proved the mutual enhancement of the two antitumor activities.
Complementary experiments were performed with the pan-caspase inhibitor Q-VD-OPh, which has been described as completely abrogating pro-apoptotic caspase cascades (Caserta et al., 2003). Q-VD-OPh completely abolished any induction of apoptosis (Fig. 4A) and partly prevented the reduction of cell numbers in real-time cell analysis (Fig. 4B), thus proving the significance of apoptosis in AdV-TRAIL-mediated cell killing and its caspase/protease dependency.
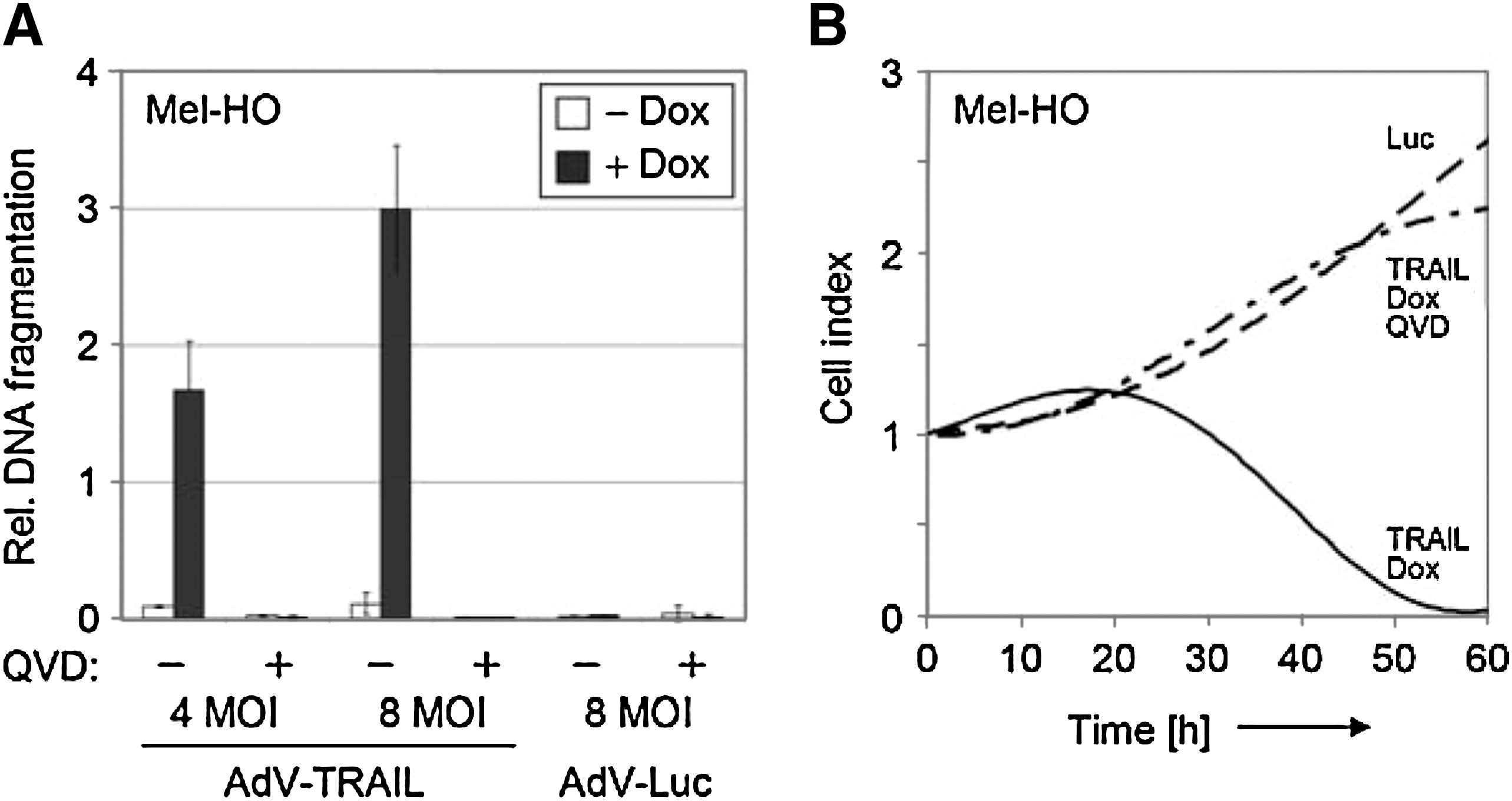
Inhibition of apoptosis and melanoma cell killing by caspase inhibition. (
Reduced melanoma growth in mouse models
The antitumoral activity of AdV-TRAIL was investigated in melanoma xenotransplants in immunodeficient mice. SK-Mel-19 and Mel-HO were not well suited for our mouse model because of either slow growth rate or poorly reproducible growth (data not shown). Thus, the experiments were performed with the moderately TRAIL-sensitive Mel-2a. In a first series of in vivo experiments, small Mel-2a tumors, which appeared at 2–6 weeks after subcutaneous melanoma cell inoculation, were injected with 108 PFU of AdV-TRAIL every second day for a total of four injections. Control tumors received corresponding amounts of the control virus AdV-Luc. Treatment with doxycycline was started after the first viral injection, for both AdV-TRAIL- and AdV-Luc-treated mice. The continuous monitoring revealed significant tumor growth retardation in response to AdV-TRAIL between days 16 and 20 (p < 0.05). A similar tendency was seen at Day 12 and between Days 24 and 28 (p < 0.1) (Fig. 5A).
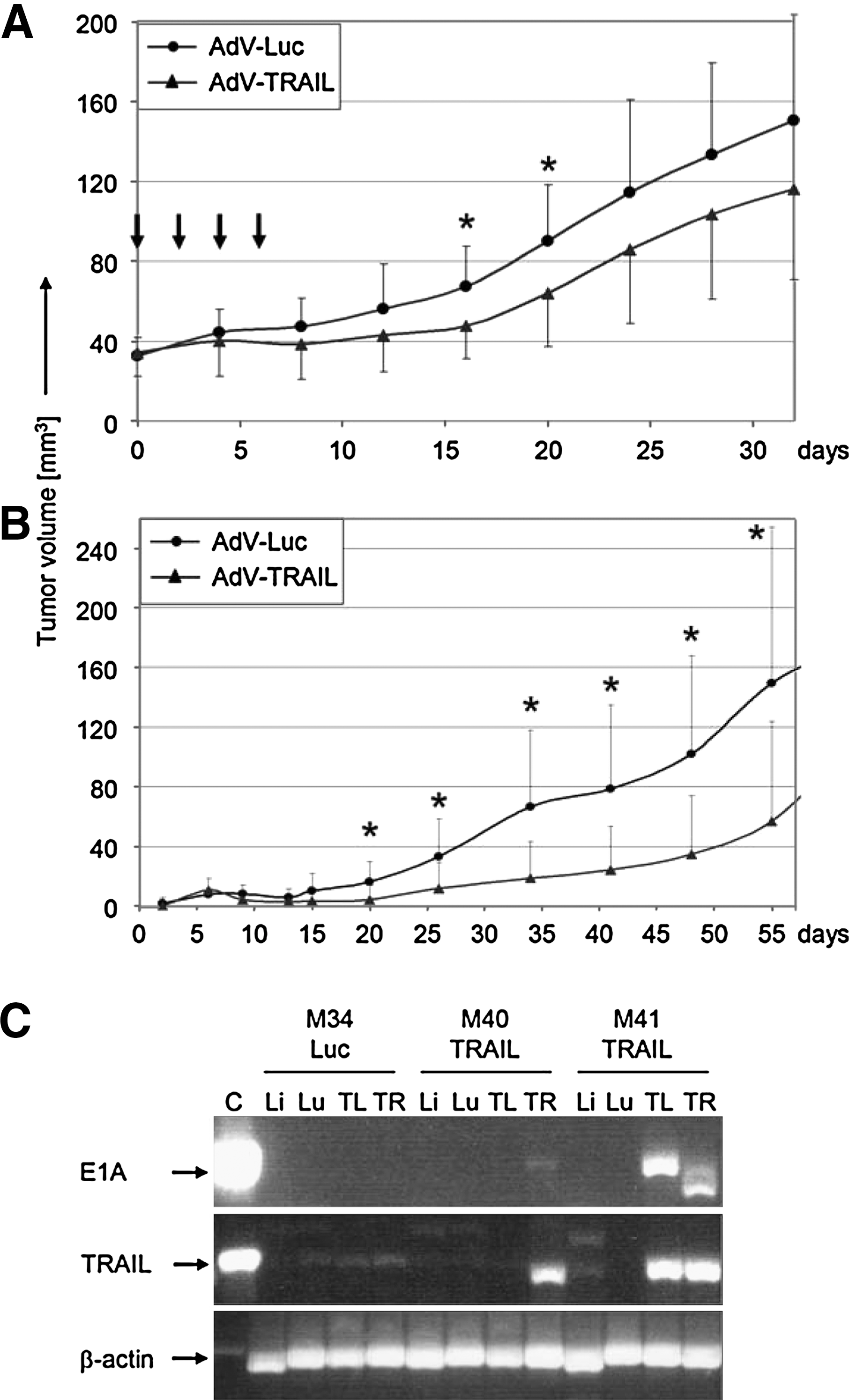
Reduced melanoma growth in nude mice. (
In another series of in vivo experiments, Mel-2a cells were transduced with AdV-TRAIL in vitro (MOI = 100) before inoculation. Then, washed, transduced cells were pooled with a 10-fold excess of non-transduced Mel-2a cells, followed by subcutaneous injection of the mixure (106 cells per site). For the control, AdV-Luc-transduced Mel-2a cells were pooled with non-transduced cells. Again, a significant delay of tumor growth and reduced mean size were seen for the use of AdV-TRAIL (p < 0.05 between Days 20 and 55, Fig. 5B). This model again proved the antitumor activity of AdV-TRAIL as well as its viral propagation in vivo.
For assessing the persistence of AdV-TRAIL in mouse tissues, samples of liver, lung, and xenotransplants were investigated by western blot and PCR analysis for E1A and TRAIL. At the time of 5 weeks after the last intratumoral injection of AdV-TRAIL (Experiment 1), no expression of TRAIL and of E1A protein was detectable by western blot analysis either in liver or lung tissue or in the injected tumors (four mice; data not shown). PCR analysis for the presence of rtTA and TRAIL DNA sequences in AdV-TRAIL revealed positivity in three of the four tumor samples (two mice), whereas one tumor sample and the normal tissues did not reveal significant amplification. No amplification of rtTA and TRAIL DNA sequences was found in a control mouse that had received AdV-Luc injections (Fig. 5C). These data indicate that AdV-TRAIL was largely removed from normal tissues with time.
Enhancement of AdV-TRAIL-mediated cell killing by combination with chemotherapy
Further attempts aimed at an enhancement of the efficacy of AdV-TRAIL in melanoma cells. The combination of soluble TRAIL with the chemotherapeutics doxorubicin, paclitaxel, and vinblastine revealed a dose-dependent enhancement of the apoptotic response in TRAIL-sensitive SK-Mel-13 as well as in weakly sensitive Mel-2a and in TRAIL-resistant MeWo melanoma cells. Whereas TRAIL-induced apoptosis was enhanced already at lower chemotherapeutic concentrations in sensitive SK-Mel-13, higher concentrations were needed for MeWo and Mel-2a (Fig. 6A). Cytotoxicity, as determined by lactate dehydrogenase release, was enhanced by chemotherapeutics at higher concentrations, but, however, was of minor effect for TRAIL (Fig. 6B). These results demonstrated that chemotherapeutic treatment was able to enhance apoptosis by TRAIL even in largely resistant melanoma cells.
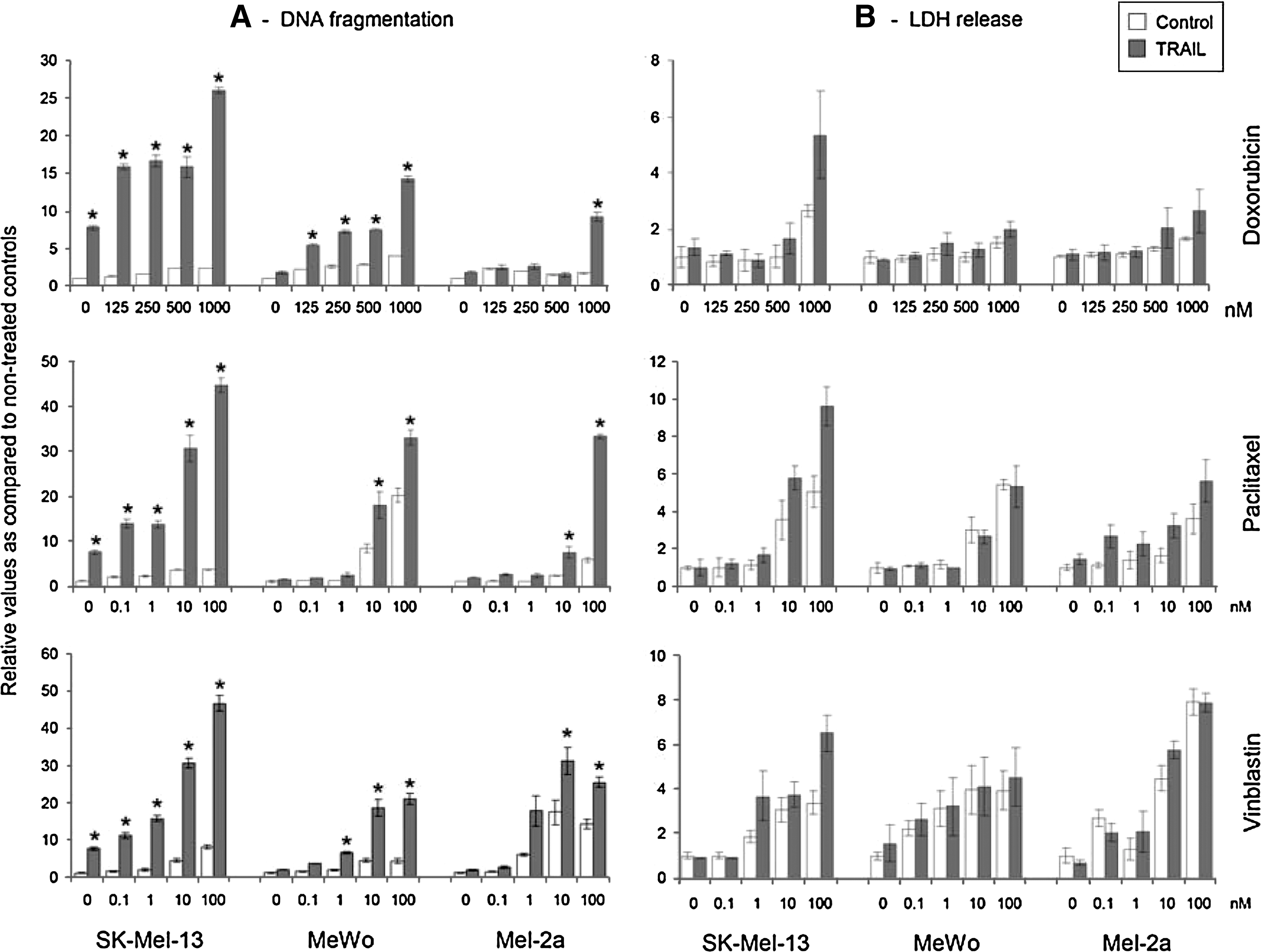
Enhanced TRAIL-induced apoptosis by combination with chemotherapeutics. (
We therefore investigated the combination of AdV-TRAIL with chemotherapeutics. SK-Mel-13 was excluded here because of low adenoviral transduction rates, and sensitive Mel-HO was used in addition to Mel-2a. When Mel-HO cells were transduced at a low MOI of 0.1–0.25, all three chemotherapeutics strongly enhanced the inhibitory effects of AdV-TRAIL on cell growth, as determined by real-time cell analysis. Similarly, combinations with doxorubicin and vinblastine were able to enhance the efficacy of AdV-TRAIL (MOI = 2) in Mel-2a (Fig. 7). These data suggested a synergistic enhancement of the activation of intrinsic apoptosis pathways by chemotherapeutics and extrinsic apoptosis pathways by death ligands and indicated strategies for enhancement of a TRAIL-mediated gene therapeutic approach.
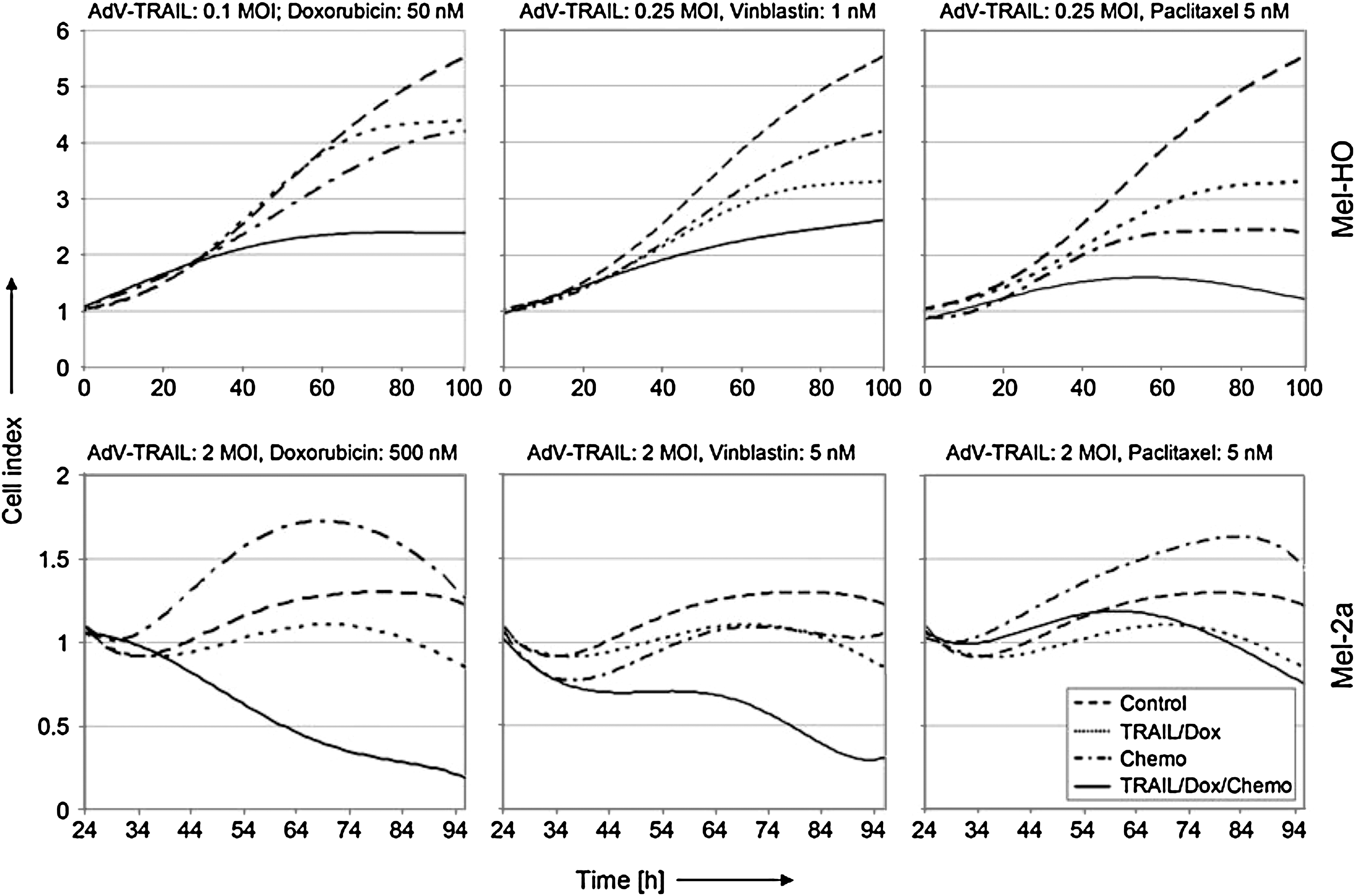
Enhancement of AdV-TRAIL effects by chemotherapeutics. Growth curves of Mel-HO and Mel-2a cells treated with AdV-TRAIL and each of the three chemotherapeutics were determined by real-time cell analysis. Cells were seeded at 5,000 per microtiter well and were treated with AdV-TRAIL and doxycycline after 24 hr (t = 0), as indicated. Chemotherapeutics (doxorubicin, vinblastine, and paclitaxel) were supplied to Mel-HO cells simultaneously with the virus, whereas Mel-2a cells received chemotherapeutics with a time gap of 24 hr. Controls received only doxycycline. Results of one of two independent experiments are shown, which consisted of duplicate and triplicate values, respectively. Both experiments revealed highly similar results.
Discussion
New concepts are needed for metastatic melanoma with regard to a complete lack of effective treatments, seen in numerous clinical trials with chemotherapy, novel small-molecule inhibitors, and combinations (Eberle et al., 2007; Garbe and Eigentler, 2007; Hersey et al., 2009). Adenoviral vectors with antitumor genes have shown efficacy in various tumor cells (Anai et al., 2006; Lebedeva et al., 2007; Song et al., 2010), and they were well tolerated in clinical trials (Alemany, 2007). As for melanoma, the clinical efficacy of antitumor genes was, however, rather limited, when replication-deficient systems had been used (Lebedeva et al., 2007; MacGill et al., 2007; Dummer et al., 2008).
Conditional replication-competent adenoviral vectors have been developed for selective tumor cell lysis. Selective replication is achieved by several strategies, which include (1) mutated E1B 55-kDa protein, which lacks the p53 inhibitory function that is required for replication in p53-wild-type cells (Bischoff et al., 1996), (2) deletion of the retinoblastoma binding site in E1A (E1AΔpRb), resulting in inhibition of viral replication in normal cells with a functional pRb-dependent regulation of cell cycle (Heise et al., 2000), or (3) control of E1A through tumor/tissue-specific promoters (Alemany, 2007). All these safety concepts are enclosed in AdV-TRAIL, namely, control of E1AΔpRb by a melanoma-specific promoter and lack of E1B.
First-generation replication-competent vectors did not reveal significant clinical efficacy, as seen for ONYX-015 in head and neck cancer, pancreatic carcinoma, and glioblastoma as well as for AdV-CG7870 in prostate cancer (Alemany, 2007). Nevertheless, an E1B-deficient adenovirus (H101) was reported as effective for head and neck tumors and has been approved in China (Xia et al., 2004). For further improvement of efficacy, conditional replication-competent adenovectors were armed with antitumor genes such as cytosine deaminase or interleukin-24 (Liu et al., 2006; Zhang et al., 2009). We have recently described an adenoviral vector for CD95L/FasL expression that resulted in efficient and selective killing of melanoma cells (Fecker et al., 2010).
Many tumor toxic agents, however, may reveal considerable side effects in vivo, like CD95L, which may result in severe liver toxicity (Ogasawara et al., 1993). TRAIL appears as particularly attractive because of selective effects in tumor cells, whereas normal cells were largely spared (Koschny et al., 2007; Newsom-Davis et al., 2009). Already in clinical trials, TRAIL and TRAIL receptor agonists have proven a safe application in patients with solid and hematological tumors (Hotte et al., 2008; Newsom-Davis et al., 2009; Trarbach et al., 2010). The use of TRAIL thus appears as a further safety element in our strategy.
Besides a good safety profile, tumor-toxic genes need high efficacy in the targeted tumor. In previous studies we have shown functioning TRAIL receptors in melanoma cells. Significant expression of both agonistic TRAIL receptors in primary melanomas, as shown by immunohistology, was further indicative of the suitability of TRAIL for melanoma treatment (Kurbanov et al., 2005). TRAIL expression by replication-deficient adenoviral vectors has been proven as effective in cell lines of prostate, breast, and bladder cancer as well as in glioma and melanoma (Griffith et al., 2000; Lin et al., 2002; Rubinchik et al., 2003). Pooled TRAIL-encoding and oncolytic adenovectors enhanced efficiency in lung, hepatocellular, colon, and squamous cell carcinoma cells (Shashkova et al., 2008; Shim et al., 2010).
Also, in conditional replication-competent adenovectors, TRAIL enhanced their efficacy in vitro and in mouse models, as seen in colon, hepatocellular, non–small cell lung, and prostate cancer cells (Sova et al., 2004; Dong et al., 2006; Jiménez et al., 2010). The AdV-TRAIL described here combines several issues for selective targeting and regulation. Thus, a melanocyte-specific tyrosinase promoter (Nettelbeck et al., 2002) was applied for selective E1A expression and viral replication in melanoma cells, whereas TRAIL expression was regulated by the inducible Tet-On system (Gossen et al., 1995). A tight regulation of the Tet-On system is, however, often impaired in adenoviral vectors (Sipo et al., 2006). The bidirectional doxycycline-regulatable expression cassette used here for induction of the transactivator rtTA and TRAIL represents a further safety issue. First, high TRAIL levels are dependent on the administration of doxycycline, and, second, induction of TRAIL was tightly restricted to melanoma cells, because of the dependence of initial rtTA expression on E1A, which was under control of the tyrosinase promoter.
TRAIL resistance appeared as a main limitation in this concept, as melanoma cells may reveal complete or partial TRAIL resistance (Kurbanov et al., 2007). MeWo was used here as a model for complete resistance, and Mel-2a for partial resistance. AdV-TRAIL efficiently induced apoptosis in sensitive Mel-HO and SK-Mel-19 as well as in weakly sensitive Mel-2a melanoma cells; in contrast, MeWo did not respond. However, cell killing assays demonstrated that these cells may at least be targeted by AdV-TRAIL through viral oncolysis. Thus, supplementation of viral oncolysis and apoptosis induction contributed to efficient and selective killing of melanoma cells by AdV-TRAIL.
As concerning normal cells, cultured normal melanocytes were largely protected from AdV-TRAIL-induced apoptosis and cytotoxicity. Cultured melanocytes are triggered to rapidly proliferate, whereas normal epidermal melanocytes are characterized by low proliferation rates and long life span (Kippenberger et al., 1997). Based on the E1AΔpRb mutation in AdV-TRAIL (Heise et al., 2000), these non-proliferating melanocytes should be even better protected in vivo. This may also apply to substantia nigra cells reported as tyrosinase-positive (Breathnach, 1988). Furthermore, an intact blood–brain barrier may prevent Ad5 penetration (Tiesjema et al., 2010). In the case of a defective blood–brain barrier in late-stage melanoma, replication of AdV-TRAIL should be largely prevented in normal cells because of E1AΔpRb and the lack of E1B (Bischoff et al., 1996; Heise et al., 2000). Even transient expression of TRAIL in these cells may be of low risk because of the tumor cell-selective induction of apoptosis by TRAIL (Kelley and Ashkenazi, 2004), which has also been discussed for glioblastoma treatment (Kuijlen et al., 2010).
Efficiency and persistence of an anti-tumor approach need to be proven in animal models. In our mouse model, direct injection of AdV-TRAIL into melanoma xenotransplants resulted in inhibited tumor growth for a period of 22 days after the last viral injection. Furthermore, subcutaneous injection of a mixture of transduced (10%) and non-transduced (90%) melanoma cells also resulted in a long-lasting reduction of tumor growth. This kind of experiment has been applied also for other tumor entities to demonstrate viral replication and spread (Fechner et al., 2003; Sova et al., 2004). Significantly reduced tumor growth and the fact that the 10% transduced cells were not overgrown by noninfected cells proved replication of AdV-TRAIL and production of infective viral particles, which had infected neighboring cells.
However, in the end no complete inhibition of tumor growth was obtained in vivo. Similar findings of only partial inhibition of tumor growth in nude mice by conditional replication-competent AdVs expressing TRAIL have been reported for hepatocellular carcinoma and colon adenocarcinoma (Pei et al., 2004; Sova et al., 2004). Acquired TRAIL resistance may limit the effect of AdV-TRAIL. Thus, we have shown previously that TRAIL-resistant cells accumulate in melanoma cell cultures after prolonged exposure to TRAIL, which coincided with downregulation of initiator caspases and of TRAIL receptors (Kurbanov et al., 2007).
Inducible TRAIL resistance appears as the reason why at the end of the experiments tumors started to grow again. Inducible TRAIL resistance of tumor cells is a major topic in apoptosis research and is regarded as a main factor limiting the success of antitumor therapy with TRAIL or agonistic TRAIL receptor antibodies (Newsom-Davis et al., 2009; Trarbach et al., 2010). Enhanced sensitivity to TRAIL has been reported to result from chemotherapeutic treatment, as shown for prostate cancer and glioblastoma cells (Shankar et al., 2005; Dorsey et al., 2009). As shown here, both TRAIL-sensitive and TRAIL-resistant melanoma cells responded with significantly enhanced apoptosis to combinations of soluble TRAIL and the chemotherapeutics doxorubicin, paclitaxel, or vinblastine. This finding could also be assigned to AdV-TRAIL, namely, enhanced cell killing was seen in TRAIL-sensitive Mel-HO and weakly sensitive Mel-2a cells, when AdV-TRAIL was combined with chemotherapeutics.
In summary, efficacy of gene therapy approaches may be enhanced with replication-competent vectors. The AdV-TRAIL described here encloses an assembly of issues for a safe application, namely, use of E1AΔpRb, lack of E1B, control of E1A by a tumor-specific promoter, use of tumor-selective TRAIL, regulation of the death ligand, and a bidirectional promoter for tight regulation. The antitumor effects are based on a combination of apoptosis and viral tumor cell lysis, resulting in efficient and selective melanoma cell killing. The limited in vivo response appears as characteristic for the highly therapy-refractory melanoma and may be based on inducible TRAIL resistance, which may be overcome by combination treatments such as by chemotherapeutics. AdV-TRAIL or related constructs may not suffice alone, but they may be further developed for complementation of chemotherapy. AdV-TRAIL appears as a proof of concept for the suitability of gene therapeutic approaches in melanoma based on replication competence and pro-apoptotic factors.
Footnotes
Acknowledgments
The study has been supported by the German Cancer Aid (Deutsche Krebshilfe) (grants 107398 and 108008).
Author Disclosure Statement
None of the authors has competing financial interests.