
Abstract
Aberrant JAK/STAT3 pathway has been reported to be related to hepatocellular carcinoma (HCC) in many cell lines. In this study, a double-regulated oncolytic adenovirus vector that can replicate and induce a cytopathic effect in alpha-fetoprotein (AFP)-positive HCC cell lines with p53 dysfunction was successfully constructed. Two therapeutic genes, suppressor of cytokine signaling 3 (SOCS3) and tumor necrosis factor–related apoptosis-inducing ligand (TRAIL), were chosen and incorporated into this vector system, respectively. The combined treatment of AFP-D55-SOCS3 and AFP-D55-TRAIL (2:3 ratio) exhibited potent antitumor activity in AFP-positive HCC cell lines compared with any other treatment both in vitro and in vivo. Specific replication and low progeny yield in AFP-positive HCC cell lines rendered these double-regulated oncolytic adenoviruses remarkably safe. Our data demonstrated that restoration of SOCS3, which inhibits the JAK/STAT3 pathway, by AFP-D55-SOCS3 not only could antagonize HCC therapeutic resistance to TRAIL and adenoviruses, but could also induce cell cycle arrest in HCC cell lines. SOCS3 could down-regulate Cyclin D1 and anti-apoptotic proteins such as XIAP, Survivin, Bcl-xL, and Mcl-1, which are responsible for the synergistic inhibitory effects of AFP-D55-SOCS3 and AFP-D55-TRAIL. Dual gene and double-regulated oncolytic adenoviruses may provide safety and excellent antitumor effects for liver cancer, which is the advantage of a cancer-targeting gene virotherapy strategy.
Introduction
Constitutive JAK activation leads to persistent activation of STAT3, which activates several target genes involved in cell cycle progress and the regulation of apoptosis. In HCC, the primary factor that contributes to the aberrant JAK/STAT3 pathway is the hypermethylation of the suppressor of cytokine signaling 3 (SOCS3) promotor, which results in SOCS3 gene silencing (Weber et al., 2005). In addition, the promoter region of SOCS3 has been shown to be frequently hypermethylated in head and neck squamous cell carcinoma (Weber et al., 2005), lung cancer (He et al., 2003), and cholangiocarcinoma (Isomoto et al., 2007). SOCS3 serves as a negative regulator of the JAK/STAT pathway, and a malfunction of SOCS3 can accelerate hepatocarcinogenesis (Alexander, 2002; Riehle et al., 2008). SOCS3 overexpression mediated by adenovirus can revoke constitutive activation of STAT3, thus inducing cell cycle arrest and apoptosis (Cui et al., 2008).
Tumor necrosis factor–related apoptosis-inducing ligand (TRAIL) is a potent antitumor gene that can selectively induce apoptosis in transformed cells through the death receptor pathway (Ashkenazi and Dixit, 1999; Ashkenazi et al., 1999; Kischkel et al., 2000). Previous studies have demonstrated that HCC cells are resistant to TRAIL-induced apoptosis despite the expression of TRAIL receptors (Yamanaka et al., 2000; Shin et al., 2002). This is due to the constitutive expression of XIAP in approximately 70% of HCC cases, which inhibits the activity of caspase3, 7, and 9 (Chai et al., 2001; Suzuki et al., 2001). Previous study demonstrated that inhibiting the JAK/STAT3 signaling pathway could down-regulate XIAP expression (Fuke et al., 2007). Therefore, restoration of SOCS3 might not only antagonize HCC resistance to chemotherapy, irradiation, and oncolytic adenoviruses, but also antagonize TRAIL resistance by down-regulating XIAP.
Cancer-targeting gene virotherapy (CTGVT) is constructed by inserting an antitumor gene into an oncolytic adenoviral vector, which has a much better antitumor effect than that of gene therapy or oncolytic adenovirus alone. This is the reason that we inserted TRAIL or SCOS3 into a double-regulated oncolytic viral vector to achieve excellent antitumor effect with enhanced safety, a strategy called the cancer-targeting dual gene virotherapy (CTGVT-DG).
Materials and Methods
Cell lines and culture conditions
HEK293, an embryonic kidney cell line containing the E1A region of serotype 5 adenovirus (Ad5), was obtained from Microbix Biosystem Inc. All other cell lines were purchased from the American Type Culture Collection and included HT29 (human colorectal carcinoma), Hep3B, PLC, HepG2, Huh7, LM6, BEL7404 (human hepatocarcinoma), HeLa (human cervical carcinoma), and L02 (normal human liver fibroblast). HEK293, A549, Hep3B, PLC, HepG2, Huh7, LM6, BEL7404, and H460 cells were cultured in Dulbecco's modified essential medium supplemented with 10% heat-inactivated fetal bovine serum, 4 mM glutamine, 50 U/ml penicillin, and 50 μg/ml streptomycin. L02 cells were grown in RPMI 1640 medium containing 5% fetal bovine serum.
Virus construction, plaque assays, and virus infection
AFP-D55 was constructed in our lab. Double-regulated oncolytic adenovirus AFP-D55 carrying enhanced green fluorescent protein (EGFP) (AFP-D55-EGFP), ZD55 carrying TRAIL (AFP-D55-TRAIL), and SOCS3 (ZD55- SOCS3) were constructed and described previously (Zhang et al., 2003). The viruses were plaque purified, propagated on HEK293 cells, and purified by centrifugation banding on cesium chloride by standard procedures. Titers of adenoviruses were determined by TCID50 assay on HEK293 cells (Barrett et al., 1996; LaBarre and Lowy, 2001). Cells were infected with adenoviruses at different doses at 37°C. Cell viability was evaluated by a standard MTT assay (Sigma).
Western blot analysis
To determine the expression of various proteins and protein dynamic change in infected cells, Western blot analysis was performed as described previously. Cells were harvested by trypsinization and resuspended in lysis buffer (62.5 mM Tris-HCl pH 6.8, 2% sodium dodecyl sulfate [SDS], 10 mM glycerol, 1.55% DTT). The total protein concentration was determined by BCA protein assay kit (Thermo). Protein samples were the separated by 12% SDS-PAGE and transferred to nitrocellulose membranes (Millipore). Membranes were blocked in 5% bovine serum albumin (BSA) and incubated with primary antibodies, which were detected by the appropriate secondary antibodies and revealed with an infrared imaging system (Odyssey). The primary antibodies used were rabbit polyclonal anti-SOCS3, anti-STAT3, mouse polyclonal anti-Mcl1, goat polyclonal anti-Survivin, anti-TRAIL (Abcam), mouse polyclonal anti-pSTST3, anti-XIAP anti-Bcl-xL (R&D Systems), rabbit polyclonal anti-caspase-8, anti-caspase-9, anti-caspase-3, anti-PARP, and anti-β-actin, mouse polyclonal anti-Cyclin D1 (Santa Cruz Biotechnology).
Cytopathic effect assay
Tumor cell lines (HT29, HeLa, Huh7, HepG2, PLC, Hep3B, BEl7404, LM6) and a normal cell line (L02) were infected with AFP-D55, AFP-D55-SOCS3, AFP-D55-TRAIL, or a combination of AFP-D55-SOCS3 and AFP-D55-TRAIL (2:3) at various multiplicities of infection (MOI), respectively. After 5 days, cells were exposed to 2% crystal violet in 20% methanol for 10 min and washed. The results were documented as photographs.
Cell viability assay
Tumor cell lines (HT29, HeLa, Huh7, HepG2, PLC, Hep3B, BEl7404, LM6) and the normal cell line (L02) were plated in 96-well plates and treated with AFP-D55, AFP-D55-SOCS3, AFP-D55-TRAIL, or a combination of AFP-D55-SOCS3 and AFP-D55-TRAIL (2:3) at 10 MOI, respectively. At the indicated time, medium was removed, and 20 μl of 0.5 mg/ml MTT was added to each well. The cells were incubated at 37°C for 4 hr, and then MTT solution was evacuated. One hundred microliters of DMSO was added to each well and mixed thoroughly. The absorbance from the plates was read on a Bio-Rad Microplate Reader Model 655 at 595 nm.
Measurement of apoptosis in vitro
For the Hoechst staining assay, Huh7 or L02 cells were seeded on chamber slides and treated with PBS or viruses at a MOI of 5. After 48 hr, cells were stained with Hoechst 33258 (Molecular Probes) and observed under the fluorescent microscope. For the flow cytometry analysis, cells after treatment were trypsinized, washed once with PBS, and stained with fluorescein isothiocyanate–labeled annexin V and propidium iodide (BD Biosciences). Fluorescence was analyzed using a FACSort flow cytometer (Becton Dickinson).
In vivo animal experiments
All animals used in these experiments were maintained in the institutional facilities in accordance with regulations and standards of the U.S. Department of Agriculture and the National Institutes of Health. Female Balb/c nude mice at 4–5 weeks obtained from the Animal Research Committee of the Institute of Biochemistry and Cell Biology (Shanghai, China) were used in all of the experiments. The tumor xenograft model was established by injecting 2 × 106 Huh7 cells into the right flank of male Balb/c nude mice. Each group comprised eight animals, and the tumor growth was monitored and measured every 3 days. Tumor volume (V) was calculated by the formula: V (mm3) = length × width2/2. When the tumors were 80–100 mm3 in size, mice were randomized into five groups and a daily dose of 6.67 × 108 plaque-forming units (PFU) of the viruses suspended in 100 μl of PBS or 100 μl of PBS alone was administrated intratumorally for 3 days. At the end of the experiments, the tumors were harvested for histopathological, immunohistochemical, and TUNEL analysis.
Histopathology and immunohistochemical staining
Tumors or livers were harvested and fixed in 4% paraformaldehyde, embedded in paraffin, and cut into 4-μm sections. For histopathology analysis, the paraffin sections of livers or tumors were stained with hematoxylin and eosin (HE). For immunohistochemistry analysis, these sections were stained with monoclonal anti-Hexon, anti-SOCS3, and anti-TRAIL antibodies at a 1:500 dilution (Abcam). The slides were then washed with PBS and incubated with the avidin–biotin–peroxidase complex reagent (Vector Laboratories) and detected with diaminobenzidine tetrahydrochloride (DAB) solution containing 0.006% H2O2. Hematoxylin was used as a counterstain. Tissue sections stained without primary antibodies were used as negative controls.
TUNEL assay
To detect the apoptotic cells in tumor specimens, an in situ cell apoptosis detection kit (Clontech) was used. The tumor sections were deparaffinized in xylene and rehydrated through decreasing concentrations of ethanol. After incubation with 20 μg/ml proteinase K and 0.3% H2O2, the sections were washed with PBS and incubated with equilibration buffer. The Biotin-11-dUTP and TdT was added in a humidified chamber at 37°C for 60 min, and the cells were combined with avidin–horseradish peroxidase at 37°C for 60 min and colorized with DAB. Hematoxylin was used as a counterstain.
Statistical analysis
The data reported in this article represent the means of three independent experiments and the bars show the standard deviation. Student's t test was used to calculate the statistical significance of the experimental results. The significance level was set at p < 0.05.
Results
Construction and characterization of the AFP-targeting oncolytic adenovirus
AFP-D55 is a double-regulated oncolytic adenovirus, whose E1A gene is driven by an AFP promoter with a SV40 enhancer upstream and whose E1B 55 kDa gene was deleted. Three genes, EGFP, TRAIL, and SOCS3, were introduced into AFP-D55, respectively, yielding AFP-D55-EGFP, AFP-D55-TRAIL, and AFP-D55-SOCS3 (Fig. 1a). As shown in Fig. 1b, AFP-D55, AFP-D55-EGFP, AFP-D55-TRAIL, and AFP-D55-SOCS3 expressed E1A protein but failed to express E1B 55 kDa protein. In contrast, wild-type Ad5 expressed both proteins. Only AFP-D55-SOCS3 expressed SOCS3 protein, and only AFP-D55-TRAIL expressed TRAIL protein. These data indicate that AFP-D55, AFP-D55-EGFP (Figs. 1b, 3b), AFP-D55-TRAIL, and AFP-D55-SOCS3 have correct structures, and foreign genes can be expressed.
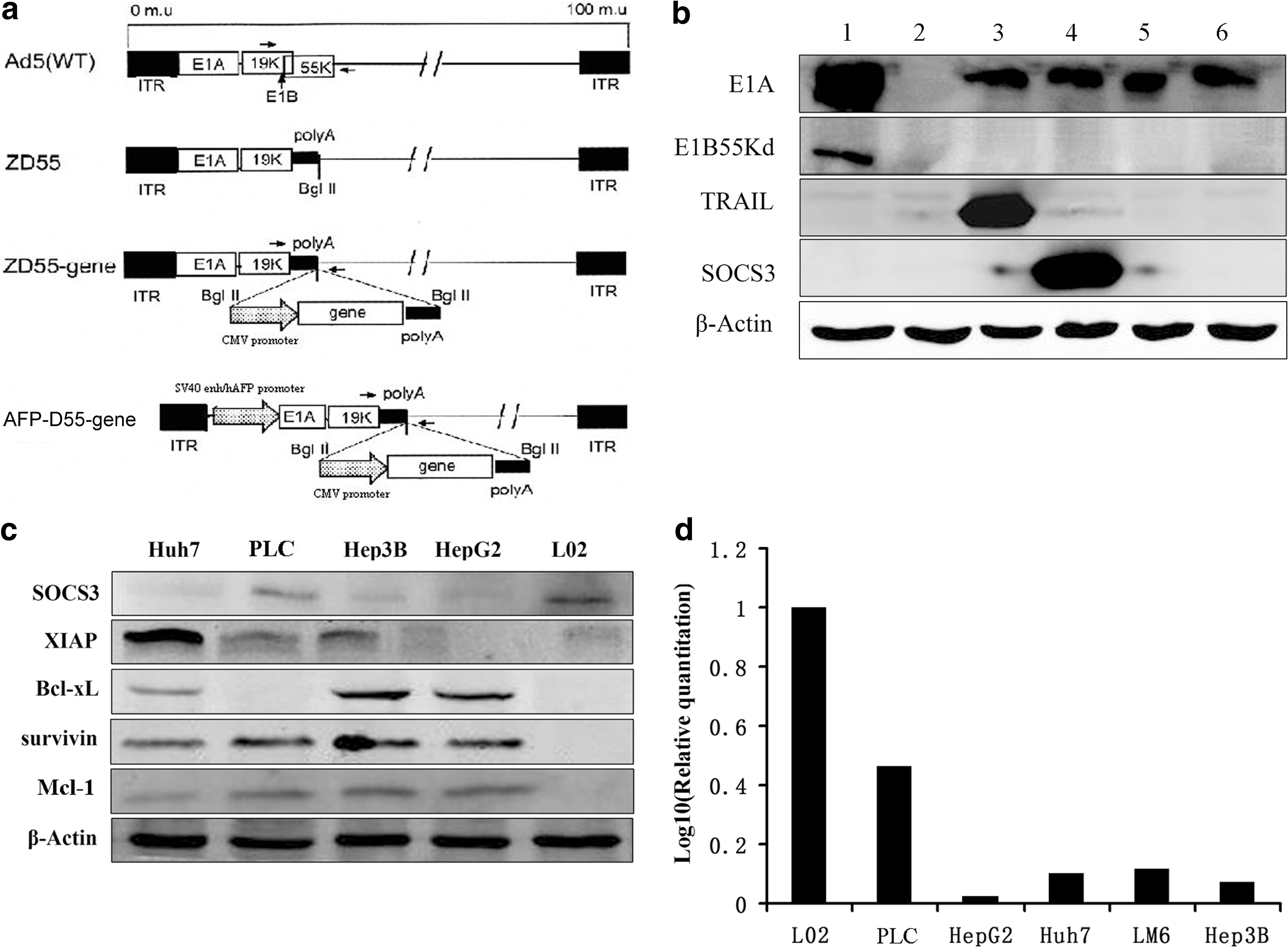
Characterization of adenoviruses and aberrant protein expressions in hepatocellular carcinoma (HCC) cell lines.
The SOCS3 was down-regulated and XIAP, Survivin, Bcl-xL, and Mcl-1 were overexpressed in some HCC cell lines.
As shown in Fig. 1c, the expression of SOCS3 was dysregulated in several HCC cell lines (Huh7, PLC, Hep3B, HepG2) when compared with the normal human liver cell line L02 at the protein expression level, a finding also observed at the mRNA expression level (Fig. 1d). Furthermore, overexpression of XIAP, Survivin, Bcl-xL, and Mcl-1 coincided with down-regulation expression of SOCS3 in these HCC cell lines compared with normal human liver cell line L02 (Fig. 1c).
Selective replication of AFP-D55, AFP-D55-SOCS3, and AFP-D55-TRAIL in vitro
The replication of these double-regulated oncolytic adenoviruses, AFP-D55, AFP-D55-SOCS3, and AFP-D55-TRAIL, were evaluated by progeny assay in both AFP-positive HCC cell lines, such as Huh7, Hep3B, HepG2, PLC, and AFP-negative HCC cell lines BEL7404 and LM6, human cervical carcinoma cell line HeLa, human colorectal carcinoma cell line HT29, and human normal liver cell line L02. ZD55, previously constructed in our lab, was used as the control (Fig. 2a). In all the AFP-positive HCC cell lines tested, the selective replication of AFP-D55, AFP-D55-SOCS3, and AFP-D55-TRAIL increased by 12- to 28-fold 48 hr after infection, which was strikingly weaker than ZD55 (p < 0.01). In these AFP-negative cell lines, AFP-D55, AFP-D55-SOCS3, and AFP-D55-TRAIL basically did not replicate, whereas ZD55 replicated continuously (p < 0.01). Although these double-regulated oncolytic adenoviruses were low in progeny yield, it was potent enough to drive the expression of exogenous genes (Fig. 2b). These data demonstrate that the safety concerns of the recombinant adenovirus AFP-D55, AFP-D55-SOCS3, and AFP-D55-TRAIL were remarkably balanced.
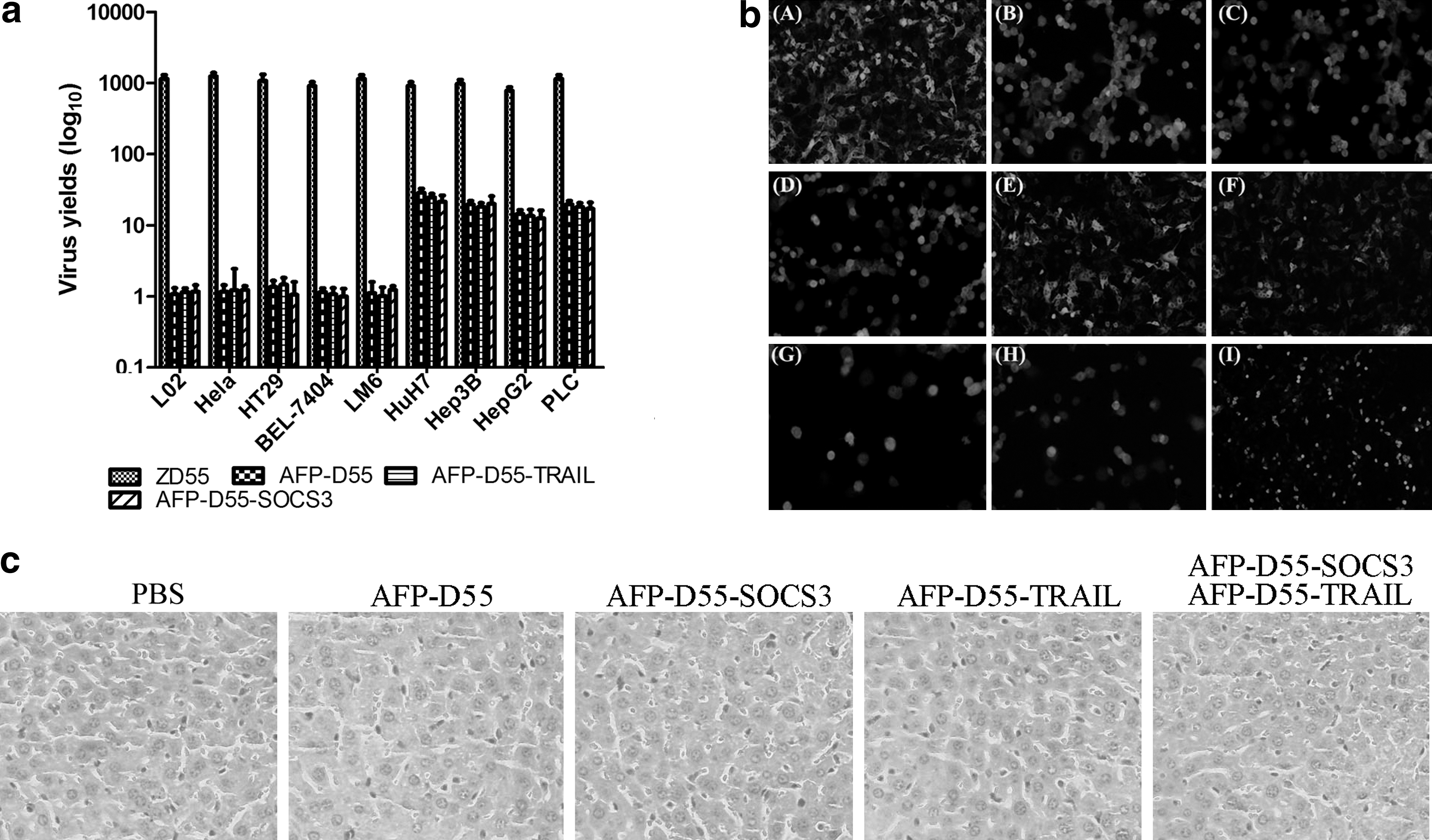
Safety of the double-regulated oncolytic adenoviruses.
Safety of the double-regulated oncolytic adenoviruses
To further evaluate the safety of the double-regulated oncolytic adenoviruses, serum levels and liver histopathological levels of hepatotoxicity were examined. For serum levels of hepatotoxicity, alanine aminotransferase, aspartate aminotransferase, and alkaline phosphatase were tested. No obvious hepatotoxicity was found in any of the double-regulated oncolytic adenovirus-treated groups compared with human Ad5 (p < 0.01), although it seemed that the hepatoxicities of these double-regulated oncolytic adenoviruses were higher than those of normal or PBS-treated groups (p < 0.01) (Table 1). But with regard to histopathological hepatotoxicity, there was no difference between the double-regulated oncolytic adenovirus-treated groups and PBS-treated groups (Fig. 2c).
Serum levels of alanine aminotransferase (ALT), aspartate aminotransferase (AST), and alkaline phosphatase (ALP) were used to test for hepatotoxicity. Hepatic function panel in mice 96 hr after receiving different drugs. Each treatment group comprised 3 BALB/c (athymic nude) mice.
SEM, standard error of the mean.
Antitumor effect of the combined treatment of AFP-D55-SOCS3 and AFP-D55-TRAIL in vitro
Several human hepatocellular carcinoma cell lines (Huh7, Hep3B, HepG2, LM6, PLC, BEL7404), one human cervical carcinoma cell line (HeLa), one human colorectal carcinoma cell line HT29, and a normal human liver cell line (L02) were infected with AFP-D55, AFP-D55-SOCS3, AFP-D55-TRAIL, and a combination of AFP-D55-SOCS3 and AFP-D55-TRAIL (2:3). A cytopathic effect (CPE) assay was performed in vitro, and the results are shown in Fig. 3a. For different AFP-positive HCC cell lines, the cytopathic effect seemed to be different for AFP-D55-SOCS3 or AFP-D55-TRAIL alone. However, the combination treatment of AFP-D55-SOCS3 and AFP-D55-TRAIL exhibited synergistic CPE. Little or no CPE was observed in cell lines treated with AFP-D55. Furthermore, in these AFP-negative cell lines, there was no significant difference in the CPE observed in all treatments.
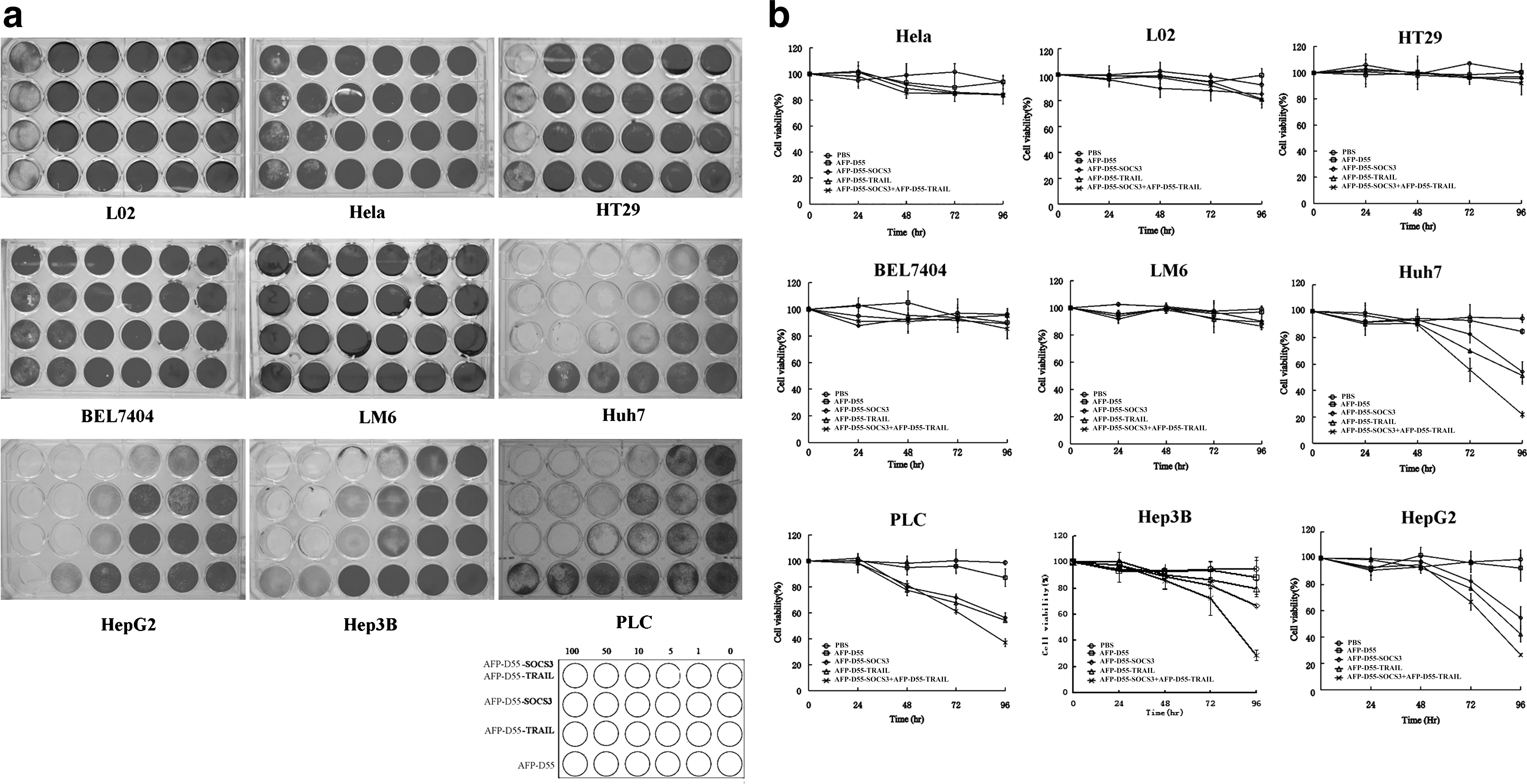
Antitumor effect of the combined treatment of AFP-D55-SOCS3 and AFP-D55-TRAIL in vitro.
To further evaluate the kinetics of CPE induced by these treatments, these cell lines were plated in 96-well plates and infected with AFP-D55, AFP-D55-SOCS3, and AFP-D55-TRAIL, as well as the combination of AFP-D55-SOCS3 and AFP-D55-TRAIL (2:3). As shown in Fig. 3b, at the same MOI of 10, daily analysis for cell viability demonstrated the inhibition of cell proliferation in AFP-positive HCC cell lines treated with AFP-D55-SOCS3 and AFP-D55-TRAIL was more significant than AFP-D55 and AD-SOCS3 or AFP-D55-TRAIL alone (p < 0.01), with a cell viability of 21%–37% on day 4. All treatments had little or no inhibition of cell proliferation on these AFP-negative cell lines (p > 0.05).
Antitumor effect of the combined treatment of AFP-D55-SOCS3 and AFP-D55-TRAIL in vivo
To validate the efficacy of the combined treatment of AFP-D55-SOCS3 and AFP-D55-TRAIL, which resulted in a potent antitumor effect when compared with either AFP-D55-SOCS3 or AFP-D55-TRAIL treatment given singly, a tumor model in nude mice bearing human HCC Huh7 was established by implantation of Huh7 cells in the right flank of the animals. When the tumors grew to 100–120 mm3, they were treated with PBS, AFP-D55, AFP-D55-SOCS3, AFP-D55-TRAIL, and a combination of AFP-D55-SOCS3 and AFP-D55-TRAIL (2:3) by intratumoral injection daily for a total virus dosage of 2 × 109 PFU for 3 days. The tumor growth was inhibited in each group treated with viruses as compared with the negative control group treated with PBS (Fig. 4a). Interestingly, the tumor growth inhibition in the combined treatment group was much stronger than that of either AFP-D55-SOCS3 or AFP-D55-TRAIL treatment alone, with about 89% of the tumor growth being inhibited 61 days after inoculation. All the tumors were then removed from the mice and weighed (Fig. 4c, d).
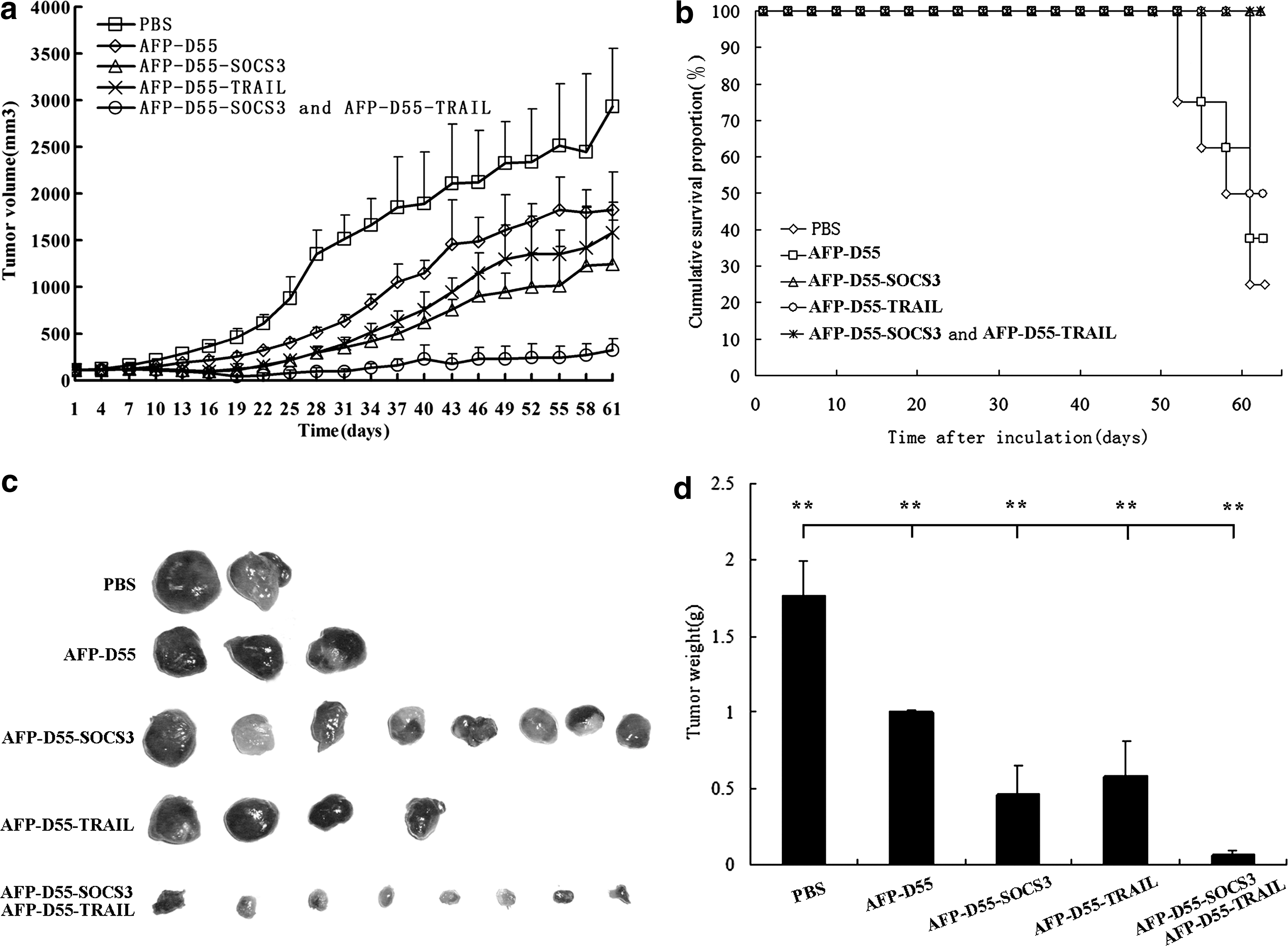
The antitumor effect of the combined treatment of AFP-D55-SOCS3 and AFP-D55-TRAIL in vivo.
As shown in Fig. 4b, a long-term survival rate of 100% in the combined treatment group was observed 61 days after inoculation through Kaplan-Meier survival analysis (100%), whereas the rates of the groups treated with AFP-D55-SOCS3 and AFP-D55-TRAIL, respectively, were 100% and 50%. The survival rate of the AFP-D55–treated group was 37.5% and that of the PBS-treated group was 25%. These results demonstrate the promising antitumor effect of the combined treatment of AFP-D55-SOCS3 and AFP-D55-TRAIL in vivo.
Overexpression of SOCS3 enhances TRAIL-induced apoptosis in HCC cell lines
To investigate whether the synergistic inhibition of cell proliferation was correlated with apoptosis, we analyzed apoptotic changes by Hoechst 33258 staining after treatment with AFP-D55, AFP-D55-SOCS3, AFP-D55-TRAIL, and a combination of AFP-D55-SOCS3 and AFP-D55-TRAIL for 48 hr in Huh7 cells. As shown in Fig. 5a, in Huh7 cells, apparent apoptosis was not observed with either AFP-D55-SOCS3 or AFP-D55-TRAIL, nor with the PBS and AFP-D55 treatments. However, apoptosis was clearly observed in the combined treatment of AFP-D55-SOCS3 and AFP-D55-TRAIL.
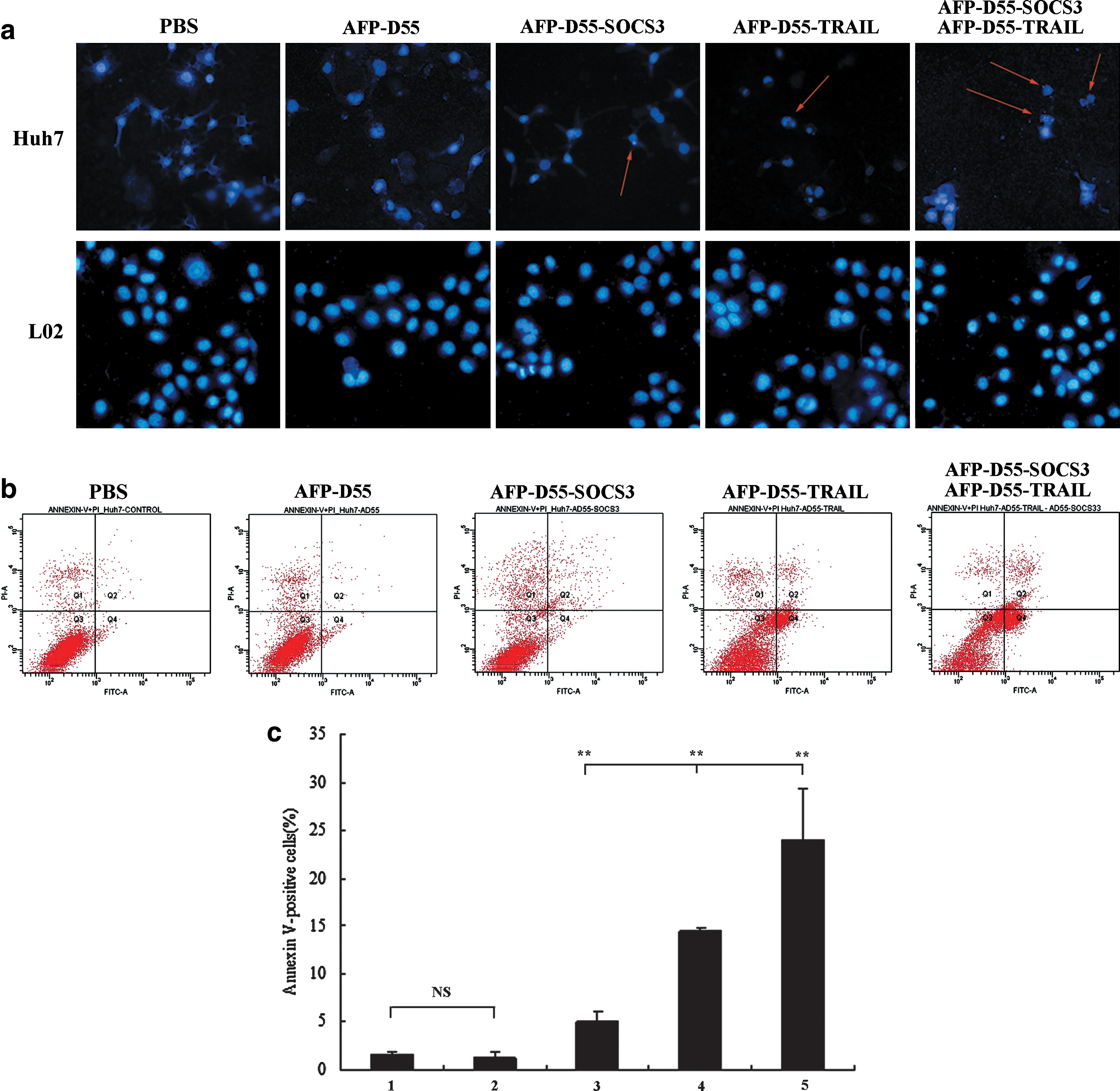
Overexpression of SOCS3 enhances TRAIL-induced apoptosis in HCC cell line.
To further analyze the induction of apoptosis by all the treatments, the percentage of apoptotic cell death was determined by FACS (Fig. 5b). The combined treatment of AFP-D55-SOCS3 and AFP-D55-TRAIL significantly increased the number of apoptotic cells to 24%. In contrast, the apoptotic percentages were 5.13% for AFP-D55-SOCS3, 14.47% for AFP-D55-TRAIL, 1.33% for AFP-D55, and 1.6% for PBS (Fig. 5c).
Western blot analysis was used to confirm the apoptosis induced by these treatments. Caspase-8, Caspase-3, and PARP cleavages were observed in all the treatments in the Huh7 cells. However, remarkable increases in the cleavages of Caspase-8, PARP, and Caspase-3, as well as an obvious decrease in Pro-caspase-9, were found with the combined treatment compared with other treatments (Fig. 6a). Therefore, these findings suggest that the TRAIL-induced apoptosis could be enhanced by the overexpression of SOCS3 in vitro.

The protein dynamic changes in AFP-positive HCC cell lines (Huh7, HepG2, Hep3B, PLC) were investigated 72 hr after infection with different double-regulated oncolytic adenoviruses. Lane 1, PBS; Lane 2, AFP-D55; Lane 3, AFP-D55-TRAIL; Lane 4, AFP-D55-SOCS3; Lane 5, AFP-D55-SOCS3 and AFP-D55-TRAIL.
The mechanism responsible for the synergistic anti-tumor effects of AFP-D55-SOCS3 and AFP-D55-TRAIL
The protein dynamic changes in AFP-positive HCC lines (Huh7, HepG2, Hep3B, PLC) were investigated 72 hr after infection with different double-regulated oncolytic adenoviruses. The phosphorylated STAT3 protein, as well as Cyclin D1, decreased in the AFP-D55-SOCS3–treated group and in the jointly treated group of AFP-D55-SOCS3 and AFP-D55-TRAIL (Fig. 6b). These results suggest that restoration of SOCS3 by AFP-D55-SOCS3 could revoke constitutive activation of STAT3 and induce cell cycle arrest. Additionally, down-regulation of XAIP expression was detected in Huh7, Hep3B, and PLC cells (Fig. 6c). Down-regulation of Survivin was revealed in Huh7, Hep3B, and HepG2 cells, and down-regulation of Mcl-1 was found in all these cell lines. The down-regulation of these molecules coincided with the decrease of phosphorylated STAT3, which antagonized TRAIL resistance in these HCC cell lines. These data showed that SOCS3 not only can induce cell cycle arrest but also can antagonize TRAIL resistance of these HCC cell lines.
Seven days after the first injection, tumor specimens were obtained from tumor-bearing nude mice and subjected to histopathology, immunohistochemistry, and TUNEL staining. Hexon is a coat protein of adenovirus and could be detected in all groups treated with the double-regulated oncolytic adenovirus (Fig. 7). The expression of SOCS3 and TRAIL could be simultaneously detected in the same tumor sample obtained from the combination-treated group, whereas SOCS3 and TRAIL were only separately detectable in the AFP-D55-SOCS3–treated group or AFP-D55-TRAIL–treated group (Figs. 7 and Supplementary Fig. S1; Supplementary Data are available online at
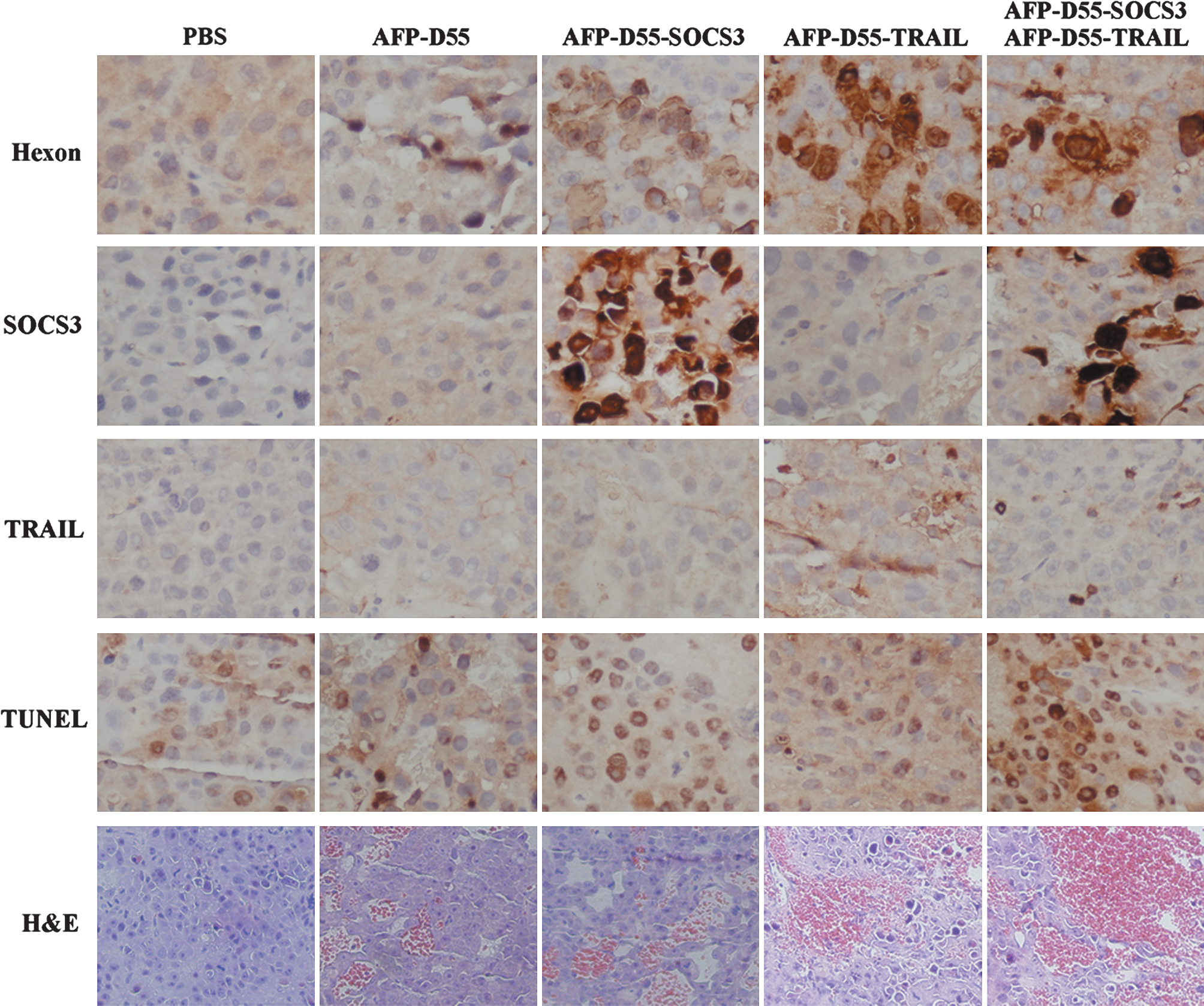
Histopathologic, immunohistochemical, and TUNEL analysis of tumor sections. Tumors or livers were harvested and fixed in 4% paraformaldehyde, embedded in paraffin, and cut in 4-μm sections. They were then subjected to immunochemical analysis for hexon, SOCS3 and TRAIL, TUNEL assay, and HE staining of tumor sections in different groups; the red area of HE staining indicates necrosis. Scale bar = 200 μm.
Discussion
In this study, a double-regulated oncolytic adenovirus vector that can replicate and induce CPEs in AFP-positive HCC cell lines with p53 dysfunction were successfully constructed. It has been suggested that overexpression of inhibitors of apoptosis (IAPs) is an important reason for tumor resistance to TRAIL. Increased levels of IAPs have been detected in most transformed and tumor cell lines (Ferreira et al., 2002; Tamm et al., 2000; Watanuki-Miyauchi et al., 2005). XIAP, a member of the IAP family, was observed to overexpress in several HCC cell lines (Huh7, BEL7404, Hep3B).
The double-regulated oncolytic adenoviruses AFP-D55, AFP-D55-SOCS3, and AFP-D55-TRAIL can specifically replicate in AFP-positive HCC cell lines. The AFP promoter was chosen because it can ensure which patients are suitable for drug administration of these double-regulated oncolytic adenoviruses, since AFP is a clinical diagnostic marker of AFP-positive HCCs. Specific replication and low progeny yield in AFP-positive HCC cell lines rendered these double-regulated oncolytic adenoviruses remarkable safe. Although these double-regulated oncolytic adenoviruses were low in progeny replication, leading to low cell oncolytic potency and low gene copy numbers in infected cells, cell growth was sufficient to drive the expression of these therapeutic genes to inhibit cell proliferation both in vitro and in vivo. We modified CTGVT by applying two therapeutic genes to create CTGVT-DG. The two genes may have compensative or synergetic effects; therefore, the CTGVT-DG strategy usually results in complete eradication of xenograft tumors in nude mice (Pei et al., 2004; Liu et al., 2005; Chu et al., 2006; Y.H. Zhang et al., 2006; Zhao et al., 2006; Z.W. Zhang et al., 2009).
The combined treatment of AFP-D55-SOCS3 and AFP-D55-TRAIL (2:3) exhibited potent antitumor activity in AFP-positive HCC cell lines compared to any other treatment both in vitro and in vivo in spite of low gene copy numbers. Notably, the synergistic inhibition of cell proliferation was correlated with cell apoptosis. The TRAIL-induced apoptosis could be enhanced by AFP-D55-SOCS3 both in vitro and in vivo. The necrotic areas of tumor sections in the combined treatment group were much larger than those in any other treatment groups. AFP-D55 had little or no inhibitory effect on cell proliferation either in vitro or in vivo in AFP-positive HCC cell lines (Huh7, BEL7404, Hep 3B). So the necrosis may not be due to the oncolysis of adenoviruses but rather to exogenous genes. This finding suggests that the combined treatment of AFP-D55-SOCS3 and AFP-D55-TRAIL can dramatically induce tumor necrosis in the tumors.
The decrease of the phosphorylated STAT3 protein in both the AFP-D55-SOCS3–treated and combined treatment groups, and down-regulation of anti-apoptotic proteins such as XIAP, Survivin, Bcl-xL, and Mcl-1 coincidently detected with the decrease of phosphorylated STAT3, indicated that the down-regulation of the anti-apoptotic proteins was actually correlated with the phosphorylated STAT3 protein decrease. This appeared to be a consequence of SOCS3 restoration, since SOCS3 sensitized HCC cells to TRAIL-induced activation of apoptosis.
In conclusion, our data demonstrate that restoration of SOCS3 by AFP-D55-SOCS3 (i.e., CTGVT) could not only antagonize HCC therapeutic resistance to TRAIL and adenoviruses, but also induce cell cycle arrest in HCC cell lines, which is the mechanism responsible for the synergistic inhibitory effects of AFP-D55-SOCS3 and AFP-D55-TRAIL. The combined treatment of AFP-D55-SOCS3 and AFP-D55-TRAIL may provide a useful strategy for the treatment of HCC with respect to enhancing antitumor potency and safety and overcoming drug resistance.
Footnotes
Acknowledgment
We thank Prof. Zheng-Xuan Shen for critical reading the manuscript and Ms. Lan-Ying Sun for help with the cell culture. This work was supported by the National Nature Science Foundation of China (No. 30623003), the National Basic Research Program of China (973 Program) (No. 2010CB529901), the National Basic Research Committee of Science and Technology (Nos. 06ZR14072 and 074119508), Important National Science & Technology Specific Project of Hepatitis and Hepatoma Related Program (2008ZX10002023).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.