
Abstract
Bax inhibitor-1 (Bi-1), an anti-apoptotic protein that belongs to the Bcl-2 family, plays an important role in the mitochondrial apoptosis pathway to suppress Bax-induced apoptosis. In several human cancers, including nasopharyngeal carcinoma, its expression was found to be increased; however, up-regulated expression of this protein has been linked to increased cell proliferations. In this study, we down-regulated the gene expression of Bi-1 in nasopharyngeal carcinoma cells by using a lentivirus transfection system packed with short hairpin RNA targeting Bi-1 and used an in vivo model to assess its efficacy as a target in human gene therapy. The data indicated that human malignant nasopharyngeal carcinoma cells, CNE-1 and SUNE-1, transfected with lentiviral short hairpin RNA targeting Bi-1 grew more slowly and showed a higher degree of apoptosis. Moreover, the tumorigenicity of CNE-1 was significantly suppressed when inoculated mice were intratumorically injected with the same vector. Taken together, these data lead us to conclude that Bi-1 plays a crucial role in CNE-1 tumorigenesis and that Bi-1 may be a novel therapeutic target for nasopharyngeal carcinoma.
Introduction
Bax inhibitor-1 (Bi-1), a novel anti-apoptotic gene, formerly known as testicular-enhanced gene transcript, is localized on human chromosome 12q and codes a membrane-spanning protein with six to seven transmembrane domains and a cytoplasmic C-terminus sticking in the endoplasmatic reticulum and nuclear envelope (Kim et al., 2003; Matsumura et al., 2003). Several studies have shown that Bi-1 is overexpressed in a variety of human tumors, such as prostate carcinoma (Grzmil et al., 2003), breast carcinoma (Grzmil et al., 2006), bladder cancer (Chae et al., 2003; Andersen et al., 2004), ovarian cancer (Kawai-Yamada et al., 2004), lung cancer (Tanaka et al., 2006), cervical cancer (Watanabe et al., 2006), and brain tumors (Schmits et al., 2002). More recently, several lines of evidence have demonstrated that specific down-regulation of Bi-1 by RNA interference (RNAi) leads to cell death in several types of human carcinoma cells (Grzmil et al., 2006; Zhang et al., 2010). These findings clearly indicate that the human Bi-1 gene has the potential to serve as a prostate cancer expression marker and as a potential target for therapeutic intervention.
Gene therapy is an experimental treatment that introduces genetic material into tissues or cells for treatment of diseases. Cancer in particular ranks as a main cause of disease-related deaths, and cancer gene therapy is rapidly evolving as a possible therapeutic intervention. On the other hand, RNAi technology, a mechanism of sequence-specific inhibition of gene expression by small interfering double-stranded RNA (Hannon, 2002), is a major technique in studying the biological function of genes and their potential for use in treatment of human diseases, particularly cancer (Howard, 2009). RNAi has been successfully used as a method to silence gene expression in several in vitro and in vivo research models (Iorns et al., 2007; Chou et al., 2010) and emerging data clearly implicate RNAi as a powerful tool in gene function studies and a potential treatment for human cancers despite the concern that systematic delivery of the lentivirus-based vectors may carry the risk of germ linkage uptake (Jiang et al., 2009; Chen et al., 2010). RNAi can be carried out by different approaches; for example, transfection of chemically synthesized double-stranded RNA to cells or delivery of short hairpin RNA (shRNA) via viral vectors. Among approaches for gene transfer into cells, viral vectors in general, and lentiviral (LV) vectors in particular, for siRNA delivery are efficient natural gene delivery systems and are one of the obvious choices for cancer gene therapy.
LV vectors, such as those derived from HIV-1, show exceptional promise as gene transfer agents and have been proven to be effective vehicles for transduction of cancer cells of various organs (Castellani et al., 2010). Compared with other viral vectors, the LV vectors bear some fundamental characteristics and have several advantages including: (1) the ability to infect more easily and integrate into the host genome and determine long-term expression in animal and human xenograft models (Bartosch and Cosset, 2004; Sinn et al., 2005; Kremer et al., 2007); (2) the capacity to be easily generated and repeatedly administered without loss of efficiency (Sinn et al., 2008; Bailey et al., 2006;); and (3) higher safety and no tendency to elicit a gross inflammatory response in vitro (Morris, 2005; Copreni et al., 2009) or in vivo (Kobinger et al., 2001). Most recently, LV vectors as effective tools in human gene therapy have been approved for many human diseases (DiGiusto et al., 2010; Sumimoto and Kawakami, 2010). However, the use of an LV vector expressing shRNA targeting an oncogene as a therapeutic tool for NPC has not been demonstrated.
To develop innovative strategies for NPC gene therapy, we first developed an efficient LV shRNA (lenti-shRNA) system to specifically knock-down Bi-1 expression in NPC cells. Using this system, we investigated the function of Bi-1 in NPC tumorigenesis and evaluated the potential of Bi-1 as a therapeutic target for NPC treatment.
Materials and Methods
LV vector construction, lentivirus production, and transduction
The LV vector system (Chen et al., 2007a) was used to mediate shRNA interference. To construct an LV vector for specifically silencing the Bi-1 gene, we designed and cloned a short hairpin siRNA template into an LV vector. The shRNA targeting human Bi-1 used was 5′-GGA GAT CAA GAT TAT ATC TGG CAC TGC-3′. We synthesized DNA fragments containing GAC AGC ACA as the loop for shRNA from Proligo and cloned them into SalI and EcoRI sites of pU6 after annealing. The LV vector silencing Bi-1 was named lenti-shBi-1, and the LV vector silencing luciferase (shLuc: 5′-TGC GCT GCT GGT GCC AAC CCT ATT CT-3′) was named lenti-shLuc and used as a control.
The VSV-G pseudotyped lentiviruses were produced by cotransfecting 293T cells with the transfer vector and three packaging vectors: pMDLg/pRRE, pRSV-REV, and pCMV-VSVG (Chen et al., 2007b). The subsequent purification and concentration of virus were performed using Millipore filtration and ultracentrifugation, respectively. NPC cells (5 × 104) were plated in 24-well plates and transduced with 2 × 106 lentivirus and 8 μg/ml polybrene (Sigma).
Cell culture
The modified human embryonic kidney cells 293T were cultured in Dulbecco's modified Eagle medium (Invitrogen) with 10% fetal bovine serum supplemented with 100 U/ml penicillin and 100 μg/ml streptomycin at 37°C in a humidified incubator with 5% CO2. Nearly confluent 293T cells were used for the packaging of lentivirus silencing Bi-1 gene. The NPC cells lines CNE-1 and SUNE-1 were grown in the same conditions. After lentivirus transduction, cells were characterized by reverse-transcription PCR (RT-PCR) and Western blot analysis. Cells were continuously cultured till harvested for analysis.
Reverse-transcription PCR
Total RNA was extracted using Trizol (Invitrogen) and quantified by spectrophotometry. RNA (1 μg) was used to synthesize cDNA by using Superscript First-Strand Synthesis Kit (Promega) following the manufacturer's instructions. The mRNA level of Bi-1 on different NPC cells lines was determined by PCR. Thermal cycle conditions were as follows: 95°C for 5 min, followed by 25 cycles of 95°C for 1 min, 55°C for 1 min, and 72°C for 1 min, with a final extension at 72°C for 5 min. The PCR products were electrophoresed on 1% agarose gel, stained with ethidium bromide (Sigma), and detected by UV irradiation. In addition, we performed real-time PCR with the Applied Biosystems SYBR Green master mix kit using a Roter-Gene 300 thermal cycling instrument (Corbett Life Science). We compared Bi-1 mRNA levels and normalized them to the GAPDH mRNA levels. The primers for amplifying Bi-1, GAPDH were as follows: Bi-1 sense: 5′-CCT CTT CTG GTG GAT GCT TTG-3′, and antisense 5′-GCC TCG CTC TGT TGA TGT GA-3′; GAPDH sense: 5′-CCA GCC GAG CCA CAT CGC TC-3′, and antisense 5′-ATG AGC CCC AGC CTT CTC CAT-3′. Densitometric analyses were preformed using Scion Image software Version Cot33 (Scion).
Western blot analysis
The cells were washed three times with cold phosphate-buffered saline (PBS), collected by scraping, and lysed in 150 μl of ice-cold Tris buffer (50 mmol/L, pH 8.0) containing edetic acid 5 mmol/L, NaCl 150 mmol/L, 1% NP-40, 0.1% SDS, 1.0 mg/L aprotinin, and 0.2 mg/L phenylmethanesufonyl fluoride for 10 min. The extracts was centrifuged at 12,000 × g for 15 min, and the concentration of protein in each lysate was determined with Coomassie brilliant blue G-250 (Sigma). Loading buffer was added to each lysate, which was subsequently boiled for 3 min and then separated by SDS-PAGE. The procedures for 12% SDS-PAGE and Western blotting were described previously (Li et al., 2007). Briefly, polyvinylidene difluoride membranes (Sigma) were incubated with an appropriate dilution of the primary antibody for 1 hr (Bi-1 polyclonal antibody and actin antibody [Santa Cruz Biotechnology] were used at 1:300 and 1:500 dilution, respectively), followed by incubation with horseradish peroxidase–conjugated second-step antibody (Amersham). The results were visualized by using the enhanced chemiluminescence substrate kit (Amersham). Densitometric analyses were preformed using Scion Image software Version Cot33.
Cell proliferation assay
The effect of Bi-1 silencing on cell growth of NPC cells lines CNE-1 and SUNE-1 was examined. After lentivirus infection, samples were seeded onto a 96-well plate at a density of 5 × 103 cells per well. At indicated time points, 20 μl of MTT solution (5 mg/ml in PBS) was introduced. Four hours after incubation, 100 μl of dimethyl sulfoxide was added to dissolve the crystals. The plate was then left to stand for 10 min at room temperature and the corresponding absorbance was measured at 490 nm. Finally, we used the culture time and OD490 to construct the growth curve.
Examination of cell morphological changes
Changes in cell morphology were observed and documented for CNE-1 and SUNE-1 cells 72 hr after lentivirus transduction. Nuclear morphology of apoptosis was assessed by 4′6-diamidino-2-phenylindole (DAPI) staining by which condensed cells or cells with fragmented nuclei were identified as apoptotic cells. They were observed by using fluorescence microscopy and the images were processed by Adobe Photoshop software.
DNA fragmentation analysis
After transfection with lenti-shLuc or lenti-shBi-1 for 72 hr, total DNA in CNE-1 and CNE-SUNE-1 cells was extracted, precipitated, and quantified (Aljanabi et al., 1997). Then, the same amounts of sample DNA were electrophoresed on a 1.0% agarose gel and stained with ethidium bromide, and the DNA was detected by a UV gel image system.
In vivo studies of CNE-1 xenograft tumor models in nude mice
To study the effect of Bi-1 silencing on tumor growth in vivo, we performed two different nude mouse experiments. In the first experiment, we used 10 4- to 6-week-old female BalB/C nude mice. The mice were divided into two groups randomly (five mice per group) and injected in the flank with 2 × 106 CNE-1 cells transfected with lenti-shBi-1 and lenti-shLuc, respectively. In the second experiment, the tumor model in nude mice involved subcutaneous inoculation of 5 × 106 CNE-1 cells, followed 1 week later by infection with 5 × 107 copies of lenti-shLuc and lenti-shBi-1 in two groups, respectively. The infections were repeated three times at 2-day intervals. We monitored tumor formation and subsequent growth every other day. Size of tumor mass was monitored by averaging the diameters of the short and long axes of visible solid tumors. For all experiments, animal handling and experimental procedures were approved by the Animal Experimental Ethics Committee of the Chinese University of Hong Kong.
Statistical analysis
The results from different treatment groups were compared using the two-tailed Student's t test. Results obtained from MTT assays were expressed as mean ± SD. A p-value of less than 0.05 was considered to be statistically significant.
Results
Suppression of Bi-1 by lenti-shBi-1 in NPC cells in vitro
Though Bi-1 was found to be overexpressed in a variety of cancers (Grzmil et al. 2006; Zhang et al., 2010); however, the difference in expression of Bi-1 between NPC cells and normal nasopharyngeal cells has not been studied. By using RT-PCR analysis, we showed that Bi-1 mRNA was overexpressed in NPC CNE-1 and SUNE cells but was undetectable in NP460Tert cells (Fig. 1A, B). Subsequently, we silenced Bi-1 expression in NPC cells with a lenti-shRNA system (Chen et al., 2007a). When we infected NPC cells with lentivirus carrying shRNAs targeting Bi-1 for 72 hr, highly efficient infection (>90%) was observed as indicated by EGFP expression (Fig. 2). We monitored the mRNA level of Bi-1 in the transduced CNE-1 cells by using real-time PCR and found that the expression of Bi-1 mRNA was effectively suppressed in CNE-1 cells following infection with lenti-shBi-1 but not with lenti-shLuc (Fig. 3A). Western blot analysis further supported the effectiveness of the silencing experiment (Fig. 3B).

Bi-1 mRNA expressions in normal nasopharyngeal NP460hTert cells as well as in malignant CNE-2, CNE-1 and SUNE-1 cells. (
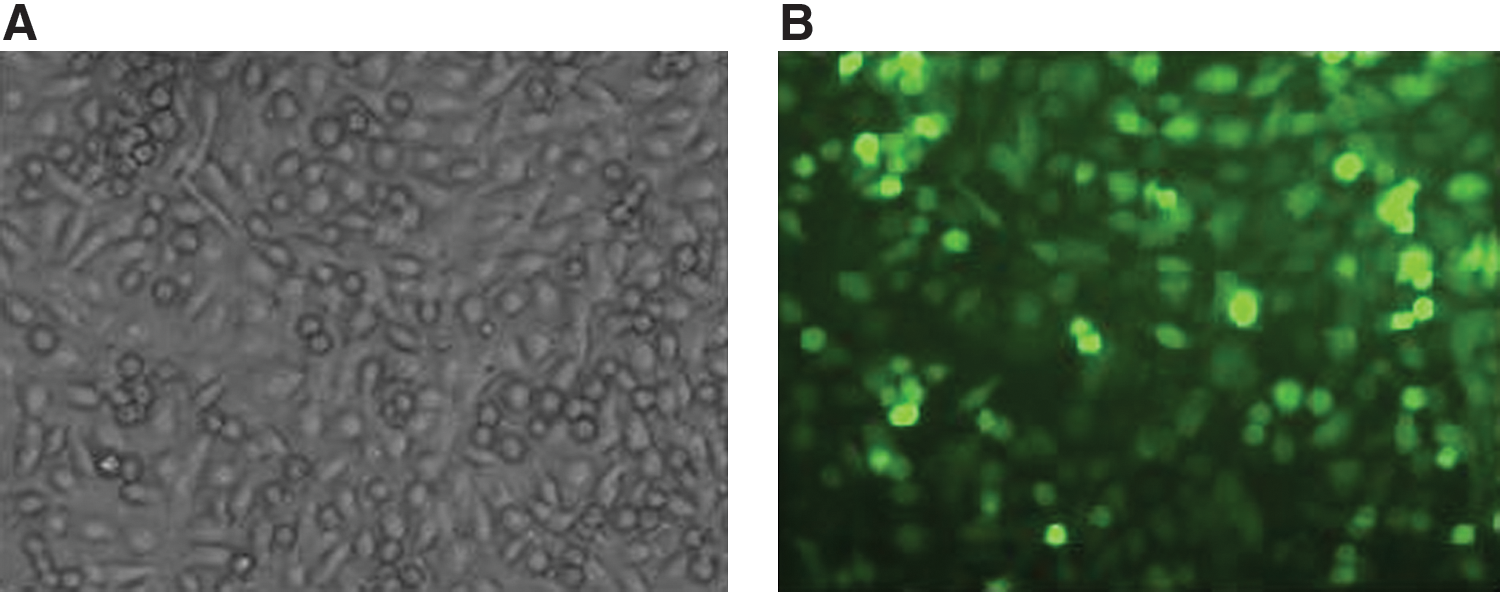
Greater than 90% transfection efficiency of lentivirus as indicated by the the percentage of CNE-1 cells with positive fluorescence signals. (

Knockdown of Bi-1 by RNAi in NPC CNE-1 cells. (
Cell proliferation significantly inhibited by lenti-shBi-1 in NPC cells
To assess the effects of lentivirus RNAi-mediated Bi-1 silencing on cell growth, we monitored cell proliferation for 5 days after cells were transfected by MTT assays. As a result we found that lenti-shBi-1 but not lenti-shLuc dramatically inhibited cell proliferation in CNE-1 and SUNE-1 cells (Fig. 4). The data suggested that the expression of Bi-1 may be essential to NPC cell growth.
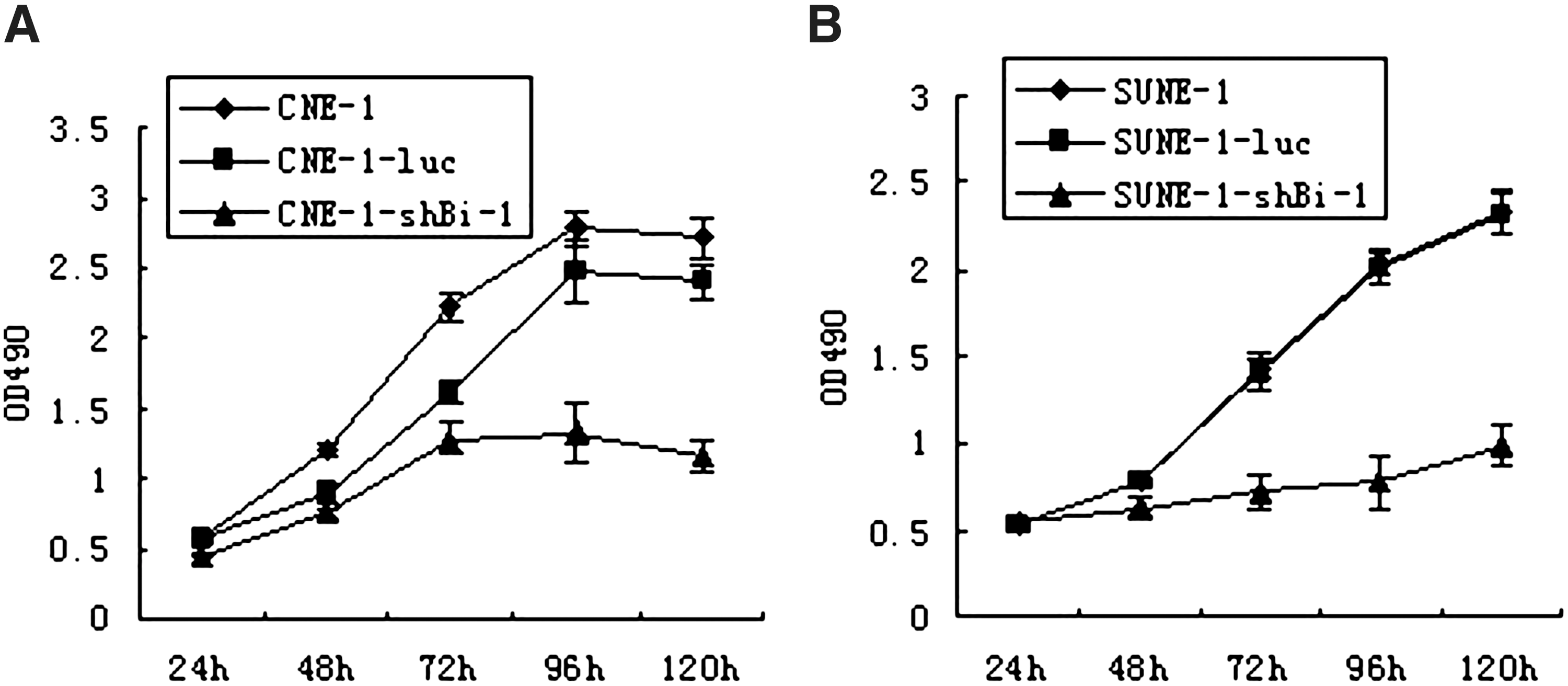
Suppression of Bi-1 by lenti-shBi-1 inhibited ex vivo NPC cell proliferation. (
Apoptosis induced by lenti-shBi-1 in NPC cells
To examine whether lenti-shBi-1 could induce cell apoptosis through knock-down Bi-1 expression in NPC cells, changes in the nuclear morphology were observed by using fluorescence microscopy after DAPI staining and DNA fragmentation analysis. Our results showed that apoptotic cells, characterized by condensed chromatin gathering at the periphery of the nuclear membrane or fragmented nuclei morphology as shown by DAPI fluorescence staining, were found in lenti-shBi-1–transduced CNE-1 and SUNE-1 cells but not in lenti-shLuc control cells (Fig. 5A). Similarly, a genomic DNA ladder formation was observed when CNE-1 and SUNE-1 cells were treated with lenti-shBi-1 for 72 hr, but not with lenti-shLuc (Fig. 5B). Taken together, these results suggested that lentivirus-mediated RNAi targeting Bi-1 can apparently suppress cell proliferation and induce cell apoptosis ex vivo.

Induction of apoptosis in NPC cells by lenti-shBi-1. (
Tumorigenicity of NPC cells reduced by lenti-shBi-1 in nude mice xenografts
To investigate the effect of Bi-1 knockdown on tumorigenesis in vivo, we inoculated the lenti-shBi-1–infected CNE-1 cells [CNE-1-RNA(+)] subcutaneously, and then compared the tumorigenicity of the cells silencing Bi-1 comparing with that of the lenti-shLuc–infected CNE-1 cells [CNE-1-RNA(−)]. At 35 days postinoculation, obvious tumors were formed in the control group mice inoculated with CNE-1-RNA(−) cells, while tumor growth was nearly completely suppressed in mice inoculated with CNE-1-RNA(+) cells. As shown in Fig. 6, we observed only small tumors in mice inoculated with CNE-1-RNA(+) cells. These observations suggested that knock-down of Bi-1 may reduce the induction of the neoplastic phenotype in vivo.

Tumorigenicity was inhibited by suppression of Bi-1 using lenti-shRNA in NPC CNE-1 cells. (
Tumor growth inhibited by lenti-shBi-1 in nude mice
To investigate whether lenti-shBi-1 targeting Bi-1 could serve as a therapeutic agent against NPC, we tested its efficacy in vivo by treating an established xenograft model with lenti-shBi-1 in nude mice. We established the NPC xenograft model by subcutaneously inoculating nude mice with 5 × 106 CNE-1 cells. After the tumor model formed, we randomly grouped mice and administered intratumoral injections of lenti-shRNAs. According to our results, tumor growth was significantly inhibited in the group treated with lenti-shBi-1 as compared with the control group treated with lenti-shLuc (Fig. 7A, B); the tumor tissue taken from nude mice was also examined through hematoxylin and eosin staining (Fig. 7C). Our results indicated that lenti-shRNA targeting Bi-1 elicited a strong antitumor effect on NPC in vivo.
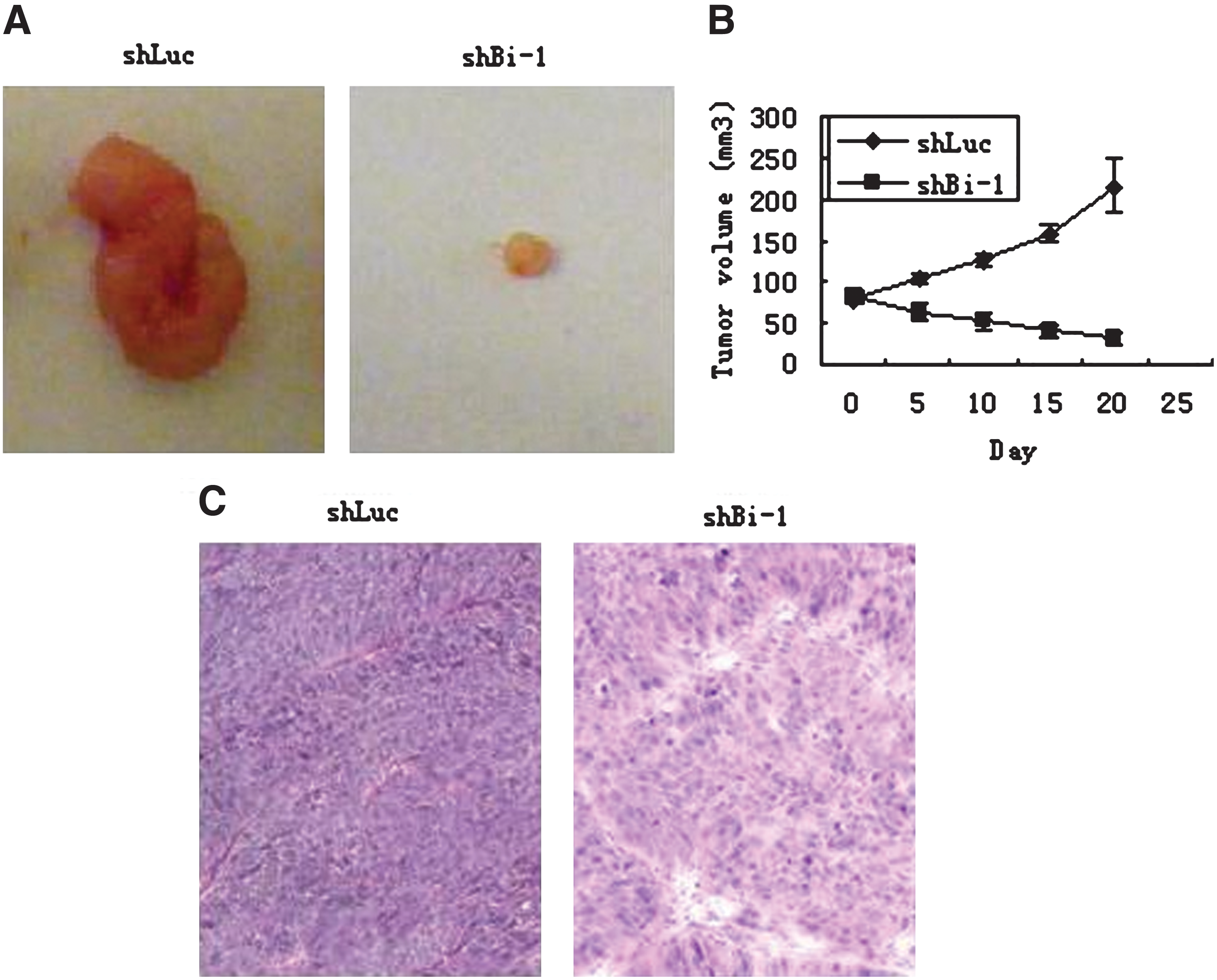
The tumor growth was suppressed by the injection of lenti-ShBi-1 in nude mice. (
Discussion
Programmed cell death is considered to be a vital phenomenon for multicellular organisms because of its involvement in removing unwanted, excessive, infected, or damaged cells to maintain tissue homeostasis (Gallaher et al., 2001). Physiologically, programmed cell death is an energy-consuming process under genetic control, and many proteins are implicated in regulating it. Evidence is emerging that the Bcl-2 protein family plays a central role in the control of programmed cell death, is composed of members inhibiting apoptosis such as Bcl-2 and Bcl-XL, and promotes apoptosis like Bax and Bak (Yang et al., 2003). Even though the pro-apoptotic proteins and the anti-apoptotic proteins appear to function independently, they can compete against one another in regulating apoptosis processes. Therefore, the ratio or balance between the pro-apoptotic proteins and the anti-apoptotic proteins appear to be more important in the regulation of apoptosis.
As a member of the Bcl-2 protein family, the anti-apoptotic protein Bi-1 has been shown to represent a new type of regulator of cell death pathways controlled by Bcl-2 and Bax (Walter et al., 1995; Huckelhoven, 2004). When overexpressed in mammalian cells, Bi-1 has been previously demonstrated to inhibit Bax-induced apoptosis, etoposide, staurosporine, and growth factor deprivation (Xu et al., 1998). These results suggest that the important role of Bi-1 in tumor progression and malignancy might be attributed to its anti-apoptotic properties. Therefore, it is important to unravel the mechanisms protecting NPC cells from undergoing apoptosis and to identify new therapeutic targets and develop new treatments.
Though RNAi is one of the most popular molecular tools in gene therapy recently, the delivery of siRNA into mammalian cells is the critical factor for a successful application of RNAi in gene function study and cancer gene therapy. To overcome this rate-limiting step, we chose an LV vector as our shRNA delivery vehicle because lentiviruses can transfect both dividing and nondividing cells with high efficiency and sustain long-term gene expression by integrating into the host genome (Kim et al., 2005; Siolas et al., 2005). Our data show that the LV expression system can easily infect over 90% of cultured cells, which is sufficient for studying the effect of RNAi on endogenous gene expression.
In the present study, we first compared the Bi-1 mRNA level in NPC cells with that in normal human nasopharyngeal cells by using RT-PCR. Our results indicated that the level of Bi-1 mRNA expression is significantly higher in NPC cells than in normal nasopharyngeal cells, which suggests that the tumorigenesis of NPC could have something to do with Bi-1 expression. In order to find the effect of Bi-1 on NPC cell growth in vitro, we next specifically suppressed Bi-1 expression by RNAi with a lentivirus expression system. In our experiment, we found that, compared with the control groups (lenti-shLuc), lenti-shBi-1 could dramatically decrease cell proliferation and viability as well as induce cell apoptosis in CNE-1 and SUNE-1 cells according to various apoptotis assays including the MTT assay, DAPI staining, and DNA laddering.
Furthermore, we conducted an in vivo experiment to explore the exact role of Bi-1 in NPC tumorigenesis. We observed an effective suppression of NPC tumorigenicity through knock-down of Bi-1 gene expression in nude mice model. Additionally, the tumorigenicity of CNE-1 cells was significantly suppressed by lenti-shBi-1 treatment after injection in an established NPC tumor xenograft mice model. These results indicated that in vivo knock-down of Bi-1 expression can inhibit the tumor growth of NPC, which suggests that Bi-1 plays an important role in NPC tumorigenesis. This further demonstrates the therapeutic value of Bi-1 inhibition for NPC treatment.
On the whole, we found that the suppression of Bi-1 through a lentivirus shRNA system could suppress ex vivo cell proliferation and in vivo tumor growth of NPC. We concluded that the Bi-1 gene could be a therapy target for NPC and an LV vector–based shRNA system may be used for clinical development in the future.
Footnotes
Acknowledgments
The authors wish to thank Ms. Hua Huang for her excellent technical assistance. This work was supported in parts by grants from the Research Grants Council of Hong Kong (RGC No. 467109 and 467507), the Special Foundation for Major Supporting Subject Program of Guangdong Province (No. GX9307), and the Planned Science and Technology Project of Guangdong Province (No. 83039).
Author Disclosure Statement
No competing financial interests exist for any of the authors.