
Abstract
Cisplatin-centered chemotherapy is the first-line treatment for human ovarian cancer. However, chemoresistance remains a major obstacle to successful treatment. Evidence has indicated that signal transducer and activator of transcription-3 (STAT3) is a determinant of chemoresistance; it was related to tumor recurrence in a large number of solid malignancies. Unfortunately, none of the compounds currently developed to block STAT3 signaling has been considered a serious clinical candidate because of toxicity or limited bioavailability. In this study, we clarified the significance of STAT3 activation in chemoresistant ovarian cancer and assessed the suitability of a novel oncolytic adenovirus (M4) designed to specifically deplete STAT3 and reverse cisplatin resistance in ovarian cancer. We showed that aberrant expression and constitutive activation of STAT3 was instrumental in cisplatin resistance in ovarian cancer cell lines and in ovarian cancer tissue samples. The M4 adenovirus could specifically deplete constitutive and inducible STAT3 and phosphorylated STAT3 proteins in ovarian cancer cells. This significantly inhibited cell survival and enhanced cisplatin-induced apoptosis. In contrast, normal human umbilical vein endothelial cells and human ovarian surface epithelial cells appeared to be unaffected by M4 treatment. Furthermore, a combined cisplatin plus M4 therapy substantially eliminated populations enriched in tumor-initiating cells. In mice, systemic intraperitoneal administration of M4 significantly potentiated the antitumor effect of cisplatin. These results suggest that M4 has great potential as a therapy against cisplatin resistance in human ovarian cancer. Thus, it warrants further clinical investigation.
Introduction
The study of drug resistance and cell survival factors in tumor cells has resulted in the identification of a number of potential targets for anticancer therapies. These targets include cytokines involved in growth and survival, and the various proteins that comprise their signaling pathways (Alas and Bonavida, 2003). On the other hand, one generally accepted hypothesis about chemoresistance is that standard therapies have failed to target tumor-initiating cells or cancer stem cells. Thus, the current forefront of cancer research is the development of new, effective strategies based on modulating a transient response to overcome drug resistance (Arafa et al., 2009).
The signal transducer and activator of transcription-3 (STAT3) is one member of a family of transcription factors activated by many cytokines, growth factors, and nonreceptor tyrosine kinases (Darnell, 1997; Bromberg and Darnell, 2000). In normal cells, the STAT3 protein plays a central role in cytokine signaling. In addition, they modulate a variety of biological functions by activating the transcription of a diverse set of genes (Smithgall et al., 2000; Rawlings et al., 2004). Studies have also revealed that STAT3 plays an important role in maintaining pluripotency and survival in embryonic stem cells (Kidder et al., 2008; Hall et al., 2009). In these cells, activation of the STAT3 protein is a reversible and tightly controlled process that typically lasts for a limited duration (Yu and Jove, 2004; Groner et al., 2008). Conversely, the constitutive activation or dysregulation of STAT3 protein is detected at high frequency in a large number of human malignancies (Turkson and Jove, 2000; Buettner et al., 2002). Huang and colleagues (2000) have previously demonstrated that constitutively active STAT3 (pSTAT3) was present in ovarian cancer cell lines, but not in normal ovary surface epithelial cells. With gene expression profiling and comparative genomic hybridization, Meinhold-Heerlein and colleagues (2005) also demonstrated that high-grade ovarian cancers were characterized by the expression of genes associated with STAT3-induced transcription. Studies have also shown that pSTAT3 was correlated with aggressive clinical behavior in ovarian carcinoma specimens (Silver et al., 2004; Duan et al., 2006; Rosen et al., 2006). However, little information is available on whether changes occur in STAT3 expression in chemoresistant ovarian cancer, recurrent ovarian cancer, or during the natural history of the disease.
Previous studies have identified upstream pathways in drug resistance, including the epidermal growth factor receptor, Janus-activated kinases, Src family kinases, the phosphatidylinositol-3-kinase (PI3K)/Akt pathway, and interleukin (IL)-6. All these pathways were shown to be elevated in ovarian cancer, which led to the phosphorylation of critical tyrosine and serine residues that activated STAT3 dimerization. Subsequently, STAT3 dimers translocated to the nucleus and initiated transcription through specific binding to DNA response elements in the promoter regions of target genes (Savarese et al., 2002; Duan et al., 2006, 2009). Studies have shown that STAT3 prevented tumor cell cycle arrest and cell death through multiple mechanisms that were previously linked to drug resistance. These mechanisms included increased expression of the oncogenes encoding cyclin D and c-Myc, and the antiapoptotic proteins MCL-1 (myeloid cell leukemia-1), survivin, and Bcl-xL (Duan et al., 2006).
The functional properties of STAT3 make it a difficult target for cancer drug therapy. STAT3 does not exhibit enzymatic activities, and it does not have any ligands of low molecular weight, the two prerequisites for conventional drug target molecules. Moreover, upstream activating events can be highly diverse in different tumor cells; thus, it is nearly impossible to predict which kinase should be targeted to prevent activation of STAT3 (Groner et al., 2008). Nevertheless, one study described the development of some small-molecule compounds that could block STAT3 signaling and displayed potent antitumoral activity (Schust et al., 2006; Iwamaru et al., 2007; Xu et al., 2009). However, before the safety and therapeutic efficacy of these promising agents can be evaluated in clinical trials, some fundamental concerns remain to be addressed. Therefore, it is necessary to reassess, refine, and incorporate new technologies into current strategies for targeted STAT3 interference in order to create suitable STAT3-blocking agents that can be developed into clinically useful anticancer therapeutics (Yue and Turkson, 2009).
A novel approach that holds promise as a highly effective treatment is the combination of conditionally replicative oncolytic adenoviruses (CRAds) and chemotherapy (Hermiston, 2000; Mathis et al., 2005; Rein et al., 2006). Initial clinical studies with oncolytic viruses have produced encouraging results in efficacy with minimal or no toxicity (Kanerva and Hemminiki, 2005). We have previously described a novel oncolytic adenoviral mutant called M4, which showed potent antitumoral efficacy by silencing the expression of STAT3 and its downstream target genes and causing selective cytolysis; this treatment showed no obvious adverse effects after systemic administration (Han et al., 2009).
The current investigation had four primary aims. First, we examined the status of STAT3, both in chemoresistance and recurrent ovarian cancer, and determined whether chemotherapy affected STAT3 expression. Second, we investigated the effect of M4 on the constitutive expression and activation of STAT3 in drug-resistant ovarian cancer cells. Third, we evaluated whether M4 could overcome drug resistance or enhance drug sensitivity in cisplatin-resistant ovarian cancer cells in vitro and in vivo. Fourth, on the basis of previous findings for ovarian cancer stem cells (Zhang et al., 2008; Alvero et al., 2009), we sought to determine whether the combination of M4 and cisplatin could eliminate populations enriched in ovarian cancer stem cells.
Materials and Methods
Cell culture
The human ovarian cancer cell lines SKOV-3 and Caov-3 were purchased from the American Type Culture Collection (Manassas, VA). Cells were cultured in RPMI 1640 supplemented with 10% fetal bovine serum, penicillin (100 units/ml), and streptomycin (100 μg/ml). A cisplatin-sensitive ovarian cancer cell line (OV2008) and its resistant variant (C13K) were gifts from R. Goel (Ottawa Regional Cancer Center, Ottawa, ON, Canada). These cells were cultured as described previously (Yan et al., 2006). Human umbilical vein endothelial cells (HUVECs) were isolated as previously described (Han et al., 2009).
Clinical samples
Patients with ovarian cancer at Tongji Hospital (Wuhan, Hubei, China) provided informed consent for the collection of ascitic fluids and solid tumor specimens. The protocol was approved by the local ethics committee. All patients received primary treatment in our hospital. After initial debulking surgery and comprehensive staging surgery, patients harbored nearly no residual tumors, or they received standard platinum-combined adjuvant chemotherapy in at least six courses. Patients were considered clinically platinum resistant when they progressed during the initial platinum-based therapy, displayed stable disease as the best response to initial therapy, or relapsed after an initial response within 6 months of completion of platinum-based therapy (Yan et al., 2010).
Samples of ascitic fluid were collected by paracentesis for palliative purposes from six patients resistant to cisplatin. Cells from this fluid were prepared in primary cultures as previously described (Witham et al., 2007). Some single tumor cells were cultured under stem cell conditions (Zhang et al., 2008).
Eight tumor tissue samples were obtained from patients who underwent initial cytoreductive surgery. Five tumor tissue samples were obtained from patients with recurrent ovarian cancer during the second surgery after treatment with cisplatin-based chemotherapy. All tissue samples were divided into two parts; one part was snap frozen and stored at −80°C for subsequent Western blot analysis, and the other part was established in primary cell cultures, as previously described (Petronelli et al., 2009). Normal human ovarian surface epithelial cells (HOSECs) were isolated as previously described (Burke et al., 2001).
Adenoviral mutant
The adenovirus Ad5/dE1A was previously constructed by deleting amino acids 121–129 in the conserved region 2 (CR2) of early region 1A (E1A) (Han et al., 2009). We constructed the M4 mutant by removing the adenovirus death protein (ADP) open reading frame in the E3 region of Ad5/dE1A and inserting a fragment of STAT3 cDNA placed in the reverse (3′ to 5′) direction (bases 960–190). The vector control for M4 (Ad5/dE1A/ADP) had only the deletion of the ADP open reading frame. The viral mutants were constructed by homologous recombination in human embryonic kidney 293 (HEK-293) cells.
Immunohistochemistry
Tissue sections were immunostained with the LSAB (labeled streptavidin–biotin) system (Dako, Glostrup, Denmark) according to the manufacturer's instructions. Negative controls were processed in parallel by incubating them with rabbit IgG at the same dilution. For STAT3 expression levels, the staining intensity pattern was scored on the following scale: 0, no staining; 1, weak staining; 2, moderate staining; and 3, intense staining. The intensity of immunostaining was evaluated only when more than 50% of the area showed STAT3 expression. On the basis of the percentage of cells with positive nuclear staining, the pSTAT3 staining patterns were categorized into five groups: 1, no detectable immunostaining; 2, <25% of cells immunostained; 3, between 25 and 50% of cells immunostained; 4, between 50 and 75% of cells immunostained; 5, >75% of cells immunostained. The immunostaining intensity was evaluated by two independent observers that were blinded to clinical and STAT3 data.
Western blot
Whole-cell lysates were prepared from ovarian cancer cell lines, tumor tissue specimens, or primary ovarian cancer cells. Standard Western blot analysis was performed with anti-STAT3, anti-phospho-STAT3 (Tyr-705), anti-MCL-1, anti-Bcl-xL, or anti-survivin antibodies (Santa Cruz Biotechnology, Santa Cruz, CA). Equivalent protein sample loading was monitored with an anti-β-actin antibody. The probe proteins were detected with the enhanced chemiluminescence system (Pierce Biotechnology/Thermo Fisher Scientific, Rockford, IL).
Drug sensitivity assay
A 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay was used to analyze cell viability, as described previously (Yan et al., 2006). Briefly, cells were cultured overnight in 96-well plates (1×104 cells per well). M4 and/or cisplatin was added at the indicated concentrations for the indicated times. After addition of MTT for 1 hr, the number of surviving cells was assessed by determining the absorbance at 490 nm (A 490nm) of the dissolved formazan product, as described by the manufacturer (Promega, Madison, WI). Cell viability was expressed as the absorbance of the experimental group (A exp group) compared with the absorbance of the control group (A control), and was calculated as follows: A exp group/A control×100.
Apoptosis assay
Cancer cell apoptosis was measured by staining with fluorescein isothiocyanate-conjugated annexin V (annexin V–FITC) (green fluorescence) and propidium iodide (PI, red fluorescence; BD Biosciences, San Jose, CA). Briefly, harvested cells were washed twice in cold phosphate-buffered saline (PBS); then, 1×105 cells in 100 μl of PBS were transferred to a 5-ml culture tube. We added 5 μl of annexin V–FITC and 10 μl of PI to each tube. After 15 min, 400 μl of binding buffer was added to each tube. Fluorescence was quantified with a FACScan (BD Biosciences).
Evaluation of ovarian cancer cell line response to IL-6
To confirm IL-6 regulation of pSTAT3 protein expression, ovarian cancer cell lines were seeded and allowed to adhere overnight. The next night, the cells were serum-starved. The cells were then pretreated with exogenous recombinant human IL-6 (R&D Systems, Minneapolis, MN) at the indicated concentrations for 1 hr. Next, cells were treated with M4 and/or cisplatin, and their resistance to cisplatin was determined in the apoptosis assay. IL-6 was added to the culture every 2 days.
Colony formation assay
A 0.7% agar gel with 10% fetal bovine serum (FBS) in Dulbecco's modified Eagle's medium (DMEM) was added to a 6-well culture dish as a base agar. Exponentially growing C13K cells were infected with M4 at a multiplicity of infection (MOI) of 10 for 3 hr, followed by a 24-hr incubation with 20 μM cisplatin. Cells were then trypsinized and placed in a 0.4% agar gel with 10% FBS in DMEM that was poured on top of the base agar. Cells were allowed to grow for 10 days. Cell growth was expressed as the percentage of colonies formed compared with those formed by untreated cells. Results represent the averages from three independent experiments.
Tumor xenograft experiments
All animal manipulations were carried out in accordance with institutional guidelines under approved protocols. For the subcutaneous xenograft model, 36 female, 4- to 6-week-old, athymic nude mice received a subcutaneous injection of 5×106 Caov-3 cells via the flank. After tumor development, mice were assigned to six treatment groups of six mice each. The first group was treated with saline once daily for five consecutive days. The second group was treated with cisplatin (0.75 mg/kg/day) every 2 days for a total of four treatments. The third group was treated with 1×108 plaque-forming unit (PFU) doses of M4 once daily for five consecutive days. The fourth group was treated with 1×108 PFU doses of M4 once daily for five consecutive days, and 4 days after the first day of M4 treatment, they were given cisplatin (0.75 mg/kg/day) every 2 days for a total of four treatments. The fifth group was treated with 1×108 PFU doses of Ad5/dE1A/ADP once daily for five consecutive days. The sixth group was treated with 1×108 PFU doses of Ad5/dE1A/ADP once daily for five consecutive days, and cisplatin was given at the same times it was given to the fourth group. The M4 and Ad5/dE1A/ADP were injected directly into tumors, and cisplatin was injected into the abdominal cavity.
Tumors were measured weekly in two dimensions with calipers, and the volume was calculated as length×width2×0.52. The mice were killed 70 days after the initiation of treatment or when the tumor volume was larger than 1200 mm3. A parallel experiment was performed to determine STAT3 and pSTAT3 protein levels, and M4 replication (n=2 samples from each group).
For the intraperitoneal xenograft model, 1×107 Caov-3 cells were injected intraperitoneally into each of 72 female mice. Two weeks after inoculation, mice were assigned to six treatment groups, each with 12 mice. The first group was treated with saline once daily for five consecutive days. The second group was treated with cisplatin (0.75 mg/kg/day) every 2 days for a total of four treatments. The third group was treated with 2×108 PFU doses of M4 once daily for five consecutive days. The fourth group was treated with 2×108 PFU doses of M4 once daily for five consecutive days, and 4 days after the first day of M4 treatment, cisplatin (0.75 mg/kg/day) was given every 2 days for a total of four treatments. The fifth group was treated with 2×108 PFU doses of Ad5/dE1A/ADP once daily for five consecutive days. The sixth group was treated with 2×108 PFU doses of Ad5/dE1A/ADP once daily for five consecutive days, and cisplatin was given at the same times it was given to the fourth group. All of the agents were given intraperitoneally. One-half of the animals (n=36) were killed 4 weeks after starting treatment to assess the antitumor efficacy of drug treatment. We measured the size of the largest tumors, the volume of ascites, and the extent of dissemination. The volume was calculated as length×width2×0.52. In the remaining 36 animals, survival was calculated from the first day of treatment until death. Seventy days after the initial treatment, the experiment was terminated.
To examine toxicity, 16 mice without tumors were assigned to four treatment groups of four mice each. All treatments were delivered as intraperitoneal injections. The first group was treated with saline once daily for five consecutive days. The second group was treated with cisplatin (0.75 mg/kg/day) every 2 days for a total of four treatments. The third group was treated with 2×108 PFU doses of M4 once daily for five consecutive days. The fourth group was treated with 2×108 PFU doses of M4 once daily for five consecutive days, and 4 days after the first day of M4 treatment, cisplatin (0.75 mg/kg/day) was given every 2 days for a total of four treatments. Body weight was measured weekly until 6 weeks after initial treatment, when all mice were killed. Blood was collected from the abdominal aorta, and serum was prepared to assay the levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST).
Statistical analysis
All values represent means±SD of at least three independent experiments. The significance of differences between groups was assessed by two-tailed Student t test. Statistical significance was defined as p<0.05. Survival was analyzed by the Kaplan–Meier method and differences were analyzed by log-rank test.
Results
STAT3 expression and activation in cisplatin-resistant ovarian cancer cells
Platinum-based chemotherapy is the cornerstone treatment of epithelial ovarian cancer. Despite good initial response rates, the majority of patients ultimately relapse with resistant disease. Increased antiapoptotic activity is one of the major mechanisms of cisplatin resistance (Guo et al., 2009). On the basis of the critical role of STAT3 in cancer cell proliferation and survival, we compared the expression profiles of STAT3 and pSTAT3 in cisplatin-resistant ovarian cancer cells with those of normal, cisplatin-sensitive, HUVECs and HOSECs. Cisplatin-resistant cells included SKOV-3 cells, originally obtained from a patient with intrinsic resistance to clinically achievable doses of cisplatin; Caov-3 cells, reported to be resistant to cisplatin (Freeburg et al., 2009); and C13K cells, established by 13 monthly selections of cells that grew in the presence of cisplatin (Hirama et al., 2006). Western blot analysis showed that high levels of STAT3 and pSTAT3 were expressed in all of the ovarian cancer cell lines. In contrast, both HUVECs and HOSECs contained obvious STAT3, but not pSTAT3 (Fig. 1A). We next evaluated the expression of STAT3 and pSTAT3 in cisplatin-resistant primary ovarian cancer cells. We found high levels of STAT3 and pSTAT3 in all primary ovarian cancer cells from six patients with cisplatin resistance (CROC) and five with recurrent cancer (ROC) (Fig. 1B–D). There was a marked trend toward higher levels of STAT3 and pSTAT3 expression in recurrent tumors compared with matched primary tumors (Fig. 1E).

Analyses of STAT3 expression and activation in ovarian cancer cells.
Exposure to diverse chemotherapeutic agents can induce alterations in gene expression and in signaling cascades that can mediate resistance. Therefore, we treated primary ovarian cancer cell cultures (POC) with cisplatin to determine whether levels of STAT3 and pSTAT3 were affected by the treatment. We found that cisplatin treatment for 24 hr resulted in increased pSTAT3 in five of eight POC cultures (Fig. 1F), but the level of STAT3 protein was not appreciably altered. The other three POC cultures (POC1, POC4, and POC7) displayed high levels of pSTAT3 before cisplatin treatment (Fig. 1F), but the levels did not increase after treatment.
We next investigated cisplatin resistance in the POC cultures with high pSTAT3 levels compared with resistance in those with low pSTAT3 levels. After 24 hr of treating POC cultures with various concentrations of cisplatin, we tested cell viability. We found little change in the cell viability of POC4 and POC7 cultures treated with 10 μM cisplatin, and only about 20% of cells died when treated with 30 μM cisplatin (Fig. 1G). In contrast, cell viability of the other POC cultures decreased by 78–88% when treated with 30 μM cisplatin (Fig. 1G). Taken together, these results suggested that pharmacological inhibition of STAT3 may be a promising therapeutic strategy in the management of chemoresistance in ovarian cancer.
M4 inhibits STAT3 pathway activation specifically in ovarian cancer cells
We reasoned that, because STAT3 is persistently activated in ovarian cancers, transient or partial inhibition of STAT signaling might be sufficient to eliminate tumor cells. We assessed the effects of M4 on STAT3 and pSTAT3 protein expression in ovarian cancer cells. Infection of Caov-3 and SKOV-3 cells with M4 (MOI, 10) resulted in a substantial loss of both STAT3 and pSTAT3 proteins (Fig. 2A and B).

M4 specifically depleted STAT3, pSTAT3, and downstream targets in ovarian cancer cells, but not in normal cells. Caov-3 cells, SKOV-3 cells, HUVECs, and HOSECs were infected with various viral mutants at an MOI of 10 and cultured for the indicated times.
The benefits of cytotoxic chemotherapies are often attenuated by damage to normal cells. Unfortunately, dose reductions that limit side effects on unaffected tissues and organs can compromise the capability of cancer treatments to both reduce the tumor burden and eradicate metastatic disease. Thus, we determined whether M4 had adverse effects on normal cells.
Infection of HUVECs and HOSECs with M4, even at an MOI of 100, did not affect the levels of STAT3 protein, did not induce cleavage of poly(ADP-ribose) polymerase (PARP), and did not activate caspase-3 (Fig. 2C). Furthermore, to rule out adverse effects of M4 on normal cell growth, we examined the level of the endogenous cell cycle protein cyclin D1, which is a short-lived protein. We found that the level of cyclin D1 protein was not affected by M4, even at 96 hr postinfection (Fig. 2C). To investigate whether M4 would reduce viability or inhibit cell growth in normal cells, HUVECs and HOSECs were infected with M4 at an MOI of 100. After 96 hr, M4-treated cells showed cell viability and proliferation similar to those of untreated cells (Fig. 2D–F).
We then examined whether M4, which effectively blocked STAT3 and pSTAT3 expression, could decrease the expression of antiapoptotic proteins MCL-1, survivin, and Bcl-xL, which were previously linked to drug resistance. We found that M4 treatment caused a clear reduction in the level of these proteins in ovarian cancer cells (Fig. 2A and B). These results demonstrated that M4 had low toxicity and efficiently blocked STAT3 signaling in ovarian cancer cells.
M4 reverses cisplatin resistance in vitro
As described previously, high levels of STAT3 and pSTAT3 were expressed in cell lines and patients who were cisplatin resistant, and in patients with recurrent ovarian cancer that had been treated with cisplatin. Therefore, we next examined whether blocking the STAT3 signaling pathway could enhance cisplatin efficacy on ovarian cancer cells. First, we tested changes in cisplatin sensitivity by treating cells first with 10 MOI of M4 or Ad5/dE1A/ADP, and then, after a 3-hr infection period, with cisplatin for 48 hr. We found that the ability of cisplatin to inhibit cell proliferation and cell survival was significantly enhanced by M4 in Caov-3, SKOV-3, and C13K cells (Table 1). These data suggested that the M4-enhanced response to cisplatin was associated with STAT3 depletion in ovarian cancer cells.
Sensitization was defined as the ratio of the IC50 value for cisplatin to the IC50 value for M4 (10 MOI) combined with cisplatin.
M4 enhances cisplatin-induced apoptosis
The ability of cancer cells to respond to chemotherapeutic agents is believed to be due, in part, to their capacity for apoptosis. It is also generally accepted that drug-induced apoptosis is governed by both upregulation of proapoptotic factors or tumor suppressors and by modulation of cell survival factors. Deregulation of one or more of these factors may prevent drug-induced apoptosis (Fraser et al., 2003). As stated previously, constitutive STAT3 activation in ovarian cancer cells may contribute to their survival advantage, and M4 treatment could increase their cisplatin sensitivity. To investigate whether the combination of STAT3 blockade and chemotherapy would have synergistic effects, we analyzed the effect of M4 on cisplatin-induced apoptosis. Cisplatin-resistant C13K human ovarian cancer cells were treated with various doses of M4, cisplatin, or both. The combination of M4 and cisplatin was administered in two different ways: 48 hr of M4 followed by 24 hr of cisplatin, and vice versa. The proapoptotic effect of M4 infection on C13K cells was dependent on both concentration (MOI) and treatment time; 10 MOI of M4 alone could induce about 40% of apoptosis rate at 72 hr postinfection (Fig. 3A and B). C13K cells failed to respond to cisplatin at concentrations up to 30 μM. However, pretreatment with M4 at an MOI of 10 led to a remarkable increase in the apoptosis response to 10 μM cisplatin, and the effect was dose dependent (Fig. 3C and D). When 10 μM cisplatin was added before 10 MOI of M4, the apoptosis rate was only 31%. However, when 10 μM cisplatin was added after 10 MOI of M4, apoptosis increased to 62.3% (Fig. 3E). These results were further supported by a quantitative colony formation assay (Fig. 3F). Taken together, the results indicated that STAT3 expression was instrumental in the cisplatin resistance of cancer cells, and the combination of M4 and cisplatin elicited a significant cytotoxic response in cisplatin-resistant human ovarian cancer cells.

Synergism of M4 plus cisplatin is dependent on the drug administration sequence.
IL-6-stimulated pSTAT3 expression and cisplatin resistance are reversed by M4
IL-6 was elevated in the serum and peritoneal fluid collected from patients with ovarian cancer. High levels of IL-6 in body fluids were previously associated with a poor prognosis and resistance to chemotherapy (Wang et al., 2010). In several tumors, the IL-6/gp130/STAT3 signaling axis constitutes an autocrine and paracrine amplification loop (Rosell et al., 2009). To evaluate the role of IL-6 in regulation of pSTAT3 protein expression in ovarian cancer cells, cisplatin-sensitive OV2008 cells were treated with various concentrations of IL-6 (0–50 ng/ml) for 1 hr. The results showed that IL-6 induced pSTAT3 in a dose-dependent manner, but did not alter the expression of total STAT3 (Fig. 4A). To determine whether IL-6 affected the response to cisplatin, OV2008 cells were pretreated with IL-6 (0–50 ng/ml) for 1 hr, followed by 20 μM cisplatin treatment. The cells treated with cisplatin alone for 24 hr showed the characteristic morphology of cell death. In contrast, pretreatment with IL-6 rescued the cells from apoptosis caused by cisplatin (Fig. 4B). To further confirm these results, apoptotic levels were measured by flow cytometry. Pretreating the cells with IL-6 at 10 ng/ml caused a significant increase in the resistance to cisplatin, and pretreatment with IL-6 at 50 ng/ml inhibited cisplatin-induced apoptosis by about 50% (Fig. 4C). This indicated that the presence of exogenous IL-6 could increase the resistance of ovarian cancer cells to cisplatin treatment.
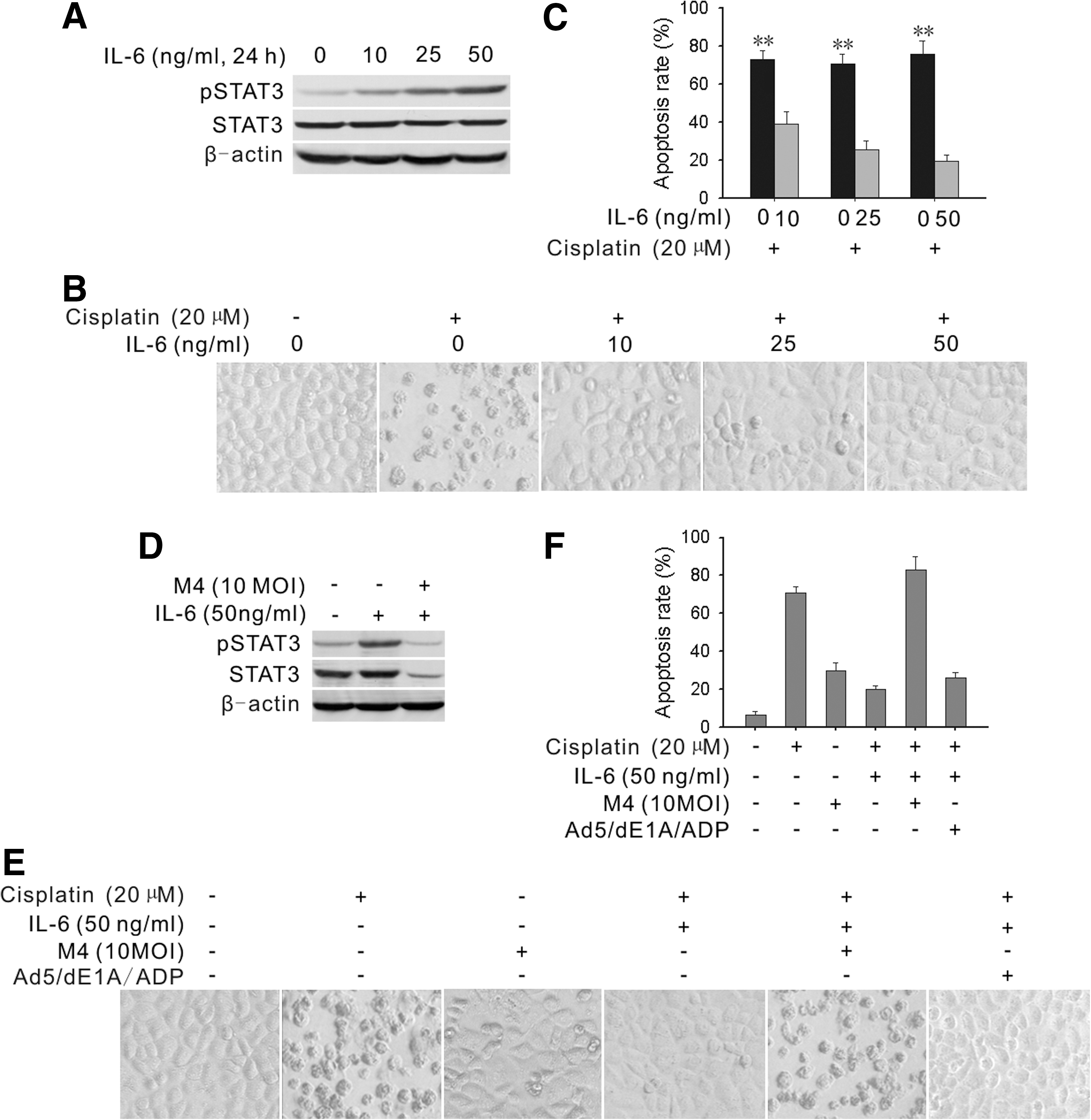
M4 blocked IL-6-induced pSTAT3 expression and counteracted IL-6-induced resistance to cisplatin-induced apoptosis.
To investigate whether M4 could counteract the effects of IL-6 on pSTAT3 activity, OV2008 cells were pretreated with IL-6 at 50 ng/ml, followed by infection with 10 MOI of M4 for 72 hr. A subsequent Western blot analysis (Fig. 4D) showed that M4 markedly reduced the levels of pSTAT3. We next determined whether M4 could counteract the IL-6-induced resistance of cells to cisplatin. Cells were pretreated with IL-6 at 50 ng/ml, and then treated with 10 MOI of M4 for 48 hr, followed by incubation with cisplatin (20 μM) for 24 hr. We found that the cells showed more cell death with M4 compared with without M4 treatment (Fig. 4E). The result was quantified in a cell apoptosis assay (Fig. 4F). Taken together, these results showed that M4 could completely block IL-6-induced cisplatin resistance.
M4 sensitizes ovarian cancer cells to cisplatin without affecting normal cell viability
To determine whether the M4 effects observed in ovarian cancer cell lines were applicable to primary ovarian cancer cells, we established primary cultures of cells collected from ascites fluid that had been drained from six patients resistant to cisplatin. All six cultures grew with similar morphology and growth rates. More than 95% of these cells expressed cytokeratin or epithelial antigen and displayed similar cobblestone morphology in culture (Fig. 5A). To determine the infectivity of Ad5 on primary ovarian cancer cells, the cells were infected with an adenoviral vector carrying the green fluorescent protein gene (Ad5-GFP) at an MOI of 10. Infectivity was defined as the percentage of cells that expressed GFP 48 hr postinfection, as assessed by flow cytometry. The results showed that as least 85% of cells could be infected with Ad5-GFP (Fig. 5B and C). We then explored whether M4 would sensitize these cisplatin-resistant primary ovarian cancer cells to cisplatin-induced cytotoxicity. We found that four of six of these cultures displayed at least a 3-fold increase in sensitivity to cisplatin after cotreatment with M4. Two cultures showed no significant change in sensitivity (cultures from patients 4 and 6) (Fig. 5D). This suggested that M4 could increase the sensitivity of primary cancer cells to cisplatin. However, a larger study will be necessary to quantify the proportion of cultures that respond to M4.
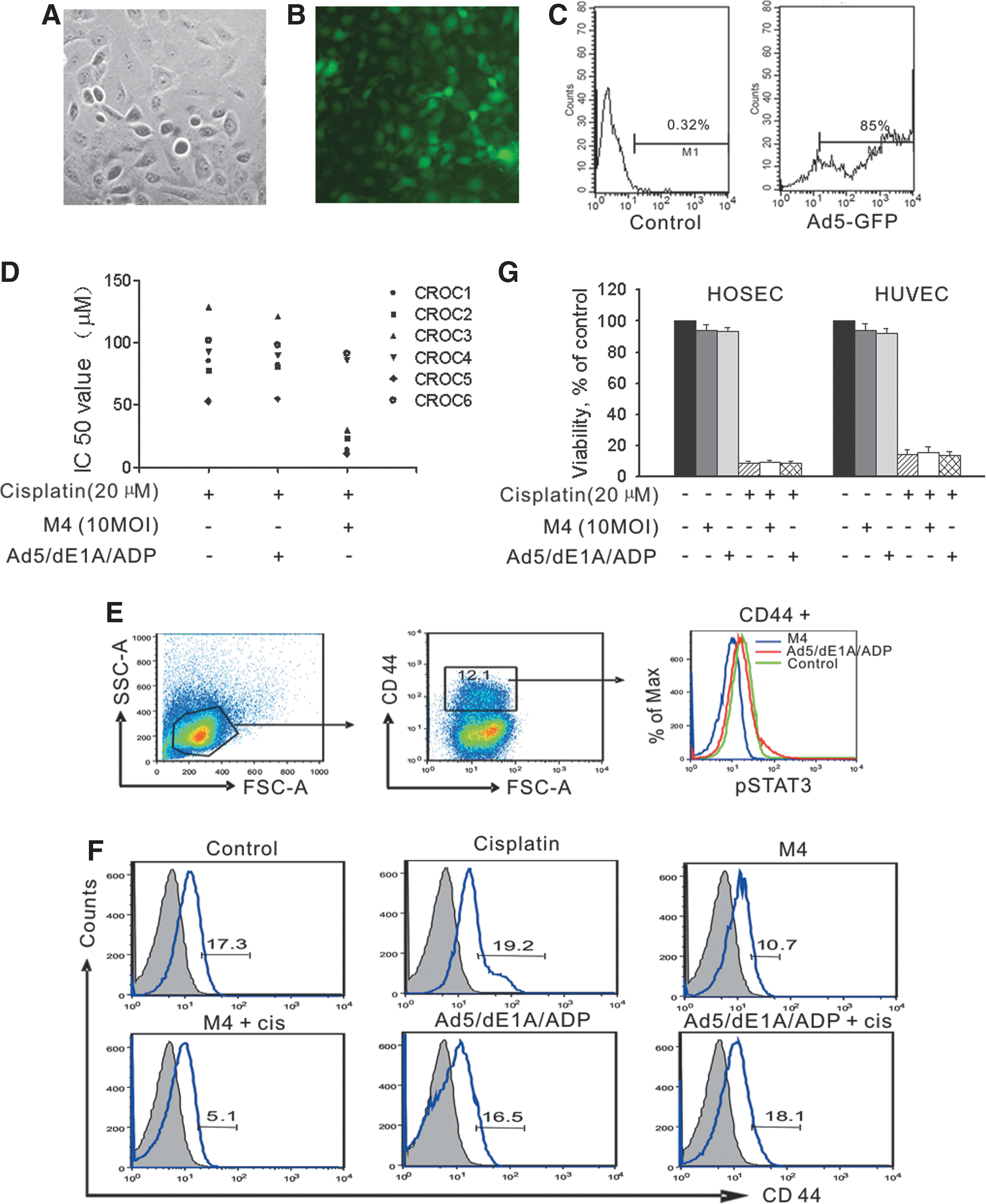
Primary ovarian cancer cells and ovarian cancer cells with stem-like properties treated with M4 were sensitized to cisplatin, but M4 did not affect cell viability of human normal cells.
Studies have identified the CD44 protein as a potential marker for ovarian cancer stem cells (Zhang et al., 2008; Alvero et al., 2009). We determined the percentage of CD44+ cells in cultures of cells from the ascites samples. Flow cytometric analysis showed that CD44+ fractions varied from 11 to 19% in the six cultures (Fig. 5E and F). We next assessed the effect of M4 on the level of pSTAT3 in the CD44+ cells. We found that, similar to findings obtained in ovarian cancer cell lines, the level of pSTAT3 in CD44+ cells was significantly decreased after 72 hr of M4 treatment at 10 MOI. Importantly, in three of six cultures, M4 treatment alone decreased the percentage of CD44+ cells, and the combination of M4 and cisplatin led to a more substantial decrease in the percentage of CD44+ cells (Fig. 5F). These results indicated that pSTAT3 may play a critical role in cisplatin resistance in ovarian cancer stem cells, and M4, either alone or combined with cisplatin, preferentially inhibited the “stemness” potential of ovarian cancer cells. Further studies are needed to better understand the underlying mechanism involved in the inhibitory effects.
To investigate whether M4 would reduce the viability of normal cells treated with cisplatin, HOSECs and HUVECs were pretreated with 100 MOI of M4 for 48 hr. After pretreatment, the cells were exposed to 20 μM cisplatin. After 24 hr, cell viability was measured. Our results showed that cisplatin alone significantly reduced cell viability. However, treatment with M4 and cisplatin did not markedly reduce cell viability compared with those treated with cisplatin alone (Fig. 5G).
M4 potentiates the antitumor effect of cisplatin in vivo
To determine whether the effects of M4 in tumor cells were applicable to tumors in vivo, we tested tumor xenografts in athymic nude mice inoculated with Caov-3 cells. Once subcutaneous tumors were well established, treatment commenced with either cisplatin (0.75 mg/kg/day, intraperitoneal injection, every 2 days, for a total of four treatments), M4 (1×108 PFU, intratumoral injection, once daily for five consecutive days), or both cisplatin and M4. Thirty days after the last administration of M4, efficient viral replication was still evident in the tumor tissue (Fig. 6A). Consequently, M4 alone caused a significant reduction in STAT3 protein levels and a marked reduction in tumor activity (Fig. 6B and C). Cisplatin alone caused less reduction in tumor growth compared with that caused by cisplatin and M4 (Fig. 6C). Surprisingly, treatment with both M4 and cisplatin dramatically reduced the tumor volume; furthermore, after 70 days, either mice exhibited complete regression or tumor volume increased little compared with that observed immediately before treatment (Fig. 6C). Complete regression was achieved more frequently with M4 plus cisplatin treatment than with M4 or cisplatin alone (Fig. 6D).

Synergistic antitumor activity of M4 plus cisplatin in the human ovarian cancer xenograft model in mice.
Peritoneal dissemination is the primary course of progression in human ovarian cancer. The amount of ascitic fluid and the size of the disseminated tumor are correlated with the patient's prognosis. Therefore, controlling ascetic fluid and peritoneal dissemination is crucial in ovarian cancer therapy (Mabuchi et al., 2004). We used intraperitoneal xenografts in mice to examine whether M4 and cisplatin alone or in combination affected the intraabdominal dissemination of ovarian cancer and ascites formation. The results showed that cisplatin combined with M4 caused significantly greater inhibition of abdominal tumor growth than either agent alone; this was consistent with the findings in the subcutaneous xenograft study (Table 2). We found large ascites volumes in saline-treated mice, substantially attenuated ascites volumes with M4 or cisplatin treatment, and no obvious abdominal effusions in mice treated with M4 plus cisplatin (Table 2). Furthermore, tumor cell dissemination to the liver or diaphragm was commonly observed in saline-treated mice, but dissemination was significantly inhibited by treatment with M4 or M4 plus cisplatin (Table 2). M4 plus cisplatin also improved survival significantly more than either agent alone (p<0.01; Fig. 6E).
Values represent means±SD (columns 3 and 4).
Significantly different from the saline group (p<0.01).
Significantly different from the cisplatin or M4 group (p<0.01).
Adenovirus tropism to the liver has an important effect on potential clinical toxicity in humans. The hepatotoxicity induced after systemic delivery of replicating adenoviruses in murine models is dose limiting (Coughlan et al., 2009). Therefore, we used changes in serum transaminase (ALT and AST) levels and body weight variations to indicate potential M4-induced toxicities and to determine whether M4 would provoke cisplatin-induced toxicities. We found that mice treated with M4 showed a slight increase in the levels of AST and ALT compared with mice treated with saline. However, M4 did not significantly increase cisplatin-induced changes in AST and ALT (Fig. 6F). In addition, mice treated with M4 and/or cisplatin tended to show slight decreases in body weight compared with mice treated with saline; however, the differences did not reach statistical significance (Fig. 6G). Overall, intraperitoneal M4 treatment was well tolerated, with no apparent toxicity throughout the study. Collectively, the results indicated that M4 had significant antitumor effects as a single agent. M4 significantly inhibited ovarian tumor growth and metastasis, decreased the production of ascites, and prolonged survival. In addition, M4 significantly enhanced the antitumor effects of cisplatin in vivo.
Discussion
Ovarian cancer is among the most lethal of all malignancies in women. At present, the preferred treatment regimen for ovarian cancer is combination chemotherapy, usually with a platinum-based drug, such as cisplatin or carboplatin. This treatment course has shown beneficial effects in a high percentage of cases, but de novo and acquired resistance has restricted the success of this potent chemotherapeutic agent. At present, acquired resistance continues to pose a major clinical challenge. The dose escalation necessary to overcome even a small increase in cellular resistance can cause severe cytotoxicity to normal tissues (Arafa et al., 2009). The development of chemoresistance is associated with many events, including defective apoptotic signaling in response to drugs; overexpression of antiapoptotic proteins, such as IL-6, Bcl-xL, survivin, or Bcl-2; and overexpression of the multidrug resistance gene product, P-glycoprotein (Duan et al., 2009). Successful management of ovarian cancer could be greatly aided by novel agents that interfere with both intrinsic and acquired mechanisms of drug resistance.
STAT3 mediates the expression of a variety of genes in response to cell stimuli, and thus, it plays a key role in many cellular processes, including cell growth and apoptosis. A growing body of evidence has shown that STAT3 is activated constitutively in many types of cancer, including ovarian cancer, where it plays important roles in survival, drug resistance, resistance to apoptosis, and other functions (Turkson and Jove, 2000; Buettner et al., 2002; Yu and Jove, 2004; Groner et al., 2008).
This study showed that high levels of STAT3 and pSTAT3 were found in cisplatin-resistant ovarian cancer cell lines. Similar results were found in clinical tumor specimens from patients with cisplatin resistance and patients with recurrent ovarian cancer that had been treated with cisplatin. Our findings showed that cisplatin treatment resulted in upregulation of pSTAT3 expression in primary ovarian cancer cells. Moreover, higher levels of pSTAT3 were associated with more resistance to cisplatin. These findings strongly supported the clinical relevance of the STAT3 pathway in chemoresistance in ovarian cancer. Thus, inhibitors of this pathway represent a potential therapy for managing this disease.
It was previously reported that STAT3-targeted therapies could restore sensitivity to conventional chemotherapies (Alas and Bonavida, 2003; Duan et al., 2006). The original study in 2001 reported the development of a STAT3 inhibitor; since then, a variety of STAT3 inhibitors have been identified. These included direct inhibitors of STAT3 protein activity that targeted SH2-mediated dimerization, the DNA-binding domain, or the N-terminal domain, and the indirect inhibitors that targeted the upstream components of the STAT3 pathway (Yue and Turkson, 2009). STAT3 inhibitors have been shown to induce antitumor effects in vitro and in vivo. The primary limitations of inhibitors are the lack of activity, selectivity, and specificity at low concentrations. It has proven to be a formidable task to develop agents that can inhibit STAT3 with sufficient potency and good tumor selectivity. One of most promising approaches for targeting STAT3 was to inhibit phosphorylation of STAT3 with small-molecule compounds. However, some studies have demonstrated that unphosphorylated STAT3 can also affect gene expression, and it is important for oncogenesis (Yang et al., 2005; Lassmann et al., 2007). This study showed a trend toward greater STAT3 expression in recurrent tumors compared with matched primary tumors. Thus, substantial effort should be focused on the discovery of novel STAT3 inhibitors that facilitate their preclinical and eventual clinical use.
In view of the limited success of available treatment modalities for a wide array of cancers, alternative and complementary therapeutic strategies should be developed. For example, virotherapy with conditionally replicating adenoviruses (CRAds) has been shown to be a promising intervention relevant to a wide array of neoplastic diseases (Stoff-Khalili et al., 2008). Adenoviruses have many advantages, including their ability to infect a wide variety of cells, their easy replication, and their relative safety. We developed a novel CRAd, called M4, which was derived from Ad5/dE1A, an adenoviral mutant with a deletion (amino acids 121–129) in the E1A CR2 region. We replaced the E3 ADP with a fragment of reverse STAT3 cDNA to create M4. We previously demonstrated that M4 caused oncolysis and efficiently silenced STAT3 (Han et al., 2009).
In the present study, we examined whether M4 could overcome drug resistance or enhance drug sensitivity in cisplatin-resistant ovarian cancer cells. Our results showed that M4 was able to effectively silence STAT3, pSTAT3, and its target genes in ovarian cancer cells, but that it spared normal cells. We also found that M4 did not induce detectable cleavage of caspase-3 or PARP in normal cells. Furthermore, M4 did not markedly reduce cell viability when used alone or in combination with cisplatin. In ovarian cancer lines, M4 could significantly enhance cisplatin-induced apoptosis, and it reversed the drug resistance phenotype induced by IL-6. When tested on primary cultures, cells were more sensitive to the combination of cisplatin and M4 compared with either treatment alone. Importantly, M4 treatment alone could substantially eliminate the populations enriched in tumor-initiating cells; moreover, the combined M4 and cisplatin treatment led to a greater decrease in the percentage of tumor-initiating cells. The fact that these enriched populations showed resistance to cisplatin, but maintained sensitivity to the combination of cisplatin and M4, indicated that there is a need to design individualized treatments that specifically address the specific patient population that exhibits enrichment in tumor-initiating cells.
In mice inoculated with Caov-3 cells, M4 significantly enhanced the growth inhibitory effects of cisplatin. The intraperitoneal xenograft model closely reflects disease progression in humans, because these mice exhibit tumor dissemination and ascites, as observed in human ovarian cancers. This frequently leads to metastasis and dissemination throughout the peritoneum (Mabuchi et al., 2007). Treatment with M4 alone noticeably inhibited intraabdominal dissemination and the production of ascites, and this resulted in prolonged survival. Moreover, the combination treatment of M4 and cisplatin achieved a more significant therapeutic effect compared with either agent alone. In the context of conventional chemotherapy, M4 combined with cisplatin may provide a therapeutic advantage with the ability to kill cancer stem cells. Furthermore, intraperitoneal M4 treatment did not show synergistic toxicity when combined with cisplatin.
In conclusion, these results confirmed the critical role of STAT3 in cisplatin resistance in human ovarian cancer. M4 treatment blocked the STAT3 signaling pathway and reversed cisplatin resistance in human ovarian cancer cells. Combined cisplatin plus M4 therapy inhibited the growth of both noncancer stem cells and cancer stem cells and substantially improved the efficacy of chemotherapy. These results provided a further rationale for future oncolytic adenovirus-based studies to promote the development of agents that inhibit cisplatin resistance in patients with ovarian cancer.
Footnotes
Acknowledgments
Grant support: National Science Foundation of China (30973472, 30900633, 30901586, 81101963), Major Innovation Medicine Program (2009ZX09103-738, 2009ZX09103-739, 2009ZX09103-740), and the “973” Program of China (no. 2009CB521808).
Author Disclosure Statement
No competing financial interests exist.