
Abstract
Because of their high efficiency, virus-based vectors are currently used in most gene therapy trials. Because such vectors bear some potential safety risks, nonviral expression systems could be an attractive alternative. Ideally, these vectors should be completely based on chromosomal elements and replicate as an autonomous unit in the recipient cell, thus avoiding the risk of insertional mutagenesis or immunological reactions of the recipient organism. Our limited knowledge of the epigenetic regulation of replication in mammalian cells does not yet allow the rational design of such constructs. But in the late 1990s it was shown that scaffold/matrix attached region (S/MAR)-based vectors can promote episomal replication and maintenance in mammalian cells. These vectors have found broad application in basic research but are now improved for their use in the safe and reproducible genetic modification of cells and organisms and in gene therapy.
Introduction
ARSs were isolated from the yeast genome and found to promote episomal replication when inserted into a plasmid (Stinchcomb et al., 1980). ARSs are short (about 125 bp), AT-rich DNA sequences with a highly conserved 11-bp core sequence. They represent binding sites for the origin recognition complex (ORC) and about 500 of these sequences are used for replication of the yeast genome (Duncker et al., 2009).
When restriction-digested mammalian DNA was inserted into yeast plasmids numerous DNA sequences promoted autonomous replication in yeast. However, transfection of such vectors into mammalian cells never resulted in episomally replicating plasmids and in most cases the ultimate fate of such constructs was either loss from the cell or integration into the genome. This strongly argues that the mammalian genome contains sequences somehow resembling ARSs in yeast but not acting as replication origins in mammalian cells. Only in some rare cases insertion of putative mammalian origins of replication into plasmids resulted in episomal maintenance under selective conditions (Nielsen et al., 2000; Price et al., 2003). Subsequent sequence analyses of various mapped mammalian origins of replication revealed no sequence homologies but rather a number of structural characteristics, such as long AT-rich regions, CpG islands, bent DNA, and the presence of scaffold/matrix attached region (S/MAR) sequences (Depamphilis, 1999; Gilbert, 2004). It is therefore believed that selection and activation of mammalian origins is not only determined by the DNA sequence itself but also by epigenetic factors, such as chromatin structure, gene expression, and even global nuclear architecture (Mechali, 2010). Besides the insertion of putative mammalian origins of replication into plasmids, various other approaches were made to construct autonomous replicons for mammalian cells. These included the construction of mammalian artificial chromosomes or the insertion of chromosomal sequences into vectors encoding the viral protein Epstein–Barr nuclear antigen (EBNA)-1. Artificial chromosomes can be constructed either by the “bottom-down” approach, that is, the truncation of existing chromosomes, or by the “bottom-up” approach, in which centromeric, telomeric, and genomic sequences are assembled and delivered to the cell. Although such artificial chromosomes have in theory great potential both for basic and applied research, they are highly complex, often contain undefined chromosomal sequences, are often structurally unstable, and are technically difficult to construct, and their delivery to the target cell still remains a great challenge (Macnab and Whitehouse, 2009). When chromosomal sequences were introduced into EBNA-1-based vectors a number of DNA sequences supporting episomal replication in this context were identified (Stoll and Calos, 2002). It is assumed that nuclear retention of these vectors is achieved by EBNA-1, which is also possibly involved in the recruitment of the ORC. In fact, replication-promoting sequences were also isolated by chromatin immunoprecipitation of genomic chromatin with antibodies directed against ORC proteins and insertion of the precipitated DNA into EBNA-1-encoding vectors (Gerhardt et al., 2006; Schepers and Papior, 2010). But again, sequence comparison of these replication-promoting sequences revealed no common sequence elements. Although this approach may contribute to our understanding of replication control in mammalian cells, there are concerns about safety problems of EBNA-1-encoding vectors when used for gene therapy because it has been shown that this protein may cause cellular transformation in lymphocytes (Wang and Sugden, 2005).
When constructing episomal vectors based exclusively on chromosomal elements, epigenetic factors that are possibly involved in the selection and activation of replication origins must be considered. DNA replication takes place in so-called DNA foci that contain clusters of active origins that are somehow associated with the nucleoskeleton, probably by an interaction with S/MARs. Already during G1 of the cell cycle the ORC loads MCM2-7 and thus generates clusters of potential origins that are licensed for activation. During the S phase cyclin/Cdk complexes are essential for activation and assembly of replication complexes at replication origins. Different regions of the genome are replicated at different time points of the S phase. This timing strongly depends on the transcriptional and chromatin status of the respective region and follows a temporal and spatial program that is dictated by chromosome structure and nuclear localization (Jackson and Pombo, 1998; Blow and Dutta, 2005; Joffe et al., 2010). Our knowledge of these processes is too limited to allow the rational design of vectors behaving as endogenous replication origins. However, it is believed that origins bind to a subnuclear structure, the nuclear matrix or skeleton, at the onset of the S phase by an interaction of S/MAR sequences, which are often observed to be associated with mapped origins of replication (Cook, 1999; Rivera-Mulia et al., 2011). As described for mammalian origins of replication, S/MAR sequences do not share high sequence homologies but are rather characterized by their structural features. Often they contain stretches with more than 70% AT-rich sequences, DNA-unwinding elements, and show binding sites for transcription factors and topoisomerase II. S/MARs play a role in targeting gene domains to matrix-associated transcription foci and have been shown to support long-term gene expression from ectopic chromosomal sites (Alvarez et al., 2000; Bode et al., 2000; Zahn-Zabal et al., 2001). The insertion of a strong S/MAR into a plasmid led to the construction of the first nonviral episomal vector for mammalian cells, which proved to be a minimal model system for the analysis of replication control but also has great potential as an alternative nonviral vector system for gene therapy.
pEPI: The First Generation of S/MAR Vectors
S/MAR sequences have been assigned a number of important functions in the orchestration of gene expression and replication (Bode et al., 2000; Chattopadhyay and Pavithra, 2007). Moreover, characterization of mapped mammalian origins of replication revealed that S/MAR sequences are frequently found in close proximity to replication origins (Depamphilis, 1997; Cayrou et al., 2010). The abilities of S/MARS to support the episomal replication of plasmids were assayed in the late 1990s. When putative mammalian origins of replication were cloned into S/MAR-containing yeast artificial chromosome (YAC) vectors, episomal vectors were retained under selective conditions in some cases and evidence was provided that at least one of these constructs binds to the nuclear matrix (Cossons et al., 1997; Nielsen et al., 2000). But the first vector shown to replicate autonomously in a variety of cell lines and retained in the absence of selection was the vector pEPI-1 (Piechaczek et al., 1999). In this construct the sequence encoding the SV40 large T-antigen was replaced by a strong S/MAR sequence derived from the human interferon (IFN)-β gene cluster. A restriction map of the resulting pEPI-1 vector is shown in Fig. 1a. When transfected into Chinese hamster ovary (CHO) cells pEPI-1 replicated as a low copy number episome with 5–10 copies per cell (Stehle et al., 2007) and was stably retained in the absence of selection for basically unlimited time (Piechaczek et al., 1999). Episomal replication of this vector was not restricted to CHO cells but was observed in all cell lines tested, including primary human cells. In all cases it was mitotically stable in the absence of selection and occurred at an average copy number less than 10 per cell (Fig. 1b) (Jackson et al., 2006). This implies that replication and segregation to daughter cells must be highly efficient.
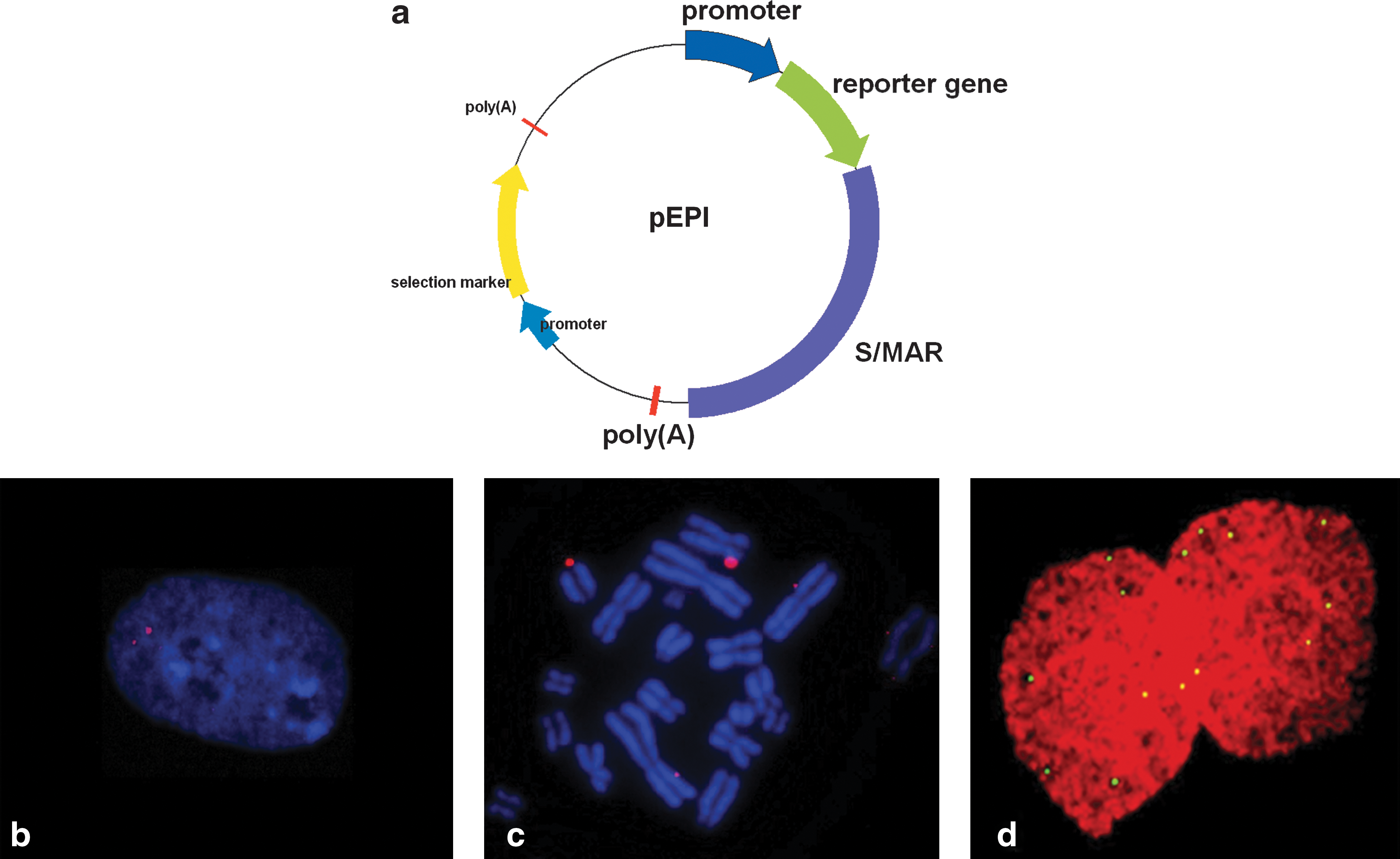
The episomal vector pEPI. (
Nuclear fractionation procedures revealed that the vector binds to the nuclear matrix, and specifically to the prominent matrix protein SAF-A (scaffold attachment factor A) (Jenke et al., 2004). Fluorescence in situ hybridization (FISH) analyses showed that pEPI-1 cosegregates with the host chromosomes during mitosis (Fig. 1c and d). However, no preferential binding sites to mitotic chromosomes could be observed, although it cannot be excluded that they always associate with specific sequence elements (Baiker et al., 2000; Stehle et al., 2007). The vector replicates once early during S phase and the origin recognition complex can assemble at various regions of the chromosome (Schaarschmidt et al., 2004), thus behaving as the initiation zone of genomic origins of replication (Depamphilis, 1999).
Although the vector pEPI carries all cis-acting sequences required to support episomal replication and maintenance, only in a small percentage (less than 5% as estimated by colony-forming assays) of transfected cells will the vector establish stably as an episome. This implies that in addition to the DNA sequence, epigenetic factors play a crucial role in establishing it as an autonomous replicon, a phenomenon not restricted to nonviral replicons but also reported for Epstein–Barr virus (EBV)-based vectors (Leight and Sugden, 2001a,b). Establishment and behavior of pEPI vectors were monitored by various microscopy techniques. Although many vector molecules reach the nucleus 24 hr posttransfection, the majority of vector molecules are subsequently lost from the cell. In cells containing a stably established replicon it was found exclusively in less condensed chromatin (Fig. 1b) regions (the interchromatin space) and frequently associated with nuclear speckles, that is, nuclear regions involved in RNA processing. The vector associates with early replicating foci and this association is stably retained over mitosis, explaining the high mitotic stability of this construct. Once established, the vector seems to be surprisingly nondynamic throughout the cell cycle (Stehle et al., 2007; Tessadori et al., 2010) and is associated with histone modifications typical of active chromatin, with an accumulation of histone 3 methylated at Lys-4 (H3K4me1,me3) in the S/MAR element (Rupprecht and Lipps, 2009). The pattern of all histone modifications remained constant during G1, S, and G2 phases, but during mitosis H3K4me1,me3 is selectively removed from the S/MAR. These modifications are typical of active chromatin and enriched on genes replicating early during S phase (Koch et al., 2007; Hiratani and Gilbert, 2009). Their accumulation on the S/MAR sequence therefore highlights the relevance of this sequence element for vector functioning. According to these data, vector establishment is a stochastic event and only those vector molecules reaching a nuclear compartment favorable for transcription and replication will assemble into an appropriate chromatin structure and be retained as an episome. Silencing the expression cassette linked to the S/MAR results in loss of the episome from the cell (Rupprecht et al., 2010). This allows the construction of vectors that can be selectively removed from the cell once expression of the transgene is no longer required in the recipient cell. Interestingly, only a certain percentage of cells (between about 30 and 70%) containing pEPI as an episome express the reporter gene encoding enhanced green fluorescent protein (eGFP) to a degree that can be detected by fluorescence-activated cell-sorting (FACS) analysis, indicating that minimal transcriptional activity is sufficient to maintain the episome. It is possible that the expression level again depends on the nuclear localization state of the vector after transfection and consequently the chromatin structure it adopts. However, the expression profile of a given cell clone will stay constant through many cell generations. Future improvements of this prototype vector therefore should aim to increase the expression level of the transgene and to improve establishment efficiency by identification and insertion of possible cis-acting sequences, such as, for example, binding sites for specific nuclear proteins that direct a DNA sequence into an active nuclear compartment.
The Next Vector Generations
Major limitations of the prototype pEPI vector are its variable expression in various cell lines, including the problem of transgene silencing and its low establishment rate. Therefore most modifications that have been made focus on improved transgene expression, increased establishment efficiency, and its application in animal disease models. But despite the respective modifications distinct features of pEPI cannot be changed to maintain its episomal replication and maintenance. By designing several deletion constructs Stehle and colleagues (2003) demonstrated that for episomal maintenance transcription must run into the S/MAR linked to an expression cassette. Whenever transcription of the transgene running into the S/MAR sequence was abrogated, that is, by deletion of the transgene, deletion of the promoter, or insertion of a termination signal between the transgene and S/MAR, the plasmids either integrated into the host genome or became lost from the cells. This was further demonstrated by the construction of an inducible vector. The transgene-S/MAR cassette was placed under the control of a tetracycline-responsive promoter (TetON) and stably transfected cells were established in the presence of doxycycline. Removing doxycycline after establishment resulted in a continuous loss of vector molecules from the cells. It was shown that this inducible vector system also functions in vivo. Hydrodynamic injection of pEPI-TetON into mice resulted in a 5-fold increase in transgene expression when doxycycline was administered to mice, even though the system is leaky and background expression was detectable in the absence of doxycycline (Rupprecht et al., 2010).
The S/MAR sequence present in the original vector has a size of approximately 2 kb. It was shown that replacement of the original 2-kb S/MAR by a tetramer of a 155-bp module comprising the core unwinding element of the human IFN upstream S/MAR (Bode et al., 1992) is sufficient for replication and episomal maintenance. In the same study the simian virus 40 (SV40) origin of replication was depleted and the resulting pEPI derivate pMARS represents a synthetic episomal vector that in terms of replicating and transcriptional behavior does not significantly differ from pEPI-1 (Jenke et al., 2004).
Because the expression cassette-S/MAR module is essential for the episomal behavior of S/MAR-based vectors, modifications of the pEPI-1 prototype vector apply only to the plasmid backbone, insertion of various promoters, and any other additional sequences without interfering with the promoter-transgene-S/MAR cassette. With the intention of applying pEPI in gene therapy, several attempts were undertaken to prevent silencing events and stimulatory effects on the innate immune system. The prototype pEPI-1 satisfies many of the requirements of an ideal vector for gene therapeutic approaches: it has been employed in several cell types (Papapetrou et al., 2006) and also for animal transgenesis (Manzini et al., 2006), it does not integrate into the host genome (Stehle et al., 2003), and contains no viral elements (Piechaczek et al., 1999). But the requirement for sustained long-term expression and persistence in every cell type has still not been achieved with this construct. Argyros and colleagues (2008) reported in a detailed analysis in vivo that the decrease in pEPI-based transgene expression over time is due to cytosine methylation of the cytomegalovirus (CMV) promoter, a phenomenon also observed for other vector constructs (Argyros et al., 2008). For this reason various other promoters were introduced into pEPI-1. The CAG promoter, for example, represents a synthetic hybrid promoter consisting of a CMV enhancer element, chicken β-actin promoter sequences, and rabbit β-globin 3′-UTR (untranslated region) sequences and was first used by Niwa and colleagues (1991). The CAG promoter is less prone to cytosine methylation and stably active in most tissues in vivo (Sawicki et al., 1998). Manzini and colleagues substituted CMV with CAG, resulting in pEPIX. Compared with pEPI-1 (∼25% eGFP-expressing cells), pEPIX displayed elevated eGFP expression levels (∼55% eGFP-expressing cells) that were stable over more than 40 generations in culture (Manzini et al., 2010). Haase and colleagues (2010) replaced the CMV promoter of pEPI-1 by a combination of CMV enhancer element and elongation factor-1 promoter (hCMV/EF1P) and achieved significantly increased and prolonged transgene expression in vitro and in vivo. Furthermore, in the same study they reduced the CpG content of the pEPI backbone, resulting in an episomal plasmid named pEPito. pEPito is not only reduced in size but also displays increased establishment in vitro, up to 6-fold over pEPI-1, as well as increased and stable transgene expression in vivo, up to 32 days after hydrodynamic injection into MF-1 mice (Haase et al., 2010). Whereas pEPI and its improved derivates work successfully in vitro, long-term gene expression in vivo has been shown to be more complex, that is, pEPI and some of these derivatives became silenced quickly when delivered into mice (Argyros et al., 2008). To overcome this limitation, one crucial first step is to replace the CMV promoter with respective tissue-specific promoters. This issue is discussed in detail in Applications (see below).
Increased and prolonged transgene expression in vitro and in vivo has also been reported for “minicircles,” which lack any residual elements for bacterial propagation (Darquet et al., 1999) and may therefore avoid immune responses of the host organism. It was shown that minicircles support enhanced transgene expression and are more stable in the recipient cell and organism than plasmids containing bacterial sequences. Originally, all methods to produce minicircles were based on the use of recombinases such as λ integrase (Darquet et al., 1999), Cre recombinase (Bigger et al., 2001; Vaysse et al., 2006), Flp recombinase (Nehlson et al., 2006), and φC31 recombinase (Chen et al., 2003; Jacobs et al., 2008); now, however, they are also synthetically produced and commercially available (Mayrhofer et al., 2008) (
Interestingly, all improved pEPI derivates and minicircles occur in the same copy number as the original pEPI-1 in established cells, suggesting stringent copy number control in the recipient cells. Given this narrow range of copy numbers per cell and the wide range of expression levels, epigenetic features, such as chromatin structure and nuclear localization, seem to have an important role in the regulation of transcription from nonviral episomes.
Applications
In vitro
As described above, successful long-term transgene expression in vitro from pEPI and its derivatives has been demonstrated in several cell lines. For example, pEPI was shown to be episomally maintained under nonselective conditions in primary human fibroblast-like cells and at low levels even in human CD34+ cells (Papapetrou et al., 2006). In another approach, pEPI was used to express a short hairpin RNA (shRNA) cassette directed against the fusion region of the bcr-abl gene in K562 cells. It remained as an episome for 4 months in the absence of selection and expression of the Bcr-Abl fusion protein was greatly reduced (Jenke et al., 2005). In a similar approach, an shRNA expression cassette was cloned into pEPI that suppresses hepatitis B virus (HBV) replication in HepG2-2.15 cells. Eight months posttransfection total intracellular HBV DNA was reduced by 77%, showing remarkable suppression of HBV DNA replication (Jenke et al., 2008). Importantly, pEPI was shown to have a basically unlimited cloning capacity. When used to construct an iBAC-S/MAR vector encoding the 135-kb human low-density lipoprotein receptor (LDLR) genomic DNA locus, it was demonstrated that despite its size iBAC-S/MAR-LDLR remained episomal for 11 weeks and completely restored LDLR function in CHO ldlr–/– a7 cells to physiological levels (Lufino et al., 2007). In a similar study, the erythroid β-globin locus control region-β-globin gene (βLCR-HBB) microlocus cassette was combined with an S/MAR. These βLCR-HBB-S/MAR constructs were able to stably express β-globin at full physiological levels (Sgourou et al., 2009). Thus, a complete genomic locus including all its regulatory elements can be inserted into S/MAR-based vectors, ensuring natural and cell-specific expression of the therapeutic gene.
In vivo
The first in vivo study with pEPI involved its use in the generation of genetically modified pigs. The vector was delivered to female pigs by sperm-mediated gene transfer (SMGT) and expression of the transgene could be demonstrated in 9 of 12 modified fetuses. In these transgene-positive animals, expression was shown in all tissues, with up to 79% positive cells (Manzini et al., 2006). This high efficiency in the generation of genetically modified animals suggests that the vector acquires all epigenetic features in the egg cell for efficient establishment and propagation in the organism. In contrast, the situation seems to be more complex when the vector is delivered into differentiated tissues.
Even though pEPI has been successfully used in in vitro studies, long-term expression in vivo has been proven to be more challenging. As outlined above, one main limiting factor of pEPI is the silencing of the transgene by cytosine methylation of the CMV promoter within 1 week of delivery to the murine liver (Argyros et al., 2008) and lung (Conese et al., 2004). One crucial step to overcome this problem was the introduction of the liver-specific AAT promoter into pEPI, which resulted in long-term transgene expression in mouse liver for at least 6 months (Fig. 2a) (Argyros et al., 2008). However, after a 70% partial hepatectomy neither the CMV- nor the AAT-based vector demonstrated episomal replication, indicating that further improvements are necessary. It is probable that the establishment efficiency of the vector in the regenerating liver is too low to detect such rare events. To overcome this problem, Wong and colleagues (2011a) developed an AAT-S/MAR-based vector that provides a survival advantage to transfected cells over untransfected cells in vivo. The AAT-S/MAR vector was constructed to encode the bcl-2 gene, which inhibits activation of the Fas apoptotic pathway in the liver. In hepatocytes cell death can be specifically induced by binding of the physiological ligand FasL to the liver transmembrane protein Fas. Administration of the agonistic Fas antibody Jo2 activates this pathway in vivo, whereas Bcl-2 overcomes this pathway. The Bcl-2-S/MAR plasmid was delivered to mouse liver by hydrodynamic injection and Jo2 was administered twice per week. Indeed, despite constant Jo2 challenges luciferase expression was detectable for more than 3 months, indicating the maintenance of Bcl-2 expression over time (Wong et al., 2011a).
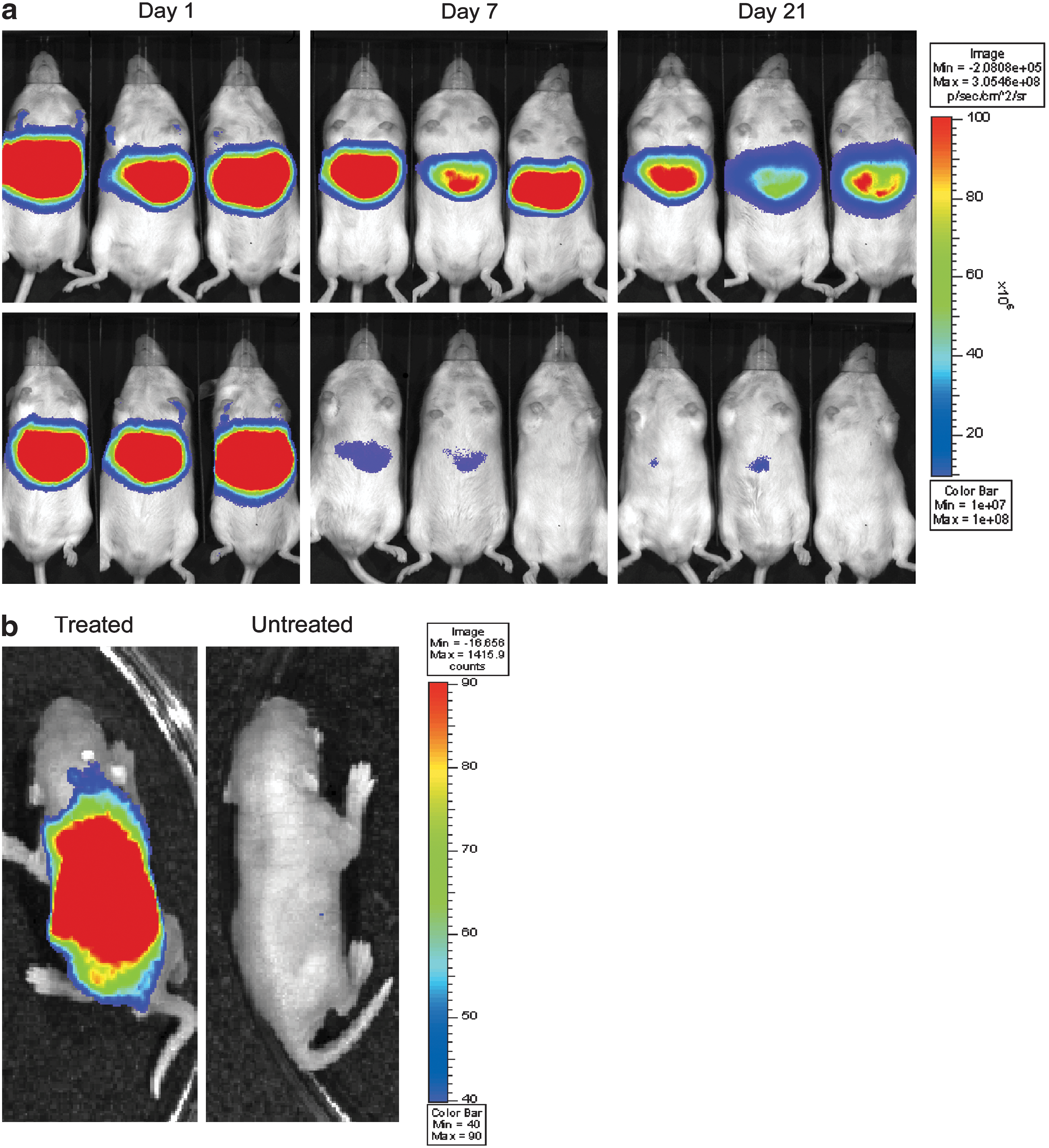
(
One of the first preclinical S/MAR-based studies (Cim et al., 2010) was based on an observation by Ferber and colleagues (2000), who reported that adenoviral delivery of the pancreatic transcription factor PDX1 (pancreatic and duodenal homeobox-1) mediated ectopic expression of insulin in mouse liver and therefore reverses streptozotocin-mediated hyperglycemia (Ferber et al., 2000). Accordingly, Cim and colleagues (2010) evaluated the level and duration of expression of the rat PDX1 transcription factor encoded on various nonviral vectors. After hydrodynamic delivery into rats, the original pEPI vector gave strong PDX1 expression on day 1 but showed a 150-fold decrease in expression on day 3. Vectors in which the CMV promoter was replaced with the AAT promoter gave sustained expression up to day 3 and a more gradual decrease afterward. However, a CpG-depleted S/MAR-based vector used in this study displayed both high levels and sustained PDX1 expression. Moreover, only this vector induced a pancreatic phenotype in rat liver. Two of three rats showed 30- to 70-fold increased insulin-2 expression on day 14 compared with day 1 (Cim et al., 2010).
A novel study reported the delivery of S/MAR-based vectors into neonatal mice. Nonviral vectors were administered via the superior temporal vein of mice at 1.5 days of age. Using a complex of polyethylenimine (PEI) and UbC-S/MAR pDNA, efficient transfection of the liver, lung, heart, spleen, brain, and kidney was achieved 24 hr postdelivery (Fig. 2b). A gradual increase in whole body luciferase expression was observed with peak expression on days 11 and 12 before reduction to background levels by day 25. On replacing the UbC promoter with the liver-specific AAT promoter, tissue-specific expression was shown in the mid-abdominal region, consistent with hepatocyte transfection during early mouse development. A similar transgene expression profile was observed with the liver-specific AAT promoter, where peak luciferase expression was detected on days 11 and 12 but fell thereafter, coinciding with the rapid proliferation of hepatocytes and reaching background level by day 20 (Wong et al., 2011b). The loss of expression was demonstrated to be due to the loss of vector genomes during the steep increase in liver weight, as similarly observed after the loss of vector genomes during partial hepatectomies in adult animals (Argyros et al., 2008). Nevertheless, use of this technique with optimized S/MAR vectors or gene delivery reagents is likely to be highly beneficial in future gene therapy strategies in early life.
Other studies have also investigated the application of S/MAR vectors in the adult central nervous system, muscle, as well as tumors. Studies of the CNS have used mainly plasmids expressing the gene of interest flanked by two S/MAR elements (Hughes et al., 2009; Kaytor et al., 2009; Yurek et al., 2009). Of interest is the study by Yurek and colleagues (2009), who investigated the delivery of a plasmid DNA expressing rat glial cell line-derived neurotrophic factor (GDNF) driven by a ubiquitin C promoter, flanked by two S/MAR elements. Injection into the rat left striatum resulted in stable overexpression of GDNF nanoparticles to levels 400–600% above normal expression 3 weeks postdelivery (Yurek et al., 2009). In another brain study, Kaytor and colleagues (2009) showed that another ubiquitin C promoter-driven luciferase plasmid with two S/MAR flanking domains was able to achieve reporter gene expression in the striatum for up to 1 year after a single intrathecal injection when complexed to polyethylene glycol (PEG)-substituted lysine 30-mer peptides (Kaytor et al., 2009). Studies of muscle have successfully shown strong levels of luciferase expression up to 3 months after delivery of pEPI-Luc, using microbubble-mediated ultrasound (Li et al., 2009). Significantly, the control plasmid not harboring the S/MAR returned to baseline levels in less than 1 month, as previously reported (Argyros et al., 2008).
Another potential area for the application S/MAR vectors is in cancer research. Delivery of plasmid pEPI-Luc into radiation
In terms of safety, nonviral vectors have a clear advantage over viral vectors. In particular, gene delivery using episomally maintained vectors is preferred over integrating vectors, which have the potential for insertional mutagenesis. S/MAR vectors are based solely on mammalian chromosomal elements and have the advantages of being able to provide extrachromosomal maintenance and long-term expression, as well as preventing epigenetic silencing. In addition, S/MAR vectors have a high insert capacity allowing simple modification, making it an ideal tool for gene therapy applications. The development of S/MAR minicircles is an exciting step in nonviral research, making it possible to provide a consistently high, maintained level of gene expression over a significant period of time. In addition, advances in inducible vectors highlight the progress made in this field and underline their potential in future gene therapeutic applications. Additional improvements in delivery techniques and nonviral formulations will further increase the prospects of S/MAR vectors in future clinical applications.
Footnotes
Acknowledgments
Work in the Lipps laboratory was supported by the Deutsche Forschungsgemeinschaft (SPP 1230). Claudia Hagedorn is supported by the Peter and Traudl Engelhorn Foundation. Suet-Ping Wong and Richard Harbottle are funded by the Myrovlytis Trust.
Author Disclosure Statement
The authors declare no competing financial interests.